
Phylogenetic Network Analyses Reveal the Influence of Transmission Clustering on the Spread of HIV Drug Resistance in Quebec from 2002 to 2022

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
- Transmitted resistance (TDR) in ART-naïve persons acquiring subtype-B infections (n = 6011): this group encompassed the predominant men having sex with men (MSM) epidemic (male singletons and male–male clusters, n = 4854) and the heterosexual (HET) subtype B epidemic from persons arriving from Haiti, the Caribbean, and Latin and South America (female singletons and mixed gender clusters, 56% female, n = 1157).
- Acquired resistance (ADR) in chronic treated persons with subtype-B infections (n = 3500): this group incorporated all genotyped persons failing first or subsequent treatment regimens (n = 6013 sequences from 3500 persons).
- Pretreatment resistance (PDR) and ADR in persons having non-B-subtype infections (n = 2065) this group included newcomers to the province arriving from francophone countries in Africa, Europe, and Asia. Overall, 1577 of those genotyped were reported as ART-naive.
2.2. Sequence Analyses of Transmission Networks and Drug Resistance
2.3. Drug Resistance and Statistical Analyses
3. Results
3.1. Overall Prevalence of Acquired and Transmitted Drug Resistance in Quebec
3.2. Longitudinal Trends in the Spread of Acquired and Transmitted Resistance
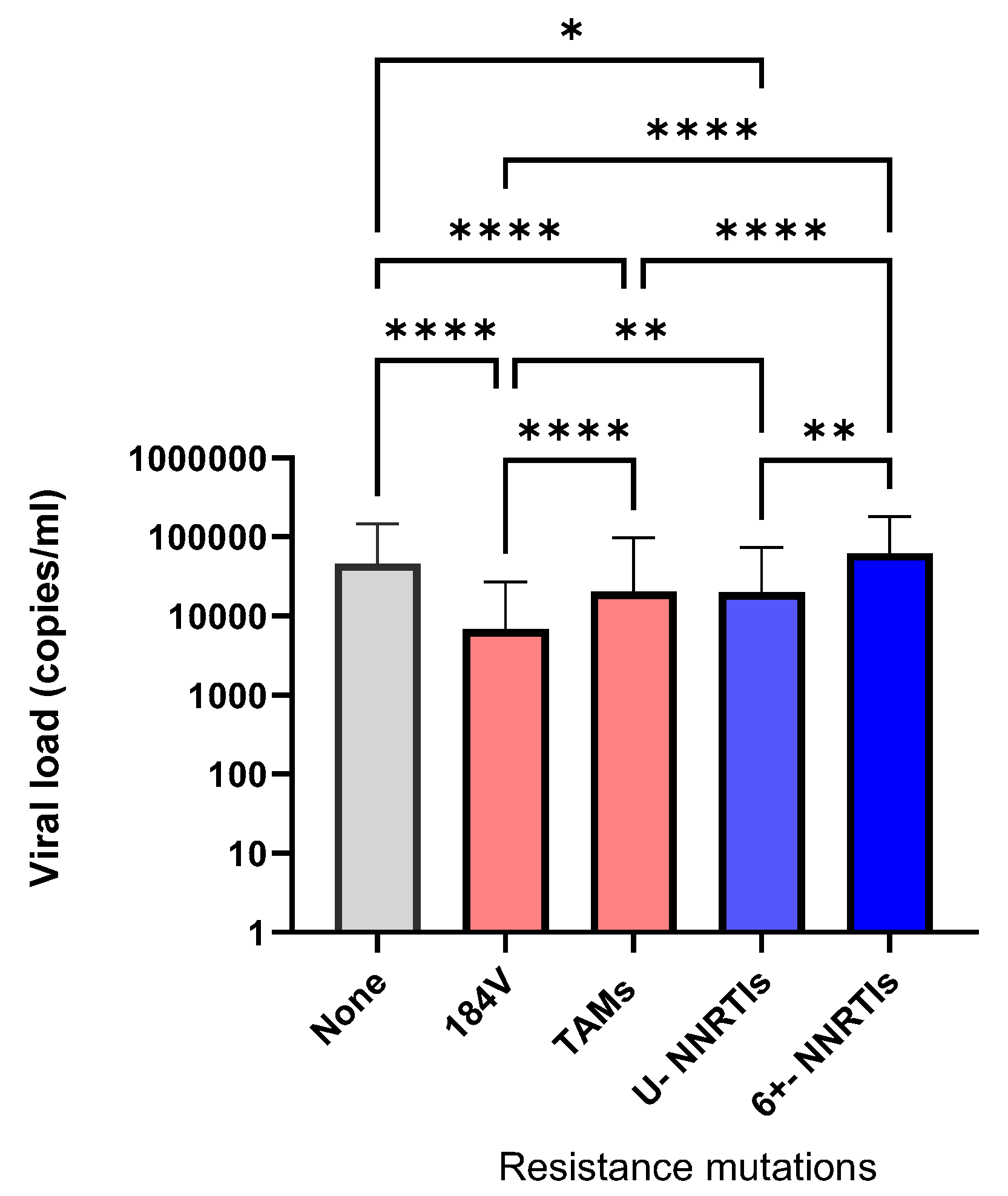
3.3. Influence of Clustered Outbreaks on the Persistence of HIV TDR in Newly Infected Persons
3.4. Acquired and Transmitted Resistance to Integrase Inhibitor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNAIDS Global AIDS Update 2023. The Path That Ends AIDS; Joint United Nations Programme on HIV/AIDS: Geneva, Switzerland, 2023; Available online: https://thepath.unaids.org/wp-content/themes/unaids2023/assets/files/2023_report.pdf (accessed on 24 July 2024).
- World Health Organization. Tackling Drug Resistance: Trends, Guidelines and Global Action. 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/255881/WHO-HIV-2017.21-eng.pdf (accessed on 24 July 2024).
- World Health Organization. HIV Drug Resistance Report 2021. 2021. Available online: https://www.aidsdatahub.org/sites/default/files/resource/who-hiv-drug-resistance-report-2021.pdf (accessed on 24 July 2024).
- Sanaubarova, A.; Pujol-Hodge, E.; Dzissyuk, N.; Lemey, P.; Vermund, S.H.; Leigh Brown, A.J.; Ali, S. High-Level Drug-Resistant Mutations among HIV-1 Subtype A6 and CRF02_AG in Kazakhstan. Viruses 2023, 15, 1407. [Google Scholar] [CrossRef] [PubMed]
- Palich, R.; Teyssou, E.; Sayon, S.; Abdi, B.; Soulie, C.; Cuzin, L.; Tubiana, R.; Valantin, M.A.; Schneider, L.; Seang, S.; et al. Kinetics of Archived M184V Mutation in Treatment-Experienced Virally Suppressed HIV-Infected Patients. J. Infect. Dis. 2022, 225, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Harris, E. WHO Documents Rising Resistance to First-Line HIV Drug. JAMA 2024, 331, 1355. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M.; Blanco, J.L.; Marcelin, A.G.; Perno, C.F.; Stellbrink, H.J.; Turner, D.; Zengin, T. HIV DNA Sequencing to Detect Archived Antiretroviral Drug Resistance. Infect. Dis. Ther. 2022, 11, 1793–1803. [Google Scholar] [CrossRef] [PubMed]
- Kabra, M.; Barber, T.J.; Allavena, C.; Marcelin, A.G.; Di Giambenedetto, S.; Pasquau, J.; Gianotti, N.; Llibre, J.M.; Rial-Crestelo, D.; De Miguel-Buckley, R.; et al. Virologic Response to Dolutegravir Plus Lamivudine in People With Suppressed Human Immunodeficiency Virus Type 1 and Historical M184V/I: A Systematic Literature Review and Meta-analysis. Open Forum Infect. Dis. 2023, 10, ofad526. [Google Scholar] [CrossRef] [PubMed]
- Landman, R.; Assoumou, L.; Lambert-Niclot, S.; Amat, K.; Katlama, C.; Allavena, C.; Lacombe, K.; Bouchaud, O.; de Castro, N.; Moins, C.; et al. ANRS 177 DUETTO randomized, open-label and multicentric trial evaluating the non-inferiority of maintenance antiretroviral dual therapy taken 4 consecutive days per week versus dual therapy 7/7 days per week. In Proceedings of the 19th European AIDS Conference, Warsaw, Poland, 18–21 October 2023. poster eP.A.106. [Google Scholar]
- Brenner, B.G.; Ibanescu, R.I.; Osman, N.; Cuadra-Foy, E.; Oliveira, M.; Chaillon, A.; Stephens, D.; Hardy, I.; Routy, J.P.; Thomas, R.; et al. The Role of Phylogenetics in Unravelling Patterns of HIV Transmission towards Epidemic Control: The Quebec Experience (2002–2020). Viruses 2021, 13, 1643. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.G.; Lowe, M.; Moisi, D.; Hardy, I.; Gagnon, S.; Charest, H.; Baril, J.G.; Wainberg, M.A.; Roger, M. Subtype diversity associated with the development of HIV-1 resistance to integrase inhibitors. J. Med. Virol. 2011, 83, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.M.; Boyles, A.; Shankar, A.; Kim, J.; Knyazev, S.; Cintron, R.; Switzer, W.M. MicrobeTrace: Retooling molecular epidemiology for rapid public health response. PLoS Comput. Biol. 2021, 17, e1009300. [Google Scholar] [CrossRef] [PubMed]
- Mbuagbaw, L.; Garcia, C.; Brenner, B.; Cecchini, D.; Chakroun, M.; Djiadeu, P.; Holguin, A.; Mor, O.; Parkin, N.; Santoro, M.; et al. Checklist for studies of HIV drug resistance prevalence and incidence (CEDRIC-HIV): Rationale and recommended use. Lancet HIV 2023, 10, e684–e689. [Google Scholar] [CrossRef]
- Bennett, D.E.; Camacho, R.J.; Otelea, D.; Kuritzkes, D.R.; Fleury, H.; Kiuchi, M.; Heneine, W.; Kantor, R.; Jordan, M.R.; Schapiro, J.M.; et al. Drug resistance mutations for surveillance of transmitted HIV-1 drug-resistance: 2009 update. PLoS ONE 2009, 4, e4724. [Google Scholar] [CrossRef]
- Tzou, P.L.; Rhee, S.Y.; Descamps, D.; Clutter, D.S.; Hare, B.; Mor, O.; Grude, M.; Parkin, N.; Jordan, M.R.; Bertagnolio, S.; et al. Integrase strand transfer inhibitor (INSTI)-resistance mutations for the surveillance of transmitted HIV-1 drug resistance. J. Antimicrob. Chemother. 2020, 75, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Sayan, M.; Sultanoglu, N.; Sanlidag, T. Dynamics of Rilpivirine Resistance-Associated Mutation: E138 in Reverse Transcriptase among Antiretroviral-Naive HIV-1-Infected Individuals in Turkey. AIDS Res. Hum. Retroviruses 2023, 39, 84–90. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.; Oliveira, M.; Ibanescu, R.I.; Kramer, V.G.; Moisi, D.; Asahchop, E.L.; Brenner, B.G.; Harrigan, P.R.; Xu, H.; Wainberg, M.A. Basis for early and preferential selection of the E138K mutation in HIV-1 reverse transcriptase. Antimicrob. Agents Chemother. 2013, 57, 4681–4688. [Google Scholar] [CrossRef] [PubMed]
- Angel, J.B.; Freilich, J.; Arthurs, E.; Ban, J.K.; Lachaine, J.; Chounta, V.; Harris, M. Adherence to oral antiretroviral therapy in Canada, 2010–2020. AIDS 2023, 37, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- van Welzen, B.J.; Van Lelyveld, S.F.L.; Ter Beest, G.; Gisolf, J.H.; Geerlings, S.E.; Prins, J.M.; Van Twillert, G.; Van Nieuwkoop, C.; Van der Valk, M.; Burger, D.; et al. Virological failure after switch to long-acting cabotegravir and rilpivirine injectable therapy: An in-depth analysis. Clin. Infect. Dis. 2024, 79, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Maruapula, D.; Moraka, N.O.; Bareng, O.T.; Mokgethi, P.T.; Choga, W.T.; Seatla, K.K.; Kelentse, N.; Koofhethille, C.K.; Zuze, B.J.L.; Gaolathe, T.; et al. Archived rilpivirine-associated resistance mutations among ART-naive and virologically suppressed people living with HIV-1 subtype C in Botswana: Implications for cabotegravir/rilpivirine use. J. Antimicrob. Chemother. 2023, 78, 2489–2495. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.G.; Oliveira, M.; Ibanescu, R.I.; Routy, J.P.; Thomas, R. Cell culture selections reveal favourable drug resistance profiles for doravirine and islatravir. J. Antimicrob. Chemother. 2021, 76, 2137–2142. [Google Scholar] [CrossRef]
- Brenner, B.G.; Oliveira, M.; Ibanescu, R.I.; Routy, J.P.; Thomas, R. Doravirine responses to HIV-1 viruses bearing mutations to NRTIs and NNRTIs under in vitro selective drug pressure. J. Antimicrob. Chemother. 2023, 78, 1921–1928. [Google Scholar] [CrossRef]
- Charpentier, C.; Storto, A.; Soulie, C.; Ferre, V.M.; Wirden, M.; Joly, V.; Lambert-Niclot, S.; Palich, R.; Morand-Joubert, L.; Landman, R.; et al. Prevalence of genotypic baseline risk factors for cabotegravir + rilpivirine failure among ARV-naive patients. J. Antimicrob. Chemother. 2021, 76, 2983–2987. [Google Scholar] [CrossRef]
- Orkin, C.; Schapiro, J.M.; Perno, C.F.; Kuritzkes, D.R.; Patel, P.; DeMoor, R.; Dorey, D.; Wang, Y.; Han, K.; Van Eygen, V.; et al. Expanded Multivariable Models to Assist Patient Selection for Long-Acting Cabotegravir + Rilpivirine Treatment: Clinical Utility of a Combination of Patient, Drug Concentration, and Viral Factors Associated With Virologic Failure. Clin. Infect. Dis. 2023, 77, 1423–1431. [Google Scholar] [CrossRef]
- Wensing, A.M.; Calvez, V.; Ceccherini-Silberstein, F.; Charpentier, C.; Gunthard, H.F.; Paredes, R.; Shafer, R.W.; Richman, D.D. 2022 update of the drug resistance mutations in HIV-1. Top. Antivir. Med. 2022, 30, 559–574. [Google Scholar] [PubMed]
- Planinic, A.; Begovac, J.; Rokic, F.; Simicic, P.; Oroz, M.; Jakovac, K.; Vugrek, O.; Zidovec-Lepej, S. Characterization of Human Immunodeficiency Virus-1 Transmission Clusters and Transmitted Drug-Resistant Mutations in Croatia from 2019 to 2022. Viruses 2023, 15, 2408. [Google Scholar] [CrossRef] [PubMed]
- Cutrell, A.G.; Schapiro, J.M.; Perno, C.F.; Kuritzkes, D.R.; Quercia, R.; Patel, P.; Polli, J.W.; Dorey, D.; Wang, Y.; Wu, S.; et al. Exploring predictors of HIV-1 virologic failure to long-acting cabotegravir and rilpivirine: A multivariable analysis. AIDS 2021, 35, 1333–1342. [Google Scholar] [CrossRef]
- Steegen, K.; Chandiwana, N.; Sokhela, S.; Venter, W.D.F.; Hans, L. Impact of rilpivirine cross-resistance on long-acting cabotegravir-rilpivirine in low and middle-income countries. AIDS 2023, 37, 1009–1011. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance-Associated Mutations | Acquired Drug Resistance (ADR) in ART-Experienced Persons (VL > 400 Copies/mL) a | Transmitted Resistance in ART-Naïve Persons c | |||
|---|---|---|---|---|---|
| Subtype B 1st ART Failure (n = 3500) | Subtype B CT Failures (n = 6013) b | Non-B-Subtype 1st ART Failure (n = 488) | Subtype B TDR (n = 6011) | Non-B-Subtype PDR (n = 1572) | |
| Any RAMs | 58.5 | 71.2 | 36.7 | 14.7 | 13.9 |
| PI RAMs | 29.5 | 39.8 | 8.2 | 2.4 | 2.0 |
| NRTI RAMs | 38.1 | 62.0 | 27.5 | 5.9 | 6.2 |
| M184V | 42.2 | 50.0 | 23.6 | 1.8 | 3.3 |
| TAMs | 34.9 | 47.9 | 16.0 | 4.5 | 4.2 |
| K65R | 1.5 | 1.9 | 1.8 | 0.1 | 0.1 |
| NNRTI RAMs | 20.5 | 39.2 | 25.8 | 8.8 | 9.4 |
| K103N | 20.2 | 27.2 | 15.8 | 4.3 | 5.7 |
| G190A | 6.9 | 8.8 | 4.1 | 2.8 | 1.2 |
| E138K | 0.6 | 0.4 | 1.0 | 0.2 | 0.4 |
| E138A | 2.5 | 3.2 | 4.9 | 2.7 | 5.1 |
| Drug Class RAMs | Specific RAMs | Cluster Group | All Subtype B (n = 6011) | Χ2 p-Value Exact Test Odds Ratio (Large Cluster vs. Singleton Transmission) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Singleton (n = 1804) | 2–5 Members (n = 1168) | 6+ Members (n = 3039) | |||||||||
| N | % | N | % | N | % | N | % | p Value | Odds Ratio | ||
| Any | 244 | 13.5 | 165 | 14.1 | 472 | 15.5 | 881 | 14.7 | ns * | ns | |
| NRTIs | Any NRTI | 152 | 8.4 | 72 | 6.1 | 131 | 4.3 | 355 | 5.9 | <0.0001 | 0.48 [0.39–0.62] |
| NRTI | M184I,V | 60 | 3.3 | 19 | 1.6 | 29 | 1.0 | 108 | 1.8 | <0.0001 | 0.28 [0.18–0.44] |
| TAMs | M41L | 44 | 2.4 | 31 | 2.7 | 39 | 1.3 | 114 | 1.9 | <0.001 | 0.52 [0.34–0.80] |
| D67N,G,E | 22 | 1.2 | 20 | 1.7 | 7 | 0.2 | 49 | 0.8 | <0.0001 | 0.18 [0.08–0.42] | |
| K70R,E | 19 | 1.1 | 2 | 0.2 | 4 | 0.3 | 25 | 0.4 | <0.0001 | 0.13 [0.04–0.34] | |
| L210W | 21 | 1.4 | 8 | 0.7 | 8 | 0.3 | 37 | 0.6 | <0.001 | 0.22 [0.10–0.51] | |
| T215 revertants | 77 | 4.3 | 47 | 4.0 | 73 | 2.4 | 197 | 3.2 | <0.001 | 0.57 [0.40–0.76] | |
| K219Q,E,N,R | 29 | 1.6 | 27 | 2.3 | 8 | 0.3 | 64 | 1.4 | <0.0001 | 0.16 [4008–0.36] | |
| NNRTIs | Any NNRTI | 136 | 7.5 | 28 | 2.4 | 366 | 12.0 | 530 | 8.8 | <0.0001 | 1.66 [1.35–2.04] |
| NNRTIs | K103N,S * | 76 | 4.2 | 48 | 4.1 | 132 | 4.3 | 256 | 4.3 | ns | ns |
| G190A,S,E | 8 | 0.4 | 10 | 0.9 | 153 | 5.0 | 171 | 2.8 | <0.0001 | 8.70 [5.3–14.4] | |
| Y181C,I,V | 14 | 0.8 | 7 | 0.6 | 11 | 0.4 | 32 | 0.5 | ns | ns | |
| E138A,G,Q,K | 45 | 2.5 | 46 | 3.9 | 94 | 3.1 | 185 | 3.1 | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brenner, B.G.; Ibanescu, R.-I.; Oliveira, M.; Margaillan, G.; Lebouché, B.; Thomas, R.; Baril, J.G.; Lorgeoux, R.-P.; Roger, M.; Routy, J.-P.; et al. Phylogenetic Network Analyses Reveal the Influence of Transmission Clustering on the Spread of HIV Drug Resistance in Quebec from 2002 to 2022. Viruses 2024, 16, 1230. https://doi.org/10.3390/v16081230
Brenner BG, Ibanescu R-I, Oliveira M, Margaillan G, Lebouché B, Thomas R, Baril JG, Lorgeoux R-P, Roger M, Routy J-P, et al. Phylogenetic Network Analyses Reveal the Influence of Transmission Clustering on the Spread of HIV Drug Resistance in Quebec from 2002 to 2022. Viruses. 2024; 16(8):1230. https://doi.org/10.3390/v16081230
Chicago/Turabian StyleBrenner, Bluma G., Ruxandra-Ilinca Ibanescu, Maureen Oliveira, Guillaume Margaillan, Bertrand Lebouché, Réjean Thomas, Jean Guy Baril, René-Pierre Lorgeoux, Michel Roger, Jean-Pierre Routy, and et al. 2024. "Phylogenetic Network Analyses Reveal the Influence of Transmission Clustering on the Spread of HIV Drug Resistance in Quebec from 2002 to 2022" Viruses 16, no. 8: 1230. https://doi.org/10.3390/v16081230