

Predominance of Dengue Virus Serotype-1/Genotype-I in Eastern and Southeastern Ethiopia
 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Methods
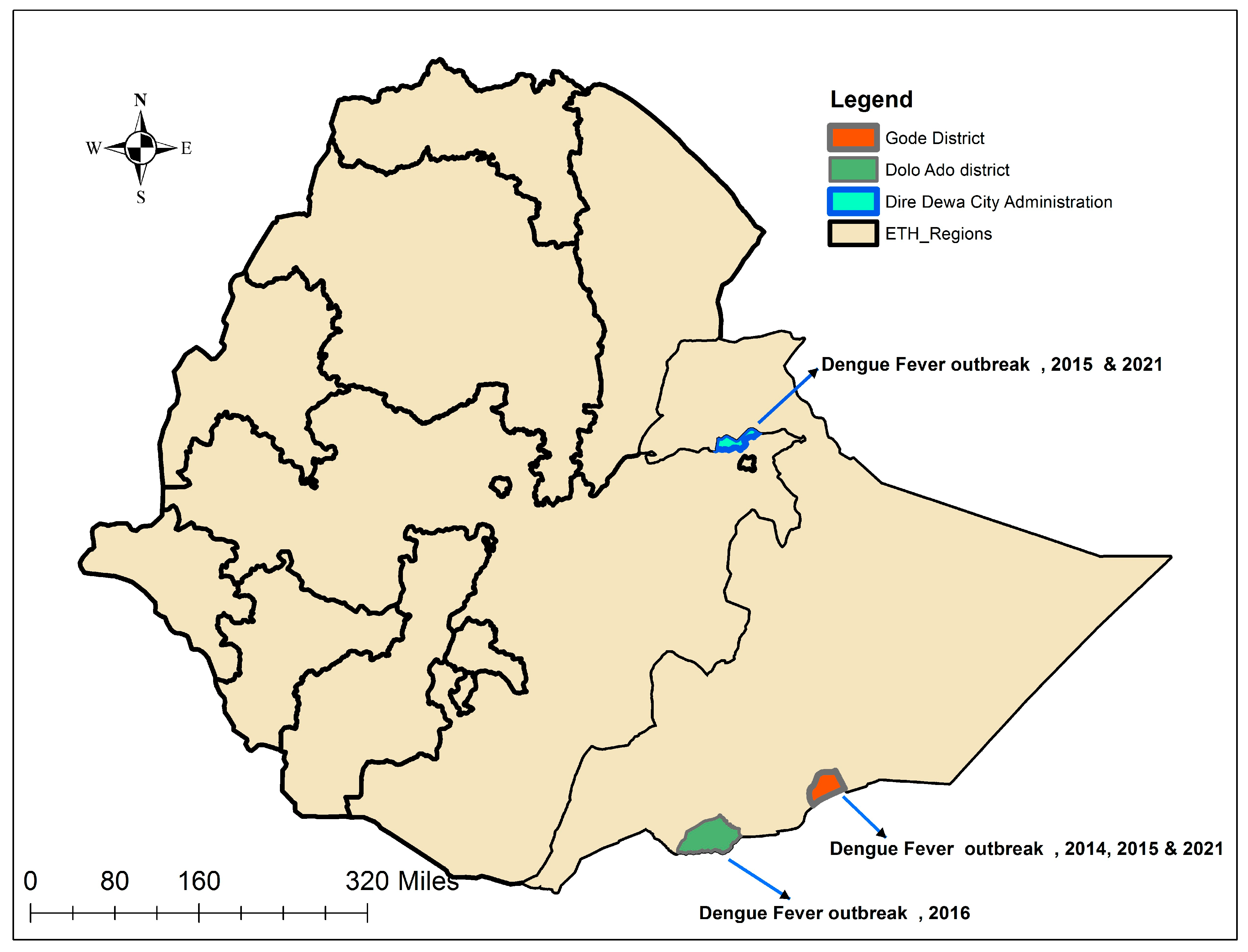
2.1. Source of Samples
2.2. Dengue RNA Amplification
2.3. Sequencing
2.4. Serotyping and Genotyping, and Phylogenetic Analysis
3. Result
3.1. PCR Detection and Sequencing
3.2. Serotyping and Genotyping
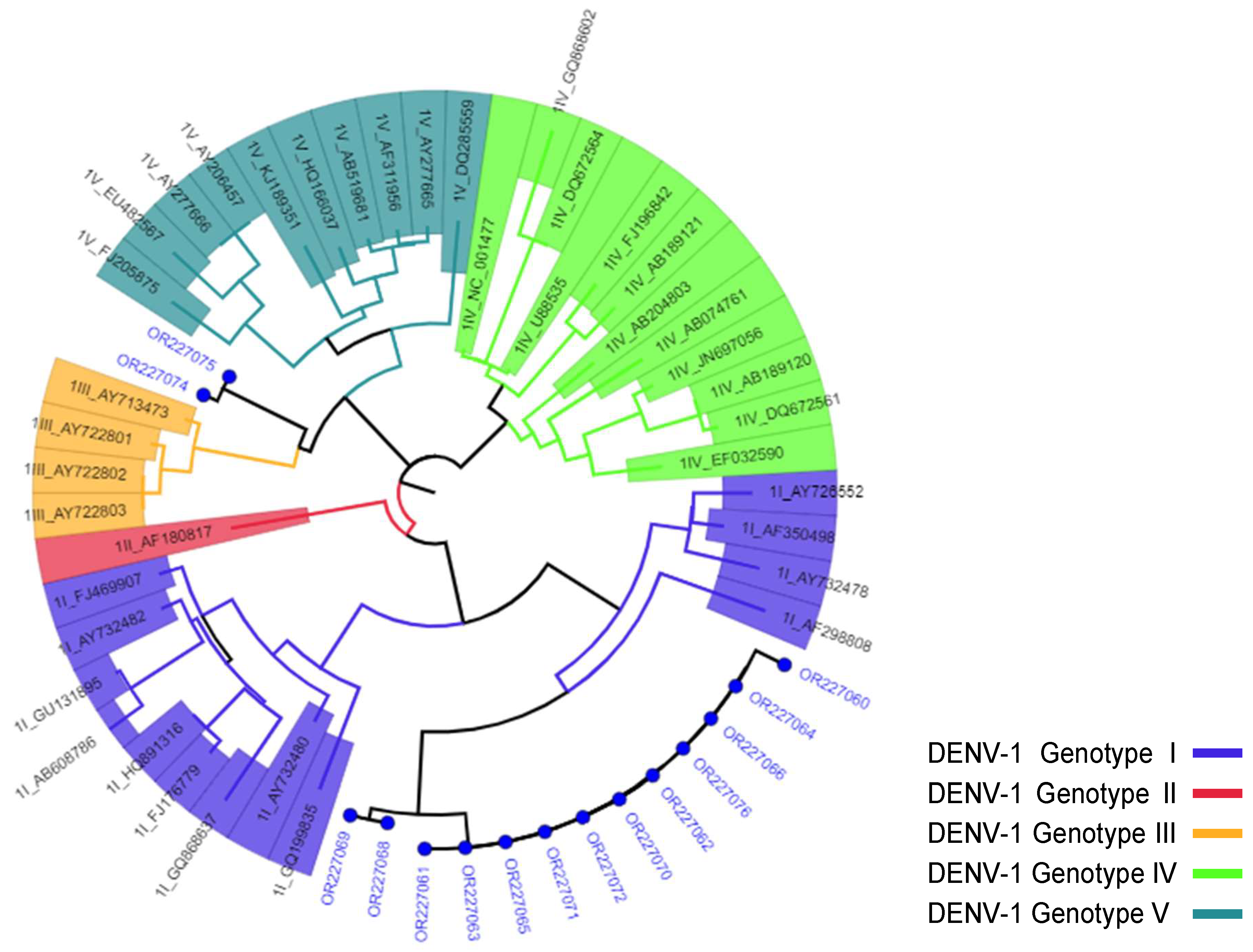
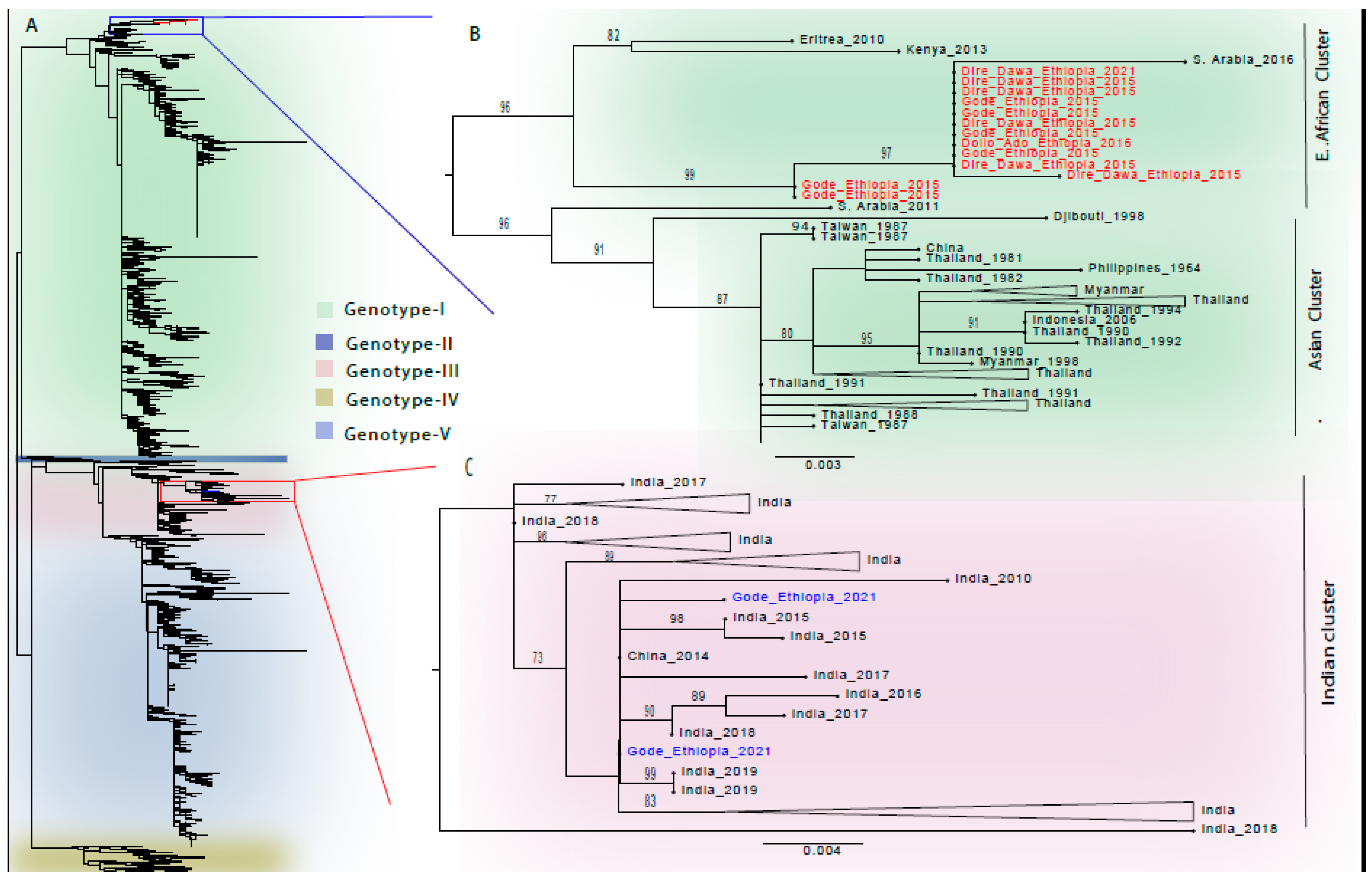
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, S.; Rothman, A. Immunopathological mechanisms in dengue and dengue hemorrhagic fever. Curr. Opin. Infect. Dis. 2006, 19, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C.; Twiddy, S.S. The origin, emergence and evolutionary genetics of dengue virus. Infect. Genet. Evol. 2003, 3, 19–28. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization: WHO. Dengue and Severe Dengue. Fact Sheet. March 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 11 August 2023).
- Eltom, K.; Enan, K.; El Hussein, A.R.M.; Elkhidir, I.M. Dengue Virus Infection in Sub-Saharan Africa Between 2010 and 2020: A Systematic Review and Meta-Analysis. Front. Cell. Infect. Microbiol. 2021, 11, 678945. [Google Scholar] [CrossRef] [PubMed]
- Woyessa, A.B.; Mengesha, M.; Kassa, W.; Kifle, E.; Wondabeku, M.; Girmay, A.; Kebede, A.; Jima, D. The first acute febrile illness investigation associated with dengue fever in Ethiopia, 2013: A descriptive analysis. Ethiop. J. Health Dev. 2014, 28, 155–161. [Google Scholar]
- Amarasinghe, A.; Kuritsky, J.N.; Letson, G.W.; Margolis, H.S. Dengue Virus Infection in Africa. Emerg. Infect. Dis. 2011, 17, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Degife, L.H.; Worku, Y.; Belay, D.; Bekele, A.; Hailemariam, Z. Factors associated with dengue fever outbreak in Dire Dawa administration city, October, 2015, Ethiopia-Case control study. BMC Public Health 2019, 19, 650. [Google Scholar] [CrossRef]
- Mesfin, Z.; Ali, A.; Abagero, A.; Asefa, Z. Dengue Fever Outbreak Investigation in Werder Town, Dollo Zone, Somali Region, Ethiopia. Infect. Drug Resist. 2022, 15, 7207–7217. [Google Scholar] [CrossRef] [PubMed]
- Mekuriaw, W.; Kinde, S.; Kindu, B.; Mulualem, Y.; Hailu, G.; Gebresilassie, A.; Sisay, C.; Bekele, F.; Amare, H.; Wossen, M.; et al. Epidemiological, Entomological, and Climatological Investigation of the 2019 Dengue Fever Outbreak in Gewane District, Afar Region, North-East Ethiopia. Insects 2022, 13, 1066. [Google Scholar] [CrossRef] [PubMed]
- Gutu, M.A.; Bekele, A.; Seid, Y.; Mohammed, Y.; Gemechu, F.; Woyessa, A.B.; Tayachew, A.; Dugasa, Y.; Gizachew, L.; Idosa, M.; et al. Another dengue fever outbreak in Eastern Ethiopia—An emerging public health threat. PLoS Negl. Trop. Dis. 2021, 15, e0008992. [Google Scholar] [CrossRef] [PubMed]
- Gemechu, D.S.; Worku, Y.; Alemu, A.; Edea, Z.A.; Feyisa, Y.D.; Watere, S.H.; Belay, D.; Hagos, A.; Woyessa, A.B. Fourth Dengue Fever Outbreak Investigation in Ethiopia: A Case Control Study. July 2015. Available online: https://www.researchsquare.com/article/rs-4729/v1 (accessed on 29 July 2024).
- Soo, K.M.; Khalid, B.; Ching, S.M.; Chee, H.Y. Meta-analysis of dengue severity during infection by different dengue virus serotypes in primary and secondary infections. PLoS ONE 2016, 11, e0154760. [Google Scholar] [CrossRef]
- Anoop, M.; Issac, A.; Mathew, T.; Philip, S.; Kareem, N.A. Genetic characterization of dengue virus serotypes causing concurrent infection in an outbreak in Ernakulam, Kerala, South India. Indian J. Exp. Biol. 2010, 48, 849–857. [Google Scholar] [PubMed]
- Caroline, A.; Bona, D.; Twerdochlib, A.L. Genetic diversity of dengue virus serotypes 1 and 2 in the State of Paraná, Brazil, based on a fragment of the capsid/premembrane junction region. Rev. Soc. Bras. Med. Trop. 2012, 45, 297–300. [Google Scholar] [CrossRef]
- Luo, Z.; Hu, B.; Zhang, F.; Lin, X.; Xie, X.; Pan, K.; Li, H.Y.; Ren, R.W.; Zhao, W.Z. Laboratory and Molecular Characterization of Dengue Viruses in a 2014 Outbreak in Guangfo Region, Southern China. PLoS Negl. Trop. Dis. 2017, 70, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Falk, K.I.; Alm, E.; Lesko, B.; Lindegren, G.; Ahlm, C.; So, S.; Lagerqvist, N. Universal Single-Probe RT-PCR Assay for Diagnosis of Dengue Virus Infections. PLoS Negl. Trop. Dis. 2014, 8, e3416. [Google Scholar]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.; Libin, P.J.K.; Theys, K.; Faria, N.R.; Nunes, M.R.T.; Restovic, M.I.; Freire, M.; Giovanetti, M.; Cuypers, L.; Nowé, A.; et al. A computational method for the identification of dengue, zika and chikungunya virus species and genotypes. PLoS Negl. Trop. Dis. 2019, 13, e0007231. [Google Scholar] [CrossRef] [PubMed]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.M.; et al. Genome Detective: An automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree. Institute of Evolutionary Biology, University of Edinburgh, Edinburgh. 2010. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 29 July 2024).
- Kalayanarooj, S.; Nimmannitya, S. Clinical and laboratory presentations of dengue patients with different serotypes. WHO Regional Office for South-East Asia. Dengue Bull. 2000, 24, 53–59. Available online: https://iris.who.int/handle/10665/148790 (accessed on 15 December 2023).
- Corwin, A.L.; Larasati, R.P.; Bangs, M.J.; Wuryadi, S.; Arjoso, S.; Sukri, N.; Listyaningsih, E.; Hartati, S.; Namursa, R.; Anwar, Z.; et al. Epidemic dengue transmission in southern Sumatra, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 257–265. [Google Scholar] [CrossRef]
- Andre, G.; Lamballerie XDe Leparc-goffart, I.; Grard, G. Molecular characterization of dengue virus serotype 1 infections in French travelers from Africa between 2013 and 2019. Front. Virol. 2023, 3, 1124848. [Google Scholar] [CrossRef]
- Alfsnes, K.; Eldholm, V.; Gaunt, M.W.; De Lamballerie, X.; Gould, E.A.; Pettersson, J.H. Tracing and tracking the emergence, epidemiology and dispersal of dengue virus to Africa during the 20th century. One Health 2021, 13, 100337. [Google Scholar] [CrossRef] [PubMed]
- Ayolabi, C.I.; Olusola, B.A.; Ibemgbo, S.A.; Okonkwo, G.O. Detection of Dengue viruses among febrile patients in Lagos Nigeria and phylogenetics of circulating Dengue serotypes in Africa. Infect. Genet. Evol. 2019, 75, 103947. [Google Scholar] [CrossRef] [PubMed]
- Yenamandra, S.P.; Koo, C.; Chiang, S.; Shi, H.; Lim, J.; Yeo, Z.Y.; Ng, L.C.; Hapuarachchi, H.C. Evolution, heterogeneity and global dispersal of cosmopolitan genotype of Dengue virus type 2. Sci. Rep. 2021, 11, 13496. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.R.; Herbinger, K.H.; Fröschl, G.; Romano, C.M.; Cabidelle, A.d.S.A.; Junior, C.C. Serotype influences on dengue severity: A cross-sectional study on 485 confirmed dengue cases in Vitória, Brazil. BMC Infect. Dis. 2016, 16, 320. [Google Scholar] [CrossRef]
- Gupta, A.; Rijhwani, P.; Pahadia, M.R.; Kalia, A.; Choudhary, S.; Bansal, D.P.; Gupta, D.; Agarwal, P.; Jat, R.K. Prevalence of Dengue Serotypes and Its Correlation With the Laboratory Profile at a Tertiary Care Hospital in Northwestern India. Cureus 2021, 13, e15029. [Google Scholar] [CrossRef]
- Ahmed, Y.M.; Salah, A.A. Epidemiology of Dengue Fever in Ethiopian Somali Region: Retrospective Health Facility Based Study. Cent. Afr. J. Public Health 2016, 2, 51–56. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Year of Outbreak | District/Region Affected | Serotype/Genotype * |
|---|---|---|---|
| OR227059 | 2014 | Gode/Somali | DENV-2/Cosmopolitan |
| OR227060 | 2015 | Dire Dawa City | DENV-1/Genotype-I |
| OR227061 | 2015 | Dire Dawa City | DENV-1/Genotype-I |
| OR227062 | 2015 | Dire Dawa City | DENV-1/Genotype-I |
| OR227063 | 2015 | Dire Dawa City | DENV-1/Genotype-I |
| OR227064 | 2015 | Dire Dawa City | DENV-1/Genotype-I |
| OR227065 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227066 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227067 | 2015 | Gode/Somali | DENV-2/Cosmopolitan |
| OR227068 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227069 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227070 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227071 | 2015 | Gode/Somali | DENV-1/Genotype-I |
| OR227072 | 2016 | Dollo Ado/Somali | DENV-1/Genotype-I |
| OR227073 | 2016 | Dollo Ado/Somali | DENV-2/Cosmopolitan |
| OR227074 | 2021 | Gode/Somali | DENV-1/Genotype-III |
| OR227075 | 2021 | Gode/Somali | DENV-1/Genotype-III |
| OR227076 | 2021 | Dire Dawa City | DENV-1/Genotype-I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsegaye, M.M.; Mekonnen, A.T.; Gebretsion, D.T.; Gelanew, T.; Alemayehu, D.H.; Tefera, D.A.; Woldemichael, T.S.; Getaneh, B.A.; Abera, E.K.; Jebessa, G.G.; et al. Predominance of Dengue Virus Serotype-1/Genotype-I in Eastern and Southeastern Ethiopia. Viruses 2024, 16, 1334. https://doi.org/10.3390/v16081334
Tsegaye MM, Mekonnen AT, Gebretsion DT, Gelanew T, Alemayehu DH, Tefera DA, Woldemichael TS, Getaneh BA, Abera EK, Jebessa GG, et al. Predominance of Dengue Virus Serotype-1/Genotype-I in Eastern and Southeastern Ethiopia. Viruses. 2024; 16(8):1334. https://doi.org/10.3390/v16081334
Chicago/Turabian StyleTsegaye, Mesfin Mengesha, Adamu Tayachew Mekonnen, Daniel Tsega Gebretsion, Tesfaye Gelanew, Dawit Hailu Alemayehu, Dessalegn Abeje Tefera, Tamirayehu Seyoum Woldemichael, Bethlehem Adnew Getaneh, Eleni Kidane Abera, Gadissa Gutema Jebessa, and et al. 2024. "Predominance of Dengue Virus Serotype-1/Genotype-I in Eastern and Southeastern Ethiopia" Viruses 16, no. 8: 1334. https://doi.org/10.3390/v16081334