
Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Virus Production
2.3. Flow Virometry
2.4. Plate-Based Virus Immunocapture Assay
2.5. p24 AlphaLISA
2.6. Neutralization Assay
3. Results
3.1. Using Flow Virometry to Evaluate the Staining of a Diverse Repertoire of Epitopes on HIV Env
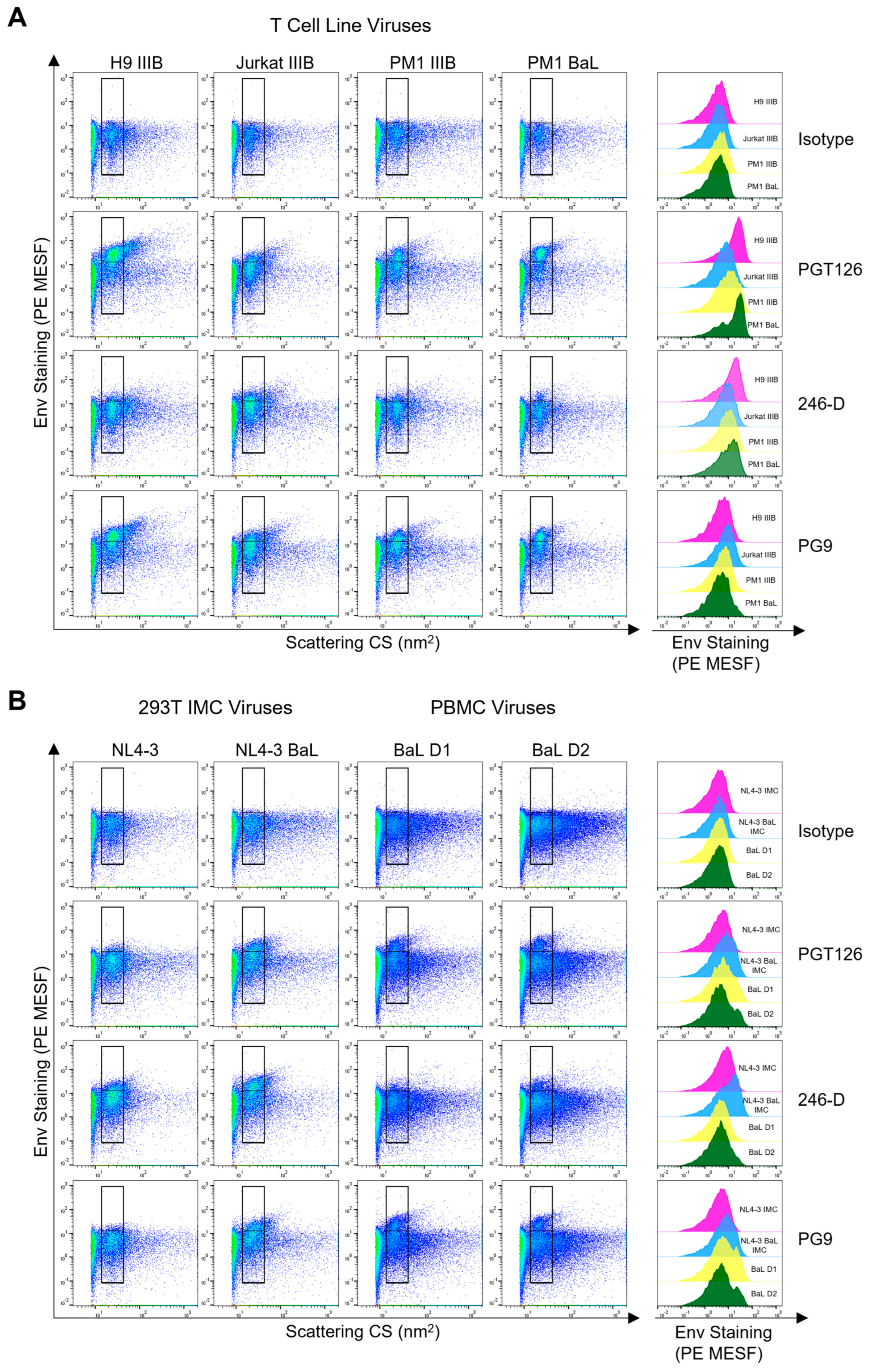
3.2. Comparing Env Staining across Different Cellular Models of Virus Production
3.3. Assessing the Performance of Antibodies in Virion Capture Assays and Virus Neutralization
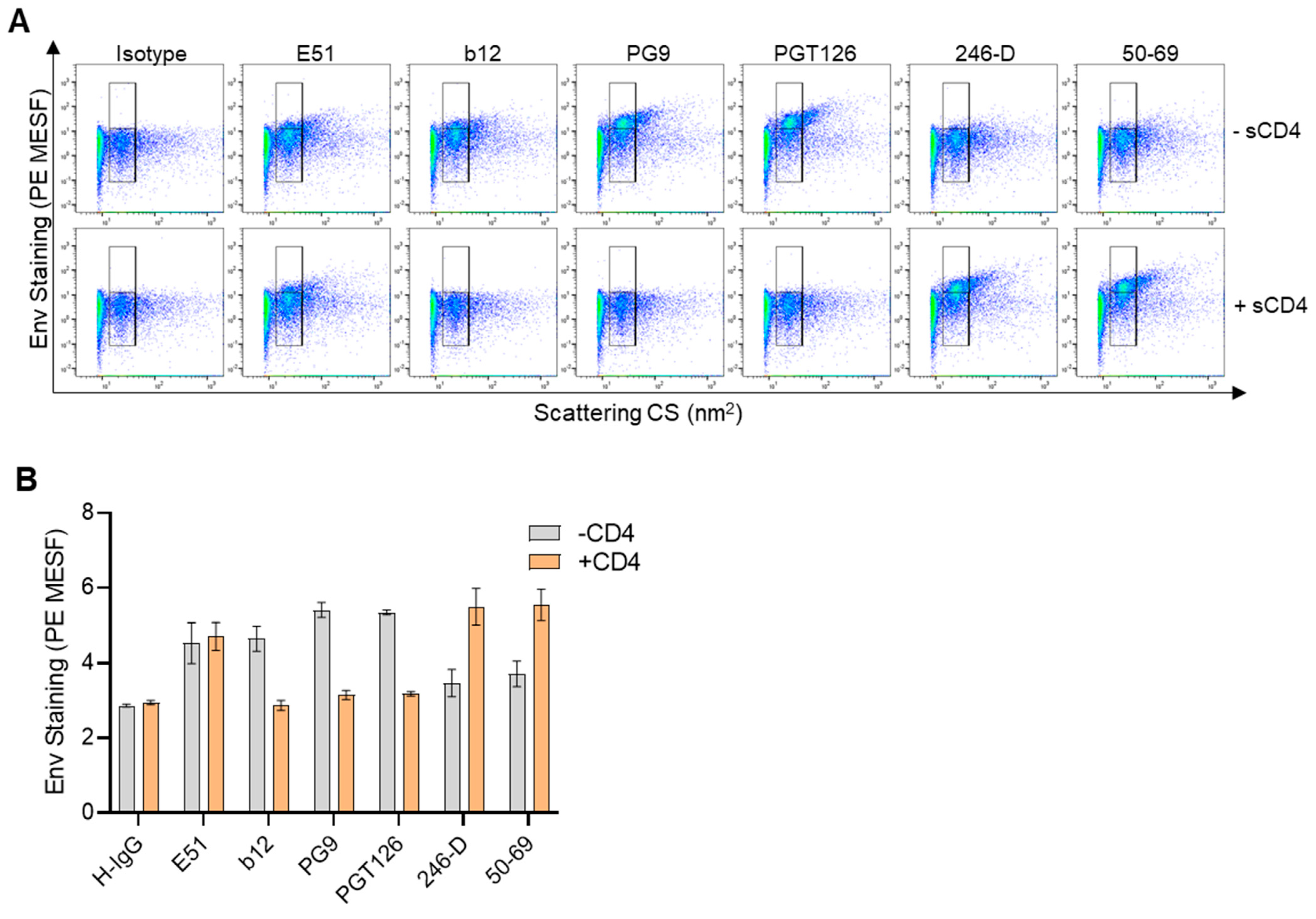
3.4. Using Flow Virometry to Evaluate the Effect of Soluble CD4 on HIV Envelope Conformation
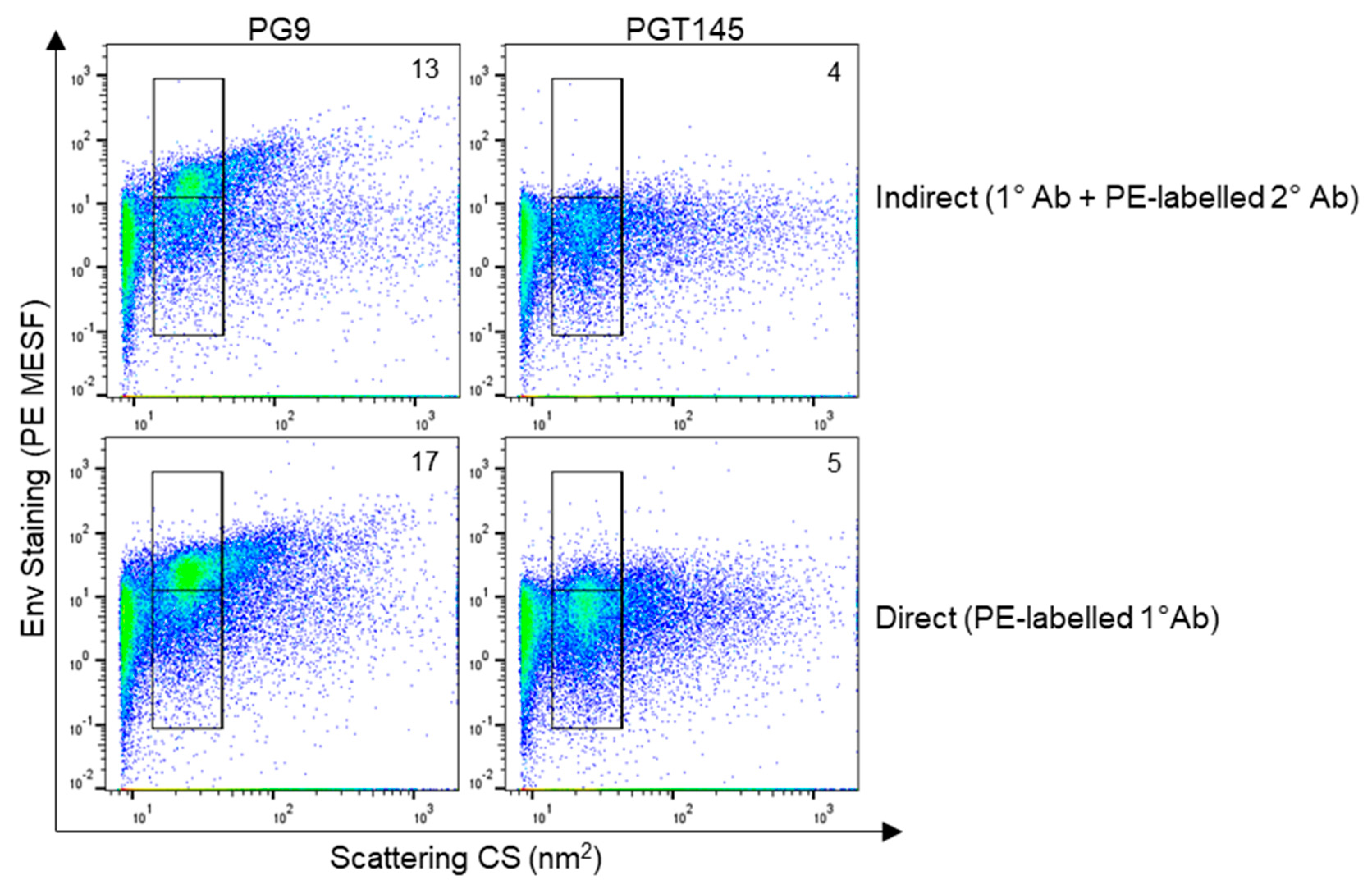
3.5. Staining HIV Env with Direct Labelling to Improve Detection Sensitivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnston, M.I.; Fauci, A.S. An HIV Vaccine—Evolving Concepts. N. Engl. J. Med. 2007, 356, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Koff, W.C.; Kwong, P.D.; Moore, J.P.; Nabel, G.J.; Sodroski, J.; Wilson, I.A.; Wyatt, R.T. HIV Vaccine Design and the Neutralizing Antibody Problem. Nat. Immunol. 2004, 5, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Overbaugh, J.; Morris, L. The Antibody Response against HIV-1. Cold Spring Harb. Perspect. Med. 2012, 2, a007039. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody Neutralization and Escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 Envelope Glycoprotein Biosynthesis, Trafficking, and Incorporation. J. Mol. Biol. 2011, 410, 582–608. [Google Scholar] [CrossRef] [PubMed]
- Chen, B. Molecular Mechanism of HIV-1 Entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell Binding and Entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866. [Google Scholar] [CrossRef] [PubMed]
- Richman, D.D.; Wrin, T.; Little, S.J.; Petropoulos, C.J. Rapid Evolution of the Neutralizing Antibody Response to HIV Type 1 Infection. Proc. Natl. Acad. Sci. USA 2003, 100, 4144–4149. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Chertova, E.; Bess, J.; Lifson, J.D.; Arthur, L.O.; Liu, J.; Taylor, K.A.; Roux, K.H. Electron Tomography Analysis of Envelope Glycoprotein Trimers on HIV and Simian Immunodeficiency Virus Virions. Proc. Natl. Acad. Sci. USA 2003, 100, 15812–15817. [Google Scholar] [CrossRef]
- Klein, J.S.; Bjorkman, P.J. Few and Far Between: How HIV May Be Evading Antibody Avidity. PLOS Pathog. 2010, 6, e1000908. [Google Scholar] [CrossRef]
- Chertova, E.; Bess, J.W.; Crise, B.J.; Sowder, R.C.; Schaden, T.M.; Hilburn, J.M.; Hoxie, J.A.; Benveniste, R.E.; Lifson, J.D.; Henderson, L.E.; et al. Envelope Glycoprotein Incorporation, Not Shedding of Surface Envelope Glycoprotein (Gp120/SU), Is the Primary Determinant of SU Content of Purified Human Immunodeficiency Virus Type 1 and Simian Immunodeficiency Virus. J. Virol. 2002, 76, 5315–5325. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Diedrich, J.K.; Kulp, D.W.; Pauthner, M.; He, L.; Park, S.-K.R.; Sok, D.; Su, C.Y.; Delahunty, C.M.; Menis, S.; et al. Global Site-Specific N-Glycosylation Analysis of HIV Envelope Glycoprotein. Nat. Commun. 2017, 8, 14954. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Finzi, A.; Sodroski, J. The Conformational States of the HIV-1 Envelope Glycoproteins. Trends Microbiol. 2020, 28, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.B.; Mothes, W. Structure and Dynamics of the Native HIV-1 Env Trimer. J. Virol. 2015, 89, 5752–5755. [Google Scholar] [CrossRef] [PubMed]
- Julien, J.-P.; Cupo, A.; Sok, D.; Stanfield, R.L.; Lyumkis, D.; Deller, M.C.; Klasse, P.-J.; Burton, D.R.; Sanders, R.W.; Moore, J.P.; et al. Crystal Structure of a Soluble Cleaved HIV-1 Envelope Trimer. Science 2013, 342, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Pancera, M.; Zhou, T.; Druz, A.; Georgiev, I.S.; Soto, C.; Gorman, J.; Huang, J.; Acharya, P.; Chuang, G.-Y.; Ofek, G.; et al. Structure and Immune Recognition of Trimeric Pre-Fusion HIV-1 Env. Nature 2014, 514, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Zolla-Pazner, S.; Cardozo, T. Structure–Function Relationships of Hiv-1 Envelope Sequence-Variable Regions Provide a Paradigm for Vaccine Design. Nat. Rev. Immunol. 2010, 10, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-Cell Responses to Transmitted Human Immunodeficiency Virus Type 1: Virion-Binding Immunoglobulin M (IgM) and IgG Antibodies Followed by Plasma Anti-Gp41 Antibodies with Ineffective Control of Initial Viremia. J. Virol. 2008, 82, 12449–12463. [Google Scholar] [CrossRef] [PubMed]
- Legrand, E.; Pellegrin, I.; Neau, D.; Pellegrin, J.-L.; Ragnaud, J.-M.; Dupon, M.; Guillemain, B.A.; Fleury, H.J. Course of Specific T Lymphocyte Cytotoxicity, Plasma and Cellular Viral Loads, and Neutralizing Antibody Titers in 17 Recently Seroconverted HIV Type 1-Infected Patients. Available online: https://www.liebertpub.com/doi/epdf/10.1089/aid.1997.13.1383 (accessed on 27 May 2023).
- Moog, C.; Fleury, H.J.; Pellegrin, I.; Kirn, A.; Aubertin, A.M. Autologous and Heterologous Neutralizing Antibody Responses Following Initial Seroconversion in Human Immunodeficiency Virus Type 1-Infected Individuals. J. Virol. 1997, 71, 3734–3741. [Google Scholar] [CrossRef]
- Beirnaert, E.; Nyambi, P.; Willems, B.; Heyndrickx, L.; Colebunders, R.; Janssens, W.; van der Groen, G. Identification and Characterization of Sera from HIV-Infected Individuals with Broad Cross-Neutralizing Activity against Group M (Env Clade A–H) and Group O Primary HIV-1 Isolates. J. Med. Virol. 2000, 62, 14–24. [Google Scholar] [CrossRef]
- Binley, J.M.; Lybarger, E.A.; Crooks, E.T.; Seaman, M.S.; Gray, E.; Davis, K.L.; Decker, J.M.; Wycuff, D.; Harris, L.; Hawkins, N.; et al. Profiling the Specificity of Neutralizing Antibodies in a Large Panel of Plasmas from Patients Chronically Infected with Human Immunodeficiency Virus Type 1 Subtypes B and C. J. Virol. 2008, 82, 11651–11668. [Google Scholar] [CrossRef] [PubMed]
- Simek, M.D.; Rida, W.; Priddy, F.H.; Pung, P.; Carrow, E.; Laufer, D.S.; Lehrman, J.K.; Boaz, M.; Tarragona-Fiol, T.; Miiro, G.; et al. Human Immunodeficiency Virus Type 1 Elite Neutralizers: Individuals with Broad and Potent Neutralizing Activity Identified by Using a High-Throughput Neutralization Assay Together with an Analytical Selection Algorithm. J. Virol. 2009, 83, 7337–7348. [Google Scholar] [CrossRef]
- Burton, D.R. Advancing an HIV Vaccine; Advancing Vaccinology. Nat. Rev. Immunol. 2019, 19, 77–78. [Google Scholar] [CrossRef]
- Spencer, D.A.; Shapiro, M.B.; Haigwood, N.L.; Hessell, A.J. Advancing HIV Broadly Neutralizing Antibodies: From Discovery to the Clinic. Front. Public Health 2021, 9, 690017. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, W.; Sun, M.; Li, T. Broadly Neutralizing Antibodies for HIV-1: Efficacies, Challenges and Opportunities. Emerg. Microbes Infect. 2020, 9, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Qureshi, H.; Deshpande, S.; Bhattacharya, J. Broadly Neutralizing Antibodies in HIV-1 Treatment and Prevention. Ther. Adv. Vaccines Immunother. 2018, 6, 61–68. [Google Scholar] [CrossRef]
- Arakelyan, A.; Fitzgerald, W.; Margolis, L.; Grivel, J.-C. Nanoparticle-Based Flow Virometry for the Analysis of Individual Virions. J. Clin. Investig. 2013, 123, 3716–3727. [Google Scholar] [CrossRef] [PubMed]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vanpouille, C.; Margolis, L. Extracellular Vesicles Carry HIV Env and Facilitate Hiv Infection of Human Lymphoid Tissue. Sci. Rep. 2017, 7, 1695. [Google Scholar] [CrossRef]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vagida, M.; Grivel, J.-C.; Margolis, L. Flow Virometry to Analyze Antigenic Spectra of Virions and Extracellular Vesicles. JoVE J. Vis. Exp. 2017, 119, e55020. [Google Scholar] [CrossRef]
- Zicari, S.; Arakelyan, A.; Fitzgerald, W.; Zaitseva, E.; Chernomordik, L.V.; Margolis, L.; Grivel, J.-C. Evaluation of the Maturation of Individual Dengue Virions with Flow Virometry. Virology 2016, 488, 20–27. [Google Scholar] [CrossRef]
- Arakelyan, A.; Petersen, J.D.; Blazkova, J.; Margolis, L. Macrophage-Derived HIV-1 Carries Bioactive TGF-Beta. Sci. Rep. 2019, 9, 19100. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.D. Optimization of Procedures for Counting Viruses by Flow Cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef] [PubMed]
- Marie, D.; Brussaard, C.P.D.; Thyrhaug, R.; Bratbak, G.; Vaulot, D. Enumeration of Marine Viruses in Culture and Natural Samples by Flow Cytometry. Appl. Environ. Microbiol. 1999, 65, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.D.; Marie, D.; Bratbak, G. Flow Cytometric Detection of Viruses. J. Virol. Methods 2000, 85, 175–182. [Google Scholar] [CrossRef]
- Bilali, N.E.; Duron, J.; Gingras, D.; Lippé, R. Quantitative Evaluation of Protein Heterogeneity within Herpes Simplex Virus 1 Particles. J. Virol. 2017, 91, 10–128. [Google Scholar] [CrossRef]
- Khadivjam, B.; El Bilali, N.; Lippé, R. Analysis and Sorting of Individual HSV-1 Particles by Flow Virometry. In Herpes Simplex Virus: Methods and Protocols; Diefenbach, R.J., Fraefel, C., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; pp. 289–303. ISBN 978-1-4939-9814-2. [Google Scholar]
- Loret, S.; Bilali, N.E.; Lippé, R. Analysis of Herpes Simplex Virus Type I Nuclear Particles by Flow Cytometry. Cytom. A 2012, 81A, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Bonar, M.M.; Tilton, J.C. High Sensitivity Detection and Sorting of Infectious Human Immunodeficiency Virus (HIV-1) Particles by Flow Virometry. Virology 2017, 505, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Bonar, M.M.; Tabler, C.O.; Haqqani, A.A.; Lapointe, L.E.; Galiatsos, J.A.; Joussef-Piña, S.; Quiñones-Mateu, M.E.; Tilton, J.C. Nanoscale Flow Cytometry Reveals Interpatient Variability in HIV Protease Activity That Correlates with Viral Infectivity and Identifies Drug-Resistant Viruses. Sci. Rep. 2020, 10, 18101. [Google Scholar] [CrossRef]
- Brittain, G.C.; Chen, Y.Q.; Martinez, E.; Tang, V.A.; Renner, T.M.; Langlois, M.-A.; Gulnik, S. A Novel Semiconductor-Based Flow Cytometer with Enhanced Light-Scatter Sensitivity for the Analysis of Biological Nanoparticles. Sci. Rep. 2019, 9, 16039. [Google Scholar] [CrossRef]
- Welsh, J.A.; Jones, J.C.; Tang, V.A. Fluorescence and Light Scatter Calibration Allow Comparisons of Small Particle Data in Standard Units across Different Flow Cytometry Platforms and Detector Settings. Cytom. A 2020, 97, 592–601. [Google Scholar] [CrossRef]
- Maltseva, M.; Langlois, M.-A. Influence of GlycoGag on the Incorporation of Host Membrane Proteins Into the Envelope of the Moloney Murine Leukemia Virus. Front. Virol. 2021, 1, 747253. [Google Scholar] [CrossRef]
- Renner, T.M.; Tang, V.A.; Burger, D.; Langlois, M.-A. Intact Viral Particle Counts Measured by Flow Virometry Provide Insight into the Infectivity and Genome Packaging Efficiency of Moloney Murine Leukemia Virus. J. Virol. 2020, 94, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Morales-Kastresana, A.; Musich, T.A.; Welsh, J.A.; Telford, W.; Demberg, T.; Wood, J.C.S.; Bigos, M.; Ross, C.D.; Kachynski, A.; Dean, A.; et al. High-Fidelity Detection and Sorting of Nanoscale Vesicles in Viral Disease and Cancer. J. Extracell. Vesicles 2019, 8, 1597603. [Google Scholar] [CrossRef] [PubMed]
- Morales-Kastresana, A.; Telford, B.; Musich, T.A.; McKinnon, K.; Clayborne, C.; Braig, Z.; Rosner, A.; Demberg, T.; Watson, D.C.; Karpova, T.S. Labeling Extracellular Vesicles for Nanoscale Flow Cytometry. Sci. Rep. 2017, 7, 1878. [Google Scholar] [CrossRef] [PubMed]
- Burnie, J.; Tang, V.A.; Welsh, J.A.; Persaud, A.T.; Thaya, L.; Jones, J.C.; Guzzo, C. Flow Virometry Quantification of Host Proteins on the Surface of HIV-1 Pseudovirus Particles. Viruses 2020, 12, 1296. [Google Scholar] [CrossRef] [PubMed]
- Burnie, J.; Persaud, A.T.; Thaya, L.; Liu, Q.; Miao, H.; Grabinsky, S.; Norouzi, V.; Lusso, P.; Tang, V.A.; Guzzo, C. P-Selectin Glycoprotein Ligand-1 (PSGL-1/CD162) Is Incorporated into Clinical HIV-1 Isolates and Can Mediate Virus Capture and Subsequent Transfer to Permissive Cells. Retrovirology 2022, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Maltseva, M.; Langlois, M.-A. Flow Virometry for Characterizing the Size, Concentration, and Surface Antigens of Viruses. Curr. Protoc. 2022, 2, e368. [Google Scholar] [CrossRef] [PubMed]
- Tang, V.A.; Renner, T.M.; Varette, O.; Le Boeuf, F.; Wang, J.; Diallo, J.-S.; Bell, J.C.; Langlois, M.-A. Single-Particle Characterization of Oncolytic Vaccinia Virus by Flow Virometry. Vaccine 2016, 34, 5082–5089. [Google Scholar] [CrossRef] [PubMed]
- Tang, V.A.; Renner, T.M.; Fritzsche, A.K.; Burger, D.; Langlois, M.-A. Single-Particle Discrimination of Retroviruses from Extracellular Vesicles by Nanoscale Flow Cytometry. Sci. Rep. 2017, 7, 17769. [Google Scholar] [CrossRef] [PubMed]
- Burnie, J.; Fernandes, C.; Chaphekar, D.; Wei, D.; Ahmed, S.; Persaud, A.T.; Khader, N.; Cicala, C.; Arthos, J.; Tang, V.A.; et al. Identification of CD38, CD97, and CD278 on the HIV Surface Using a Novel Flow Virometry Screening Assay. Sci. Rep. 2023, 13, 23025. [Google Scholar] [CrossRef]
- Persaud, A.T.; Khela, J.; Fernandes, C.; Chaphekar, D.; Burnie, J.; Tang, V.A.; Colpitts, C.C.; Guzzo, C. Virion-Incorporated CD14 Enables HIV-1 to Bind LPS and Initiate TLR4 Signaling in Immune Cells. J. Virol. 2024, 98, e00363-24. [Google Scholar] [CrossRef]
- Tang, V.A.; Fritzsche, A.K.; Renner, T.M.; Burger, D.; van der Pol, E.; Lannigan, J.A.; Brittain, G.C.; Welsh, J.A.; Jones, J.C.; Langlois, M.-A. Engineered Retroviruses as Fluorescent Biological Reference Particles for Small Particle Flow Cytometry. bioRxiv 2019, 614461. [Google Scholar] [CrossRef]
- Musich, T.; Jones, J.C.; Keele, B.F.; Jenkins, L.M.M.; Demberg, T.; Uldrick, T.S.; Yarchoan, R.; Robert-Guroff, M. Flow Virometric Sorting and Analysis of HIV Quasispecies from Plasma. JCI Insight 2017, 2, e90626. [Google Scholar] [CrossRef]
- Staropoli, I.; Dufloo, J.; Ducher, A.; Commere, P.-H.; Sartori-Rupp, A.; Novault, S.; Bruel, T.; Lorin, V.; Mouquet, H.; Schwartz, O.; et al. Flow Cytometry Analysis of HIV-1 Env Conformations at the Surface of Infected Cells and Virions: Role of Nef, CD4, and SERINC5. J. Virol. 2020, 94, pp.10–1128. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Jones, J.C. Small Particle Fluorescence and Light Scatter Calibration Using FCMPASS Software. Curr. Protoc. Cytom. 2020, 94, e79. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.-Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and Potent Neutralizing Antibodies from an African Donor Reveal a New HIV-1 Vaccine Target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.-P.; Wang, S.-K.; Ramos, A.; Chan-Hui, P.-Y.; Moyle, M.; et al. Broad Neutralization Coverage of HIV by Multiple Highly Potent Antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Gorny, M.K.; Palker, T.; Karwowska, S.; Zolla-Pazner, S. Epitope Mapping of Two Immunodominant Domains of Gp41, the Transmembrane Protein of Human Immunodeficiency Virus Type 1, Using Ten Human Monoclonal Antibodies. J. Virol. 1991, 65, 4832–4838. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.; Prévost, J.; Baxter, A.E.; von Bredow, B.; Ding, S.; Medjahed, H.; Delgado, G.G.; Brassard, N.; Stürzel, C.M.; Kirchhoff, F.; et al. Uninfected Bystander Cells Impact the Measurement of HIV-Specific Antibody-Dependent Cellular Cytotoxicity Responses. mBio 2018, 9, e00358-18. [Google Scholar] [CrossRef] [PubMed]
- Nyambi, P.N.; Mbah, H.A.; Burda, S.; Williams, C.; Gorny, M.K.; Nádas, A.; Zolla-Pazner, S. Conserved and Exposed Epitopes on Intact, Native, Primary Human Immunodeficiency Virus Type 1 Virions of Group M. J. Virol. 2000, 74, 7096–7107. [Google Scholar] [CrossRef]
- Chen, Q.; Nie, J.; Huang, W.; Jiao, Y.; Li, L.; Zhang, T.; Zhao, J.; Wu, H.; Wang, Y. Development and Optimization of a Sensitive Pseudovirus-Based Assay for HIV-1 Neutralizing Antibodies Detection Using A3R5 Cells. Hum. Vaccines Immunother. 2017, 14, 199–208. [Google Scholar] [CrossRef]
- Rujas, E.; Kucharska, I.; Tan, Y.Z.; Benlekbir, S.; Cui, H.; Zhao, T.; Wasney, G.A.; Budylowski, P.; Guvenc, F.; Newton, J.C.; et al. Multivalency Transforms SARS-CoV-2 Antibodies into Broad and Ultrapotent Neutralizers. bioRxiv 2020. [Google Scholar] [CrossRef]
- Montefiori, D.C. Measuring HIV Neutralization in a Luciferase Reporter Gene Assay. In HIV Protocols; Prasad, V.R., Kalpana, G.V., Eds.; Methods In Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2009; pp. 395–405. ISBN 978-1-59745-170-3. [Google Scholar]
- Doores, K.J.; Burton, D.R. Variable Loop Glycan Dependency of the Broad and Potent HIV-1-Neutralizing Antibodies PG9 and PG16. J. Virol. 2010, 84, 10510–10521. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Pauthner, M.; Andrabi, R.; Rantalainen, K.; Berndsen, Z.; Diedrich, J.K.; Menis, S.; Sok, D.; Bastidas, R.; Park, S.-K.R.; et al. Differential Processing of HIV Envelope Glycans on the Virus and Soluble Recombinant Trimer. Nat. Commun. 2018, 9, 3693. [Google Scholar] [CrossRef] [PubMed]
- Brandenberg, O.F.; Magnus, C.; Rusert, P.; Regoes, R.R.; Trkola, A. Different Infectivity of HIV-1 Strains Is Linked to Number of Envelope Trimers Required for Entry. PLOS Pathog. 2015, 11, e1004595. [Google Scholar] [CrossRef]
- Yang, X.; Kurteva, S.; Ren, X.; Lee, S.; Sodroski, J. Stoichiometry of Envelope Glycoprotein Trimers in the Entry of Human Immunodeficiency Virus Type 1. J. Virol. 2005, 79, 12132–12147. [Google Scholar] [CrossRef]
- Orentas, R.J.; Hildreth, J.E.K. Association of Host Cell Surface Adhesion Receptors and Other Membrane Proteins with HIV and SIV. AIDS Res. Hum. Retroviruses 1993, 9, 1157–1165. [Google Scholar] [CrossRef]
- Frank, I.; Stoiber, H.; Godar, S.; Stockinger, H.; Steindl, F.; Katinger, H.W.D.; Dierich, M.P. Acquisition of Host Cell-Surface-Derived Molecules by HIV-1. AIDS 1996, 10, 1611–1620. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Cornell, R.J.; Zhou, J.Y.; Zhou, J.T.; Hirsch, V.M.; Johnson, P.R. Complement Control Proteins, CD46, CD55, and CD59, as Common Surface Constituents of Human and Simian Immunodeficiency Viruses and Possible Targets for Vaccine Protection. Virology 1994, 205, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, J.A.; Bar-On, Y.; Lu, C.-L.; Fera, D.; Lockhart, A.A.K.; Lorenzi, J.C.C.; Nogueira, L.; Golijanin, J.; Scheid, J.F.; Seaman, M.S.; et al. Non-Neutralizing Antibodies Alter the Course of HIV-1 Infection in Vivo. Cell 2017, 170, 637–648.e10. [Google Scholar] [CrossRef]
- Ramdas, P.; Sahu, A.K.; Mishra, T.; Bhardwaj, V.; Chande, A. From Entry to Egress: Strategic Exploitation of the Cellular Processes by HIV-1. Front. Microbiol. 2020, 11, 559792. [Google Scholar] [CrossRef]
- Huang, C.; Venturi, M.; Majeed, S.; Moore, M.J.; Phogat, S.; Zhang, M.-Y.; Dimitrov, D.S.; Hendrickson, W.A.; Robinson, J.; Sodroski, J.; et al. Structural Basis of Tyrosine Sulfation and VH-Gene Usage in Antibodies That Recognize the HIV Type 1 Coreceptor-Binding Site on Gp120. Proc. Natl. Acad. Sci. USA 2004, 101, 2706–2711. [Google Scholar] [CrossRef] [PubMed]
- Choe, H.; Li, W.; Wright, P.L.; Vasilieva, N.; Venturi, M.; Huang, C.-C.; Grundner, C.; Dorfman, T.; Zwick, M.B.; Wang, L.; et al. Tyrosine Sulfation of Human Antibodies Contributes to Recognition of the CCR5 Binding Region of HIV-1 Gp120. Cell 2003, 114, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Gorny, M.K.; Gianakakos, V.; Sharpe, S.; Zolla-Pazner, S. Generation of Human Monoclonal Antibodies to Human Immunodeficiency Virus. Proc. Natl. Acad. Sci. USA 1989, 86, 1624–1628. [Google Scholar] [CrossRef] [PubMed]
- Peachman, K.K.; Wieczorek, L.; Polonis, V.R.; Alving, C.R.; Rao, M. The Effect of sCD4 on the Binding and Accessibility of HIV-1 Gp41 MPER Epitopes to Human Monoclonal Antibodies. Virology 2010, 408, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.A.; Abrams, B.; Iyer, S.B.; Hoffman, R.A.; Bishop, J.E. Determination of CD4 Antigen Density on Cells: Role of Antibody Valency, Avidity, Clones, and Conjugation. Cytometry 1998, 33, 197–205. [Google Scholar] [CrossRef]
- BD QuantibriteTM Beads. Available online: https://www.bdbiosciences.com/content/bdb/paths/generate-tds-document.us.340495.pdf (accessed on 5 January 2024).
- Gratama, J.W.; D’hautcourt, J.-L.; Mandy, F.; Rothe, G.; Barnett, D.; Janossy, G.; Papa, S.; Schmitz, G.; Lenkei, R.; European Working Group on Clinical Cell Analysis. Flow Cytometric Quantitation of Immunofluorescence Intensity: Problems and Perspectives. Cytometry 1998, 33, 166–178. [Google Scholar] [CrossRef]
- Lusso, P. The Quest for an HIV-1 Vaccine: Will mRNA Deliver Us from Evil? Expert Rev. Vaccines 2023, 22, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Wagh, K.; Bhattacharya, T.; Williamson, C.; Robles, A.; Bayne, M.; Garrity, J.; Rist, M.; Rademeyer, C.; Yoon, H.; Lapedes, A.; et al. Optimal Combinations of Broadly Neutralizing Antibodies for Prevention and Treatment of HIV-1 Clade C Infection. PLoS Pathog. 2016, 12, e1005520. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Louder, M.K.; Wagh, K.; Bailer, R.T.; deCamp, A.; Greene, K.; Gao, H.; Taft, J.D.; Gazumyan, A.; Liu, C.; et al. Improving Neutralization Potency and Breadth by Combining Broadly Reactive HIV-1 Antibodies Targeting Major Neutralization Epitopes. J. Virol. 2015, 89, 2659–2671. [Google Scholar] [CrossRef]
- Miner, M.D.; Corey, L.; Montefiori, D. Broadly Neutralizing Monoclonal Antibodies for HIV Prevention. J. Int. AIDS Soc. 2021, 24, e25829. [Google Scholar] [CrossRef]
- Labrijn, A.F.; Poignard, P.; Raja, A.; Zwick, M.B.; Delgado, K.; Franti, M.; Binley, J.; Vivona, V.; Grundner, C.; Huang, C.-C.; et al. Access of Antibody Molecules to the Conserved Coreceptor Binding Site on Glycoprotein Gp120 Is Sterically Restricted on Primary Human Immunodeficiency Virus Type 1. J. Virol. 2003, 77, 10557–10565. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Liu, J.; Bess, J.; Chertova, E.; Lifson, J.D.; Grisé, H.; Ofek, G.A.; Taylor, K.A.; Roux, K.H. Distribution and Three-Dimensional Structure of AIDS Virus Envelope Spikes. Nature 2006, 441, 847. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Arkesteijn, G.J.A.; Bremer, M.; Cimorelli, M.; Dignat-George, F.; Giebel, B.; Görgens, A.; Hendrix, A.; Kuiper, M.; Lacroix, R.; et al. A Compendium of Single Extracellular Vesicle Flow Cytometry. J. Extracell. Vesicles 2023, 12, e12299. [Google Scholar] [CrossRef] [PubMed]
- Binley, J.M.; Sanders, R.W.; Clas, B.; Schuelke, N.; Master, A.; Guo, Y.; Kajumo, F.; Anselma, D.J.; Maddon, P.J.; Olson, W.C.; et al. A Recombinant Human Immunodeficiency Virus Type 1 Envelope Glycoprotein Complex Stabilized by an Intermolecular Disulfide Bond between the Gp120 and Gp41 Subunits Is an Antigenic Mimic of the Trimeric Virion-Associated Structure. J. Virol. 2000, 74, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.W.; Vesanen, M.; Schuelke, N.; Master, A.; Schiffner, L.; Kalyanaraman, R.; Paluch, M.; Berkhout, B.; Maddon, P.J.; Olson, W.C.; et al. Stabilization of the Soluble, Cleaved, Trimeric Form of the Envelope Glycoprotein Complex of Human Immunodeficiency Virus Type 1. J. Virol. 2002, 76, 8875–8889. [Google Scholar] [CrossRef]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.-P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Peña, A.T.; Korzun, J.; et al. A Next-Generation Cleaved, Soluble HIV-1 Env Trimer, BG505 SOSIP.664 Gp140, Expresses Multiple Epitopes for Broadly Neutralizing but Not Non-Neutralizing Antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HIV Env Trimer Domain | Antibody | ARP Catalog # | Mean (PE MESF) | SD |
|---|---|---|---|---|
| V3 glycan supersite | 257-D IV | 1510 | 0.08 | 0.15 |
| F425 B4e8 | 7626 | - | - | |
| 39F | 11437 | - | - | |
| PGT126 | 12344 | 11.27 | 0.19 | |
| 10065D | 13426 | - | - | |
| 2557 | 13429 | 0.60 | 0.27 | |
| 2558 | 13432 | 1.49 | 0.28 | |
| 2191 | 11682 | 1.98 | 0.34 | |
| 10074 | 12477 | 1.42 | 0.28 | |
| 447-52D | 4030 | 0.65 | 0.10 | |
| F425 B4a1 | 7625 | 0.01 | 0.06 | |
| 3074 | 12040 | - | - | |
| 2G12 | 1476 | 0.62 | 0.12 | |
| 268-D IV | 1511 | - | - | |
| 3869 | 12039 | - | - | |
| PGT128 | 13352 | 10.92 | 0.64 | |
| 2219 | 11683 | - | - | |
| PGT121 | 12343 | 0.74 | 0.16 | |
| 5F7 | 2533 | 0.57 | 0.20 | |
| gp160 | 902 | 522 | 1.16 | 0.34 |
| Chessie 6 | 810 | 0.20 | 0.08 | |
| ID6 | 2343 | 0.46 | 0.14 | |
| gp120 | IgG YZ23 | 12047 | 0.05 | 0.14 |
| Constant region 1 | A32 | 11438 | - | - |
| 654-30D | 7369 | - | - | |
| CH38 | 12548 | - | - | |
| CH57 | 12549 | - | - | |
| CH90 | 12552 | - | - | |
| A32-AAA | 12558 | 0.02 | 0.09 | |
| 16H3 | 12559 | 0.17 | 0.07 | |
| 3B3 | 12560 | 0.16 | 0.08 | |
| Chessie 13-39.1 | 1209 | 0.09 | 0.06 | |
| Constant region 5 | 670-30D | 7370 | 0.06 | 0.06 |
| CD4bs | VRC01 | 12033 | 0.21 | 0.10 |
| 3BNC117 | 12474 | 0.27 | 0.07 | |
| VRC-CH31 | 12565 | - | - | |
| CH106 | 12566 | 0.14 | 0.08 | |
| F105 | 857 | 0.02 | 0.09 | |
| * N6/PGDM1400x10E8 | 13390 | 2.53 | 0.35 | |
| N6 | 12968 | 0.22 | 0.19 | |
| VRC03 | 12032 | 1.19 | 0.22 | |
| NIH45-46 G54W | 12174 | 1.85 | 0.57 | |
| IgG1 b12 | 2640 | 1.47 | 0.07 | |
| HJ16 | 12138 | 1.52 | 0.09 | |
| CD4i | E51 | 11439 | 2.76 | 0.14 |
| scFv m9 | 11710 | 0.05 | 0.08 | |
| F425 A1g8 | 7624 | 1.07 | 0.37 | |
| 17b | 4091 | 0.81 | 0.14 | |
| 48d | 1756 | 0.95 | 0.23 | |
| gp120 interface | 35022 | 12586 | 0.07 | 0.08 |
| MPER | Z13e1 | 11557 | - | - |
| 10E8 | 12294 | - | - | |
| 7H6 | 12295 | - | - | |
| 10E8v4 | 12865 | - | - | |
| 2F5 | 1475 | - | - | |
| 4E10 | 10091 | - | - | |
| V1V2 | CH58 | 12550 | - | - |
| CH59 | 12551 | - | - | |
| HG107 | 12553 | - | - | |
| HG120 | 12554 | - | - | |
| CH01 | 12561 | - | - | |
| CH01 | 12562 | - | - | |
| CH03 | 12563 | - | - | |
| CH04 | 12564 | - | - | |
| PG9 | 12149 | 7.62 | 0.16 | |
| PG16 | 12150 | 2.94 | 0.24 | |
| 2909 | 12141 | - | - | |
| PGT145 | 12703 | 0.08 | 0.07 | |
| 697-30D | 7371 | - | - | |
| Ibalizumab (PG9-iMab) | 12633 | 0.40 | 0.07 | |
| gp41 | 246-D | 1245 | 1.95 | 0.19 |
| 7B2 | 12556 | 1.02 | 0.32 | |
| 7B2-AAA | 12557 | 1.87 | 0.07 | |
| 240-D | 1242 | 1.74 | 0.31 | |
| T32 | 11391 | 0.47 | 0.12 | |
| NC | 11482 | 0.08 | 0.06 | |
| 5F3 | 6882 | 0.01 | 0.18 | |
| F240 | 7623 | 1.97 | 0.12 | |
| D50 | 11393 | 0.11 | 0.08 | |
| 1577 | 1172 | 0.08 | 0.06 | |
| Chessie 8 | 13049 | 0.08 | 0.09 | |
| 167-D IV | 11681 | - | - | |
| 50-69 | 531 | 1.91 | 0.20 | |
| D5 | 12296 | - | - | |
| 126-7 | 9967 | 0.03 | 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burnie, J.; Fernandes, C.; Patel, A.; Persaud, A.T.; Chaphekar, D.; Wei, D.; Lee, T.K.H.; Tang, V.A.; Cicala, C.; Arthos, J.; et al. Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems. Viruses 2024, 16, 935. https://doi.org/10.3390/v16060935
Burnie J, Fernandes C, Patel A, Persaud AT, Chaphekar D, Wei D, Lee TKH, Tang VA, Cicala C, Arthos J, et al. Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems. Viruses. 2024; 16(6):935. https://doi.org/10.3390/v16060935
Chicago/Turabian StyleBurnie, Jonathan, Claire Fernandes, Ayushi Patel, Arvin Tejnarine Persaud, Deepa Chaphekar, Danlan Wei, Timothy Kit Hin Lee, Vera A. Tang, Claudia Cicala, James Arthos, and et al. 2024. "Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems" Viruses 16, no. 6: 935. https://doi.org/10.3390/v16060935
APA StyleBurnie, J., Fernandes, C., Patel, A., Persaud, A. T., Chaphekar, D., Wei, D., Lee, T. K. H., Tang, V. A., Cicala, C., Arthos, J., & Guzzo, C. (2024). Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems. Viruses, 16(6), 935. https://doi.org/10.3390/v16060935