
A Non-Hemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs against the Contemporary Pandemic Genotype II African Swine Fever Virus
 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Animals
2.3. Virus Passage, Titration, and Replication Test In Vitro
2.4. ASFV Genome Next-Generation Sequencing
2.5. Safety, Efficacy, and Duration of Protection Testing of VNUA-ASFV-LAVL3 in Pigs
2.6. Quantitative PCR (qPCR) for ASFV
2.7. Detection of ASFV-Specific Antibody and Cellular Responses in Pigs
2.8. Statistical Analysis
3. Results
3.1. VNUA-ASFV-LAVL3 Loses Hemadsorption Activity during Adaptation in 3D4/21 Cells
3.2. Genetic Characterization of VNUA-ASFV-LAVL3
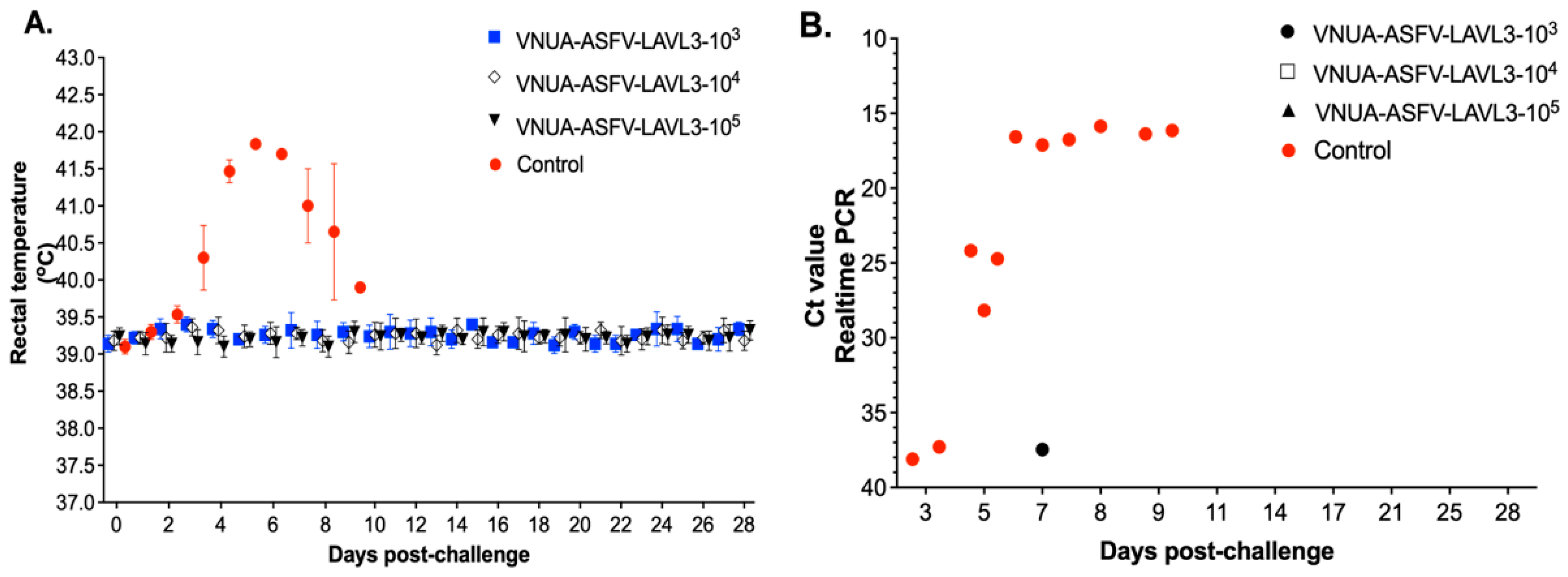
3.3. VNUA-ASFV-LAVL3 Exhibits High Safety in Pigs
3.4. Vaccination with VNUA-ASFV-LAVL3 Confers Full Protection in Pigs
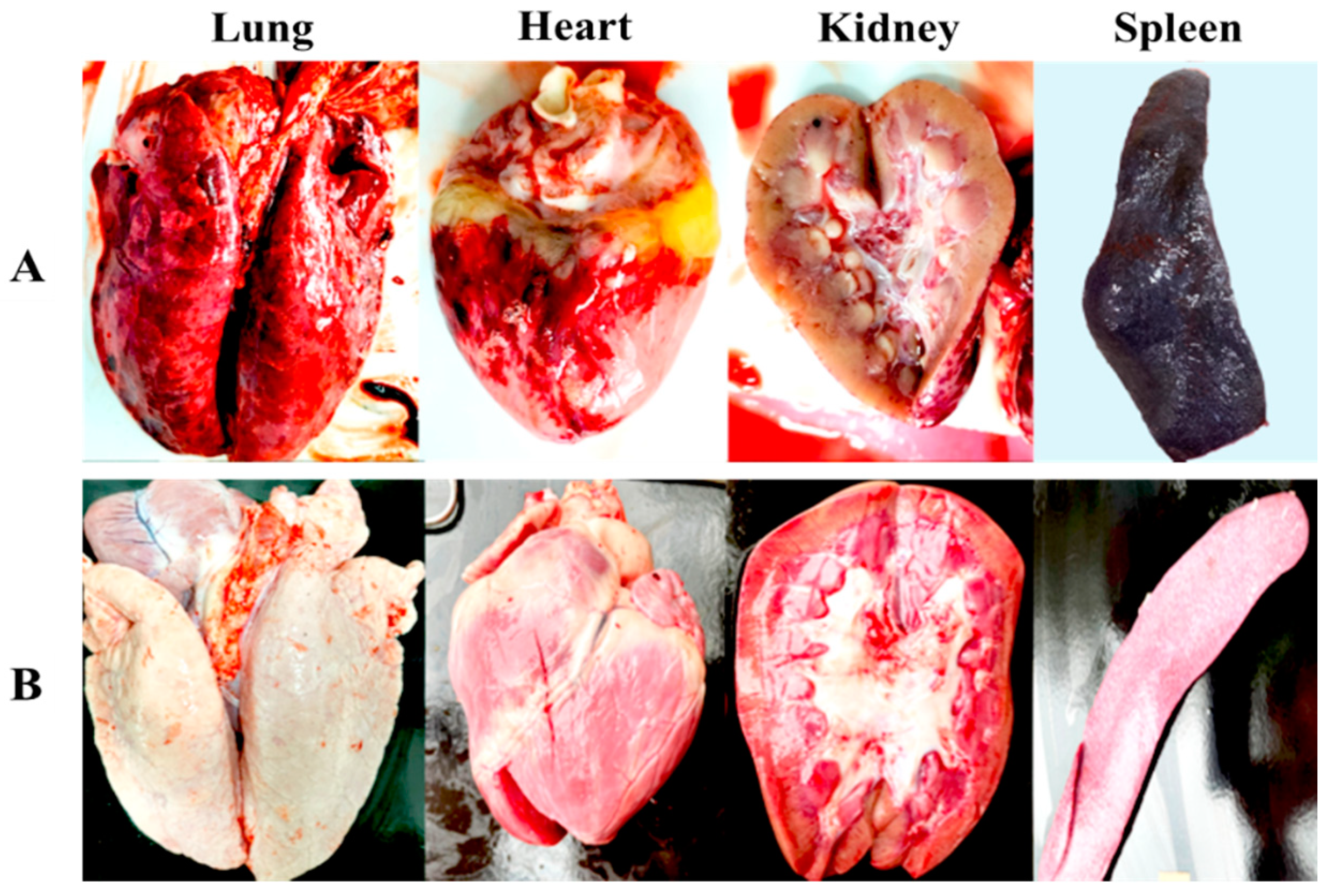
3.5. VNUA-ASFV-LAVL3 Vaccination Prevents Replication of Wild-Type ASFV and Protects against Pathological Lesions in Pigs
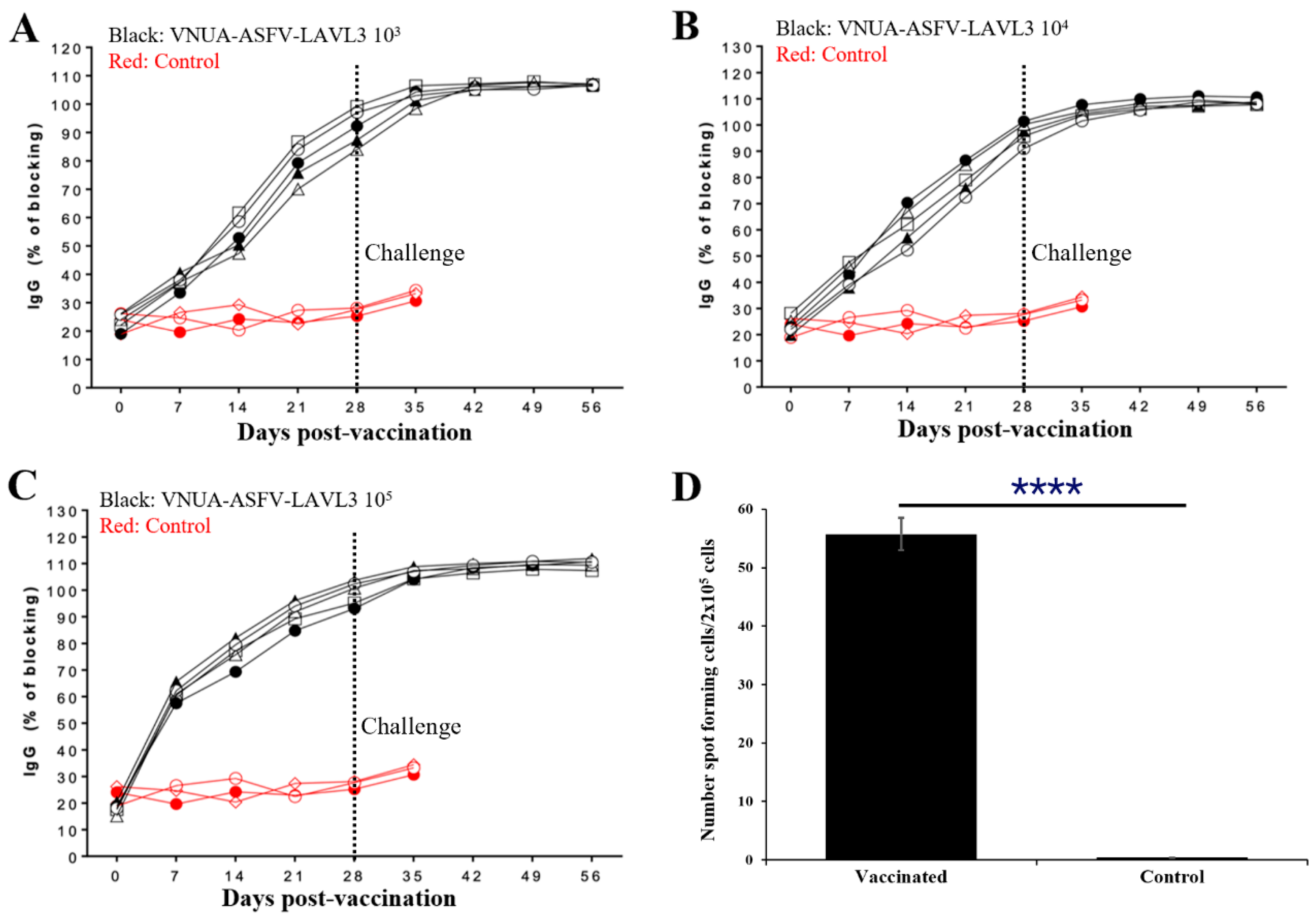
3.6. Pigs with VNUA-ASFV-LAVL3 Induce Robust ASFV-Specific Humoral and Cellular Immune Responses
3.7. Pigs Can Be Protected against Wild-Type ASFV Challenge for Up to 2 Months after a Single Vaccination with VNUA-ASFV-LAVL3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African swine fever epidemiology and control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef]
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; et al. African Swine Fever Virus: A Review. Life 2022, 12, 1255. [Google Scholar] [CrossRef] [PubMed]
- Karger, A.; Pérez-Núñez, D.; Urquiza, J.; Hinojar, P.; Alonso, C.; Freitas, F.B.; Revilla, Y.; Le Potier, M.F.; Montoya, M. An Update on African Swine Fever Virology. Viruses 2019, 11, 864. [Google Scholar] [CrossRef]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef]
- Ruedas-Torres, I.; Thi To Nga, B.; Salguero, F.J. Pathogenicity and virulence of African swine fever virus. Virulence 2024, 15, 2375550. [Google Scholar] [CrossRef]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An update on the epidemiology and pathology of African swine fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernández-Pinero, J.; Sánchez-Vizcaíno, J.M. Gaps in African swine fever: Analysis and priorities. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 235–247. [Google Scholar] [CrossRef] [PubMed]
- Mighell, E.; Ward, M.P. African Swine Fever spread across Asia, 2018–2019. Transbound. Emerg. Dis. 2021, 68, 2722–2732. [Google Scholar] [CrossRef] [PubMed]
- Le, V.P.; Jeong, D.G.; Yoon, S.W.; Kwon, H.M.; Trinh, T.B.N.; Nguyen, T.L.; Bui, T.T.N.; Oh, J.; Kim, J.B.; Cheong, K.M.; et al. Outbreak of African Swine Fever, Vietnam, 2019. Emerg. Infect. Dis. 2019, 25, 1433–1435. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Nguyen, H.T.; Dang, H.V. Genetic characterization of African swine fever viruses circulating in North Central region of Vietnam. Transbound. Emerg. Dis. 2021, 68, 1697–1699. [Google Scholar] [CrossRef]
- Berends, J.; Bendita da Costa Jong, J.; Cooper, T.L.; Dizyee, K.; Morais, O.; Pereira, A.; Smith, D.; Rich, K.M. Investigating the Socio-Economic and Livelihoods Impacts of African Swine Fever in Timor-Leste: An Application of Spatial Group Model Building. Front. Veter-Sci. 2021, 8, 687708. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Cho, K.H.; Mai, N.T.A.; Park, J.Y.; Trinh, T.B.N.; Jang, M.K.; Nguyen, T.T.H.; Vu, X.D.; Nguyen, T.L.; Nguyen, V.D.; et al. Multiple variants of African swine fever virus circulating in Vietnam. Arch. Virol. 2022, 167, 1137–1140. [Google Scholar] [CrossRef] [PubMed]
- Jean-Pierre, R.P.; Hagerman, A.D.; Rich, K.M. An analysis of African Swine Fever consequences on rural economies and smallholder swine producers in Haiti. Front. Veter-Sci. 2022, 9, 960344. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Medina, E.; O’Donnell, V.; Silva, E.; Espinoza, N.; Velazquez-Salinas, L.; Moran, K.; Daite, D.A.; Barrette, R.; Faburay, B.; Holland, R.; et al. Experimental Infection of Domestic Pigs with an African Swine Fever Virus Field Strain Isolated in 2021 from the Dominican Republic. Viruses 2022, 14, 1090. [Google Scholar] [CrossRef] [PubMed]
- Pavone, S.; Iscaro, C.; Dettori, A.; Feliziani, F. African Swine Fever: The State of the Art in Italy. Animals 2023, 13, 2998. [Google Scholar] [CrossRef]
- Brellou, G.D.; Tassis, P.D.; Apostolopoulou, E.P.; Fortomaris, P.D.; Leontides, L.S.; Papadopoulos, G.A.; Tzika, E.D. Report on the First African Swine Fever Case in Greece. Veter Sci. 2021, 8, 163. [Google Scholar] [CrossRef]
- Wang, L.; Ganges, L.; Dixon, L.K.; Bu, Z.; Zhao, D.; Truong, Q.L.; Richt, J.A.; Jin, M.; Netherton, C.L.; Benarafa, C.; et al. 2023 International African Swine Fever Workshop: Critical Issues That Need to Be Addressed for ASF Control. Viruses 2023, 16, 4. [Google Scholar] [CrossRef]
- Shi, J.; Wang, L.; McVey, D.S. Of pigs and men: The best-laid plans for prevention and control of swine fevers. Anim. Front. 2021, 11, 6–13. [Google Scholar] [CrossRef]
- Pikalo, J.; Porfiri, L.; Akimkin, V.; Roszyk, H.; Pannhorst, K.; Kangethe, R.T.; Wijewardana, V.; Sehl-Ewert, J.; Beer, M.; Cattoli, G.; et al. Vaccination with a Gamma Irradiation-Inactivated African Swine Fever Virus Is Safe But Does Not Protect Against a Challenge. Front. Immunol. 2022, 13, 832264. [Google Scholar] [CrossRef]
- Rock, D.L. Thoughts on African Swine Fever Vaccines. Viruses 2021, 13, 943. [Google Scholar] [CrossRef] [PubMed]
- Gaudreault, N.N.; Richt, J.A. Subunit Vaccine Approaches for African Swine Fever Virus. Vaccines 2019, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Jones, J.O.; Andino, R. Rationalizing the development of live attenuated virus vaccines. Nat. Biotechnol. 2010, 28, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.; Franzoni, G.; Oggiano, A. Cell Lines for the Development of African Swine Fever Virus Vaccine Candidates: An Update. Vaccines 2022, 10, 707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Z.; Ge, S.; Zuo, Y.; Lu, H.; Lv, Y.; Han, N.; Cai, Y.; Wu, X.; Wang, Z. Attenuated African swine fever virus through serial passaging of viruses in cell culture: A brief review on the knowledge gathered during 60 years of research. Virus Genes 2023, 59, 13–24. [Google Scholar] [CrossRef]
- Truong, Q.L.; Wang, L.; Nguyen, T.A.; Nguyen, H.T.; Tran, S.D.; Vu, A.T.; Le, A.D.; Nguyen, V.G.; Hoang, P.T.; Nguyen, Y.T.; et al. A Cell-Adapted Live-Attenuated Vaccine Candidate Protects Pigs against the Homologous Strain VNUA-ASFV-05L1, a Representative Strain of the Contemporary Pandemic African Swine Fever Virus. Viruses 2023, 15, 2089. [Google Scholar] [CrossRef]
- Brake, D.A. African Swine Fever Modified Live Vaccine Candidates: Transitioning from Discovery to Product Development through Harmonized Standards and Guidelines. Viruses 2022, 14, 2619. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Camós, L.; López, E.; Rodriguez, F. African swine fever vaccines: A promising work still in progress. Porc. Health Manag. 2020, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Luo, R.; Sun, Y.; Qiu, H.J. Current efforts towards safe and effective live attenuated vaccines against African swine fever: Challenges and prospects. Infect. Dis. Poverty 2021, 10, 137. [Google Scholar] [CrossRef]
- Muñoz-Pérez, C.; Jurado, C.; Sánchez-Vizcaíno, J.M. African swine fever vaccine: Turning a dream into reality. Transbound. Emerg. Dis. 2021, 68, 2657–2668. [Google Scholar] [CrossRef]
- Liu, L.; Wang, X.; Mao, R.; Zhou, Y.; Yin, J.; Sun, Y.; Yin, X. Research progress on live attenuated vaccine against African swine fever virus. Microb. Pathog. 2021, 158, 105024. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of the A137R Gene from the Pandemic Strain of African Swine Fever Virus Attenuates the Strain and Offers Protection against the Virulent Pandemic Virus. J. Virol. 2021, 95, e0113921. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African Swine Fever Virus Bearing an I226R Gene Deletion Elicits Robust Immunity in Pigs to African Swine Fever. J. Virol. 2021, 95, e0119921. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of MGF360 and MGF505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef]
- Rathakrishnan, A.; Reis, A.L.; Petrovan, V.; Goatley, L.C.; Moffat, K.; Lui, Y.; Vuong, M.T.; Ikemizu, S.; Davis, S.J.; Dixon, L.K. A protective multiple gene-deleted African swine fever virus genotype II, Georgia 2007/1, expressing a modified non-haemadsorbing CD2v protein. Emerg. Microbes Infect. 2023, 12, 2265661. [Google Scholar] [CrossRef]
- Borca, M.V.; Rai, A.; Ramirez-Medina, E.; Silva, E.; Velazquez-Salinas, L.; Vuono, E.; Pruitt, S.; Espinoza, N.; Gladue, D.P. A Cell Culture-Adapted Vaccine Virus against the Current African Swine Fever Virus Pandemic Strain. J. Virol. 2021, 95, e0012321. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- de León, P.; Bustos, M.J.; Carrascosa, A.L. Laboratory methods to study African swine fever virus. Virus Res. 2013, 173, 168–179. [Google Scholar] [CrossRef]
- Truong, Q.L.; Nguyen, T.L.; Nguyen, T.H.; Shi, J.; Vu, H.L.X.; Lai, T.L.H.; Nguyen, V.G. Genome Sequence of a Virulent African Swine Fever Virus Isolated in 2020 from a Domestic Pig in Northern Vietnam. Microbiol. Resour. Announc. 2021, 10, e00193-21. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent end points. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kim, J.I.; Yi, G. geneCo: A visualized comparative genomic method to analyze multiple genome structures. Bioinformatics 2019, 35, 5303–5305. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chen, H.; Liu, L.; Cao, Y.; Jiang, T.; Zou, Y.; Peng, Y. Classification and characterization of multigene family proteins of African swine fever viruses. Brief Bioinform. 2021, 22, bbaa380. [Google Scholar] [CrossRef] [PubMed]
- Haines, F.J.; Hofmann, M.A.; King, D.P.; Drew, T.W.; Crooke, H.R. Development and validation of a multiplex, real-time RT PCR assay for the simultaneous detection of classical and African swine fever viruses. PLoS ONE 2013, 8, e71019. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Yáñez, R.J.; Almazán, F.; Viñuela, E.; Rodriguez, J.F. African swine fever virus encodes a CD2 homolog responsible for the adhesion of erythrocytes to infected cells. J. Virol. 1993, 67, 5312–5320. [Google Scholar] [CrossRef]
- Salguero, F.J. Comparative Pathology and Pathogenesis of African Swine Fever Infection in Swine. Front. Veter-Sci. 2020, 7, 282. [Google Scholar] [CrossRef]
- Chu, X.; Ge, S.; Zuo, Y.; Cui, J.; Sha, Z.; Han, N.; Wu, B.; Ni, B.; Zhang, H.; Lv, Y.; et al. Thoughts on the research of African swine fever live-attenuated vaccines. Vaccine, 2024, in press. [CrossRef]
- Leitão, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.M.E.; Portugal, F.C.; Vigário, J.D.; Martins, C.L.V. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. J. Gen. Virol. 2001, 82 Pt 3, 513–523. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Rodze, I.; Nieto, R.; Cano-Gómez, C.; Fernandez-Pinero, J.; Arias, M. Attenuated and non-haemadsorbing (non-HAD) genotype II African swine fever virus (ASFV) isolated in Europe, Latvia 2017. Transbound. Emerg. Dis. 2019, 66, 1399–1404. [Google Scholar] [CrossRef]
- Orosco, F.L. Host immune responses against African swine fever virus: Insights and challenges for vaccine development. Open Veter J. 2023, 13, 1517–1535. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, H.H.; Denyer, M.S.; Lacasta, A.; Stirling, C.M.; Argilaguet, J.M.; Netherton, C.L.; Oura, C.A.; Martins, C.; Rodríguez, F. Cellular immunity in ASFV responses. Virus Res. 2013, 173, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Pan, I.C.; Huang, T.S.; Hess, W.R. New method of antibody detection by indirect immunoperoxidase plaque staining for serodiagnosis of African swine fever. J. Clin. Microbiol. 1982, 16, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.L.; Parkhouse, R.M.E.; Penedos, A.R.; Martins, C.; Leitão, A. Systematic analysis of longitudinal serological responses of pigs infected experimentally with African swine fever virus. J. Gen. Virol. 2007, 88 Pt 9, 2426–2434. [Google Scholar] [CrossRef]
- Luong, H.Q.; Lai, H.T.L.; Truong, L.Q.; Nguyen, T.N.; Vu, H.D.; Nguyen, H.T.; Nguyen, L.T.; Pham, T.H.; McVey, D.S.; Vu, H.L.X. Comparative Analysis of Swine Antibody Responses following Vaccination with Live-Attenuated and Killed African Swine Fever Virus Vaccines. Vaccines 2023, 11, 1687. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Sun, E.; Huang, L.; Ding, L.; Zhu, Y.; Zhang, J.; Shen, D.; Zhang, X.; Zhang, Z.; Ren, T.; et al. Highly lethal genotype I and II recombinant African swine fever viruses detected in pigs. Nat. Commun. 2023, 14, 3096. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Position | Mutation Type | Gene | Nucleotide Change | Amino Acid Change | Protein Change |
|---|---|---|---|---|---|
| 6343 to 6344 | Insertion | Uncharacterized | 61nt | Uncharacterized | Uncharacterized |
| 6366 | Substitution | Uncharacterized | T to A | Uncharacterized | Uncharacterized |
| 6371 | Substitution | Uncharacterized | G to A | Uncharacterized | Uncharacterized |
| 8555 to 8718 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 8719 to 9003 | Deletion | 285L | Deletion | Deletion | Deletion |
| 9004 to 9131 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 9132 to 9515 | Deletion | MGF110-8L | Deletion | Deletion | Deletion |
| 9516 to 9732 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 9733 to 10107 | Deletion | MGF_100-1R | Deletion | Deletion | Deletion |
| 10108 to 10265 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 10266 to 11138 | Deletion | MGF_100-9L | Deletion | Deletion | Deletion |
| 11139 to 11428 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 11429 to 11788 | Deletion | MGF110-11L | Deletion | Deletion | Deletion |
| 11789 to 11877 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 11878 to 12243 | Deletion | MGF110-14L | Deletion | Deletion | Deletion |
| 12244 to 12432 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 12433 to 12792 | Deletion | MGF110-12L | Deletion | Deletion | Deletion |
| 12793 to 12878 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 12879 to 13702 | Deletion | MGF110-13L | Deletion | Deletion | Deletion |
| 13703 to 13882 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 13883 to 15046 | Deletion | MGF360-4L | Deletion | Deletion | Deletion |
| 15047 to 15862 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 15863 to 16990 | Deletion | MGF360-6L | Deletion | Deletion | Deletion |
| 16991 to 17868 | Deletion | Uncharacterized | Deletion | Deletion | Deletion |
| 17869 to 17972 | Deletion | X69R | 104 nt | 34 aa | Shorter protein |
| 20076 | Deletion | MGF_300-2R | C | D to I | Disruption |
| 20188 | Substitution | MGF_300-2R | T to C | S to N | Disruption |
| 23952–23962 | Deletion | Uncharacterized | 11 nt | Uncharacterized | Uncharacterized |
| 24103 | Substitution | MGF_360-10L | T to C | D to Q | Disruption |
| 24752 | Deletion | MGF_360-10L | A | D to E | Disruption |
| 28221–28324 | Deletion | MGF_360-12L | 104 nt | 34aa | Shorter protein |
| 30324 | Deletion | Uncharacterized | A | Uncharacterized | Uncharacterized |
| 30871 | Substitution | MGF_360-14L | G to A | H to A | Abnormal protein |
| 30876–30877 | Substitution | MGF_360-14L | TC to AT | I to S | Abnormal protein |
| 30879 | Substitution | MGF_360-14L | C to T | D to V | Abnormal protein |
| 30882–30915 | Deletion | MGF_360-14L | 34 nt | 11 aa | Abnormal protein |
| 30918 | Substitution | MGF_360-14L | T to A | N to Y | Abnormal protein |
| 30923 | Substitution | MGF_360-14L | T to A | Y to F | Abnormal protein |
| 30930 | Substitution | MGF_360-14L | T to A | S to C | Abnormal protein |
| 30938 | Substitution | MGF_360-14L | A to C | V to G | Abnormal protein |
| 33291 | Deletion | MGF_505-2R | A | K to T | Shorter protein |
| 36919 | Substitution | MGF_505-5R | T to C | Y to N | Substitution |
| 40351 | Substitution | MGF_505-7R | T to G | V to G | Substitution |
| 41114 | Deletion | Uncharacterized | T | Uncharacterized | Uncharacterized |
| 42931–42947 | Deletion | Uncharacterized | 17 nt | Uncharacterized | Uncharacterized |
| 43100–43133 | Deletion | Uncharacterized | 34 nt | Uncharacterized | Uncharacterized |
| 46011 | Deletion | A104R | A | K to S | Disruption |
| 53169 | Deletion | Uncharacterized | T | Uncharacterized | Uncharacterized |
| 58635 | Substitution | F1055L | T to C | I to W | Disruption |
| 58913 | Substitution | F1055L | T to G | H to L | Disruption |
| 60945–61020 | Deletion | F1055L | 76 nt | 25 aa | Disruption |
| 61819 | Substitution | Uncharacterized | T to C | Uncharacterized | Uncharacterized |
| 72222 | Substitution | EP402R | T to C | C to R | Disruption |
| 72824 | Deletion | EP402R | T | R to E | Disruption |
| 72965 | Deletion | EP402R | C | P to T | Disruption |
| 75240–75320 | Deletion | M1249L | 81 nt | 27 aa | Abnormal protein |
| 77036 | Substitution | M1249L | T to C | M to V | Abnormal protein |
| 78038 | Substitution | M1249L | T to G | N to H | Abnormal protein |
| 85588 | Substitution | C475L | T to G | R to S | Substitution |
| 93313 | Substitution | B962L | A to G | Y to H | Substitution |
| 102005 | Substitution | B385R | A to G | T to A | Substitution |
| 105743–105744 | Insertion | B407L | 40nt | 13 aa | Disruption |
| 105777 | Substitution | B407L | T to C | S to G | Disruption |
| 107968–107969 | Insertion | G1340L | 19 nt | 6 aa | Abnormal protein |
| 109399 | Substitution | G1340L | A to C | V to G | Abnormal protein |
| 114357–114561 | Deletion | G1211R | 205 nt | 68 aa | Disruption |
| 120939–121034 | Deletion | CP2475L | 96 nt | 32 aa | Abnormal protein |
| 122809 | Substitution | CP2475L | T to C | N to S | Abnormal protein |
| 124660 | Substitution | CP530R | A to G | N to D | Substitution |
| 130367–130446 | Deletion | NP1450L | 80 nt | 26 aa | Disruption |
| 131201–131500 | Deletion | NP1450L | 300 nt | 100 aa | Disruption |
| 131815 | Substitution | NP1450L | A to G | I to T | Disruption |
| 136825–136841 | Deletion | D250R | 17 nt | 6 aa | Shorter protein |
| 141015 | Substitution | D1133L | T to C | E to A | Disruption |
| 141043 | Substitution | D1133L | T to G | T to S | Disruption |
| 141285–141420 | Deletion | D1133L | 136 nt | 48 aa | Disruption |
| 145156–145171 | Deletion | S183L | 16 nt | 5 aa | Disruption |
| 145490 | Substitution | S273R | C to A | S to S | None |
| 146353–146375 | Deletion | P1192R | 23 nt | 7 aa | Disruption |
| 153308 | Substitution | H233R | T to C | V to A | Disruption |
| 153598–153665 | Deletion | H233R | 68 | 22 aa | Disruption |
| 154180 | Substitution | H240R | C to T | L to F | Substitution |
| 166366 | Substitution | E248R | A to C | P to P | None |
| 173569–173592 | Deletion | I177L | 24 | 8 aa | Abnormal protein |
| 174320–174320 | Deletion | Uncharacterized | A | Uncharacterized | Uncharacterized |
| 182192 | Deletion | MGF 360-18R | A | E to N | Disruption |
| 182665 | Substitution | MGF 360-18R | T to C | L to N | Disruption |
| Organs | Quantitative ASFV Real-Time PCR | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 103 TCID50 Vaccinated (Pig Number) | 104 TCID50 Vaccinated (Pig Number) | 105 TCID50 Vaccinated (Pig Number) | Control (Pig Number) | |||||||||||||||
| 80 | 81 | 82 | 83 | 84 | 85 | 86 | 87 | 88 | 89 | 90 | 91 | 92 | 93 | 94 | 96 | 97 | 98 | |
| Brain | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Heart | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Lung | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Liver | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Stomach | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Spleen | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Kidney | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Bladder | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Tonsil | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| ILN | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| MLN | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| SLN | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| BM | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + |
| Group | No. of Pigs | Vaccination | Wild-Type ASFV Challenge at | Mean (±SD) Time to Death (Days) | Mean (±SD) Fever Duration (Days) | Survival/Protection Rate (%) | Clinical Signs |
|---|---|---|---|---|---|---|---|
| A | 5 | 104 VNUA-ASFV-LAVL3 | 1-month post-vaccination | 0 | 0 | 5/100 | Healthy |
| 3 | No vaccination, contact | 10.3 (±0.57) | 5.6 (±0.57) | 0/0 | High fever, ASF clinical signs | ||
| B | 3 | No vaccination | 8.3 (±0.57) | 4.3 (±0.57) | 0/0 | High fever, ASF clinical signs | |
| C | 5 | 104 VNUA-ASFV-LAVL3 | 2-month post-vaccination | 0 | 0 | 5/100 | Healthy |
| 3 | No vaccination, contact | 10.3 (±0.57) | 5.6 (±0.57) | 0/0 | High fever, ASF clinical signs | ||
| D | 3 | No vaccination | 8.6 (±0.57) | 4.6 (±0.57) | 0/0 | High fever, ASF clinical signs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, Q.L.; Wang, L.; Nguyen, T.A.; Nguyen, H.T.; Le, A.D.; Nguyen, G.V.; Vu, A.T.; Hoang, P.T.; Le, T.T.; Nguyen, H.T.; et al. A Non-Hemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs against the Contemporary Pandemic Genotype II African Swine Fever Virus. Viruses 2024, 16, 1326. https://doi.org/10.3390/v16081326
Truong QL, Wang L, Nguyen TA, Nguyen HT, Le AD, Nguyen GV, Vu AT, Hoang PT, Le TT, Nguyen HT, et al. A Non-Hemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs against the Contemporary Pandemic Genotype II African Swine Fever Virus. Viruses. 2024; 16(8):1326. https://doi.org/10.3390/v16081326
Chicago/Turabian StyleTruong, Quang Lam, Lihua Wang, Tuan Anh Nguyen, Hoa Thi Nguyen, Anh Dao Le, Giap Van Nguyen, Anh Thi Vu, Phuong Thi Hoang, Trang Thi Le, Huyen Thi Nguyen, and et al. 2024. "A Non-Hemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs against the Contemporary Pandemic Genotype II African Swine Fever Virus" Viruses 16, no. 8: 1326. https://doi.org/10.3390/v16081326