
The First Isolation of Insect-Specific Alphavirus (Agua Salud alphavirus) in Culex (Melanoconion) Mosquitoes in the Brazilian Amazon

 ,
,  , , , , , , ,
, , , , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Ethical Consent
2.3. Hematophagous Arthropod Collection and Taxonomic Identification
2.4. Viral Isolation in Cell Cultures and Indirect Immunofluorescence (IIF) Test
2.5. Next-Generation Sequencing (NGS) and Nucleotide Sequence Assembly
2.6. Phylogenetic Inference
3. Results
3.1. Viral Isolation and Indirect Immunofluorescence Test
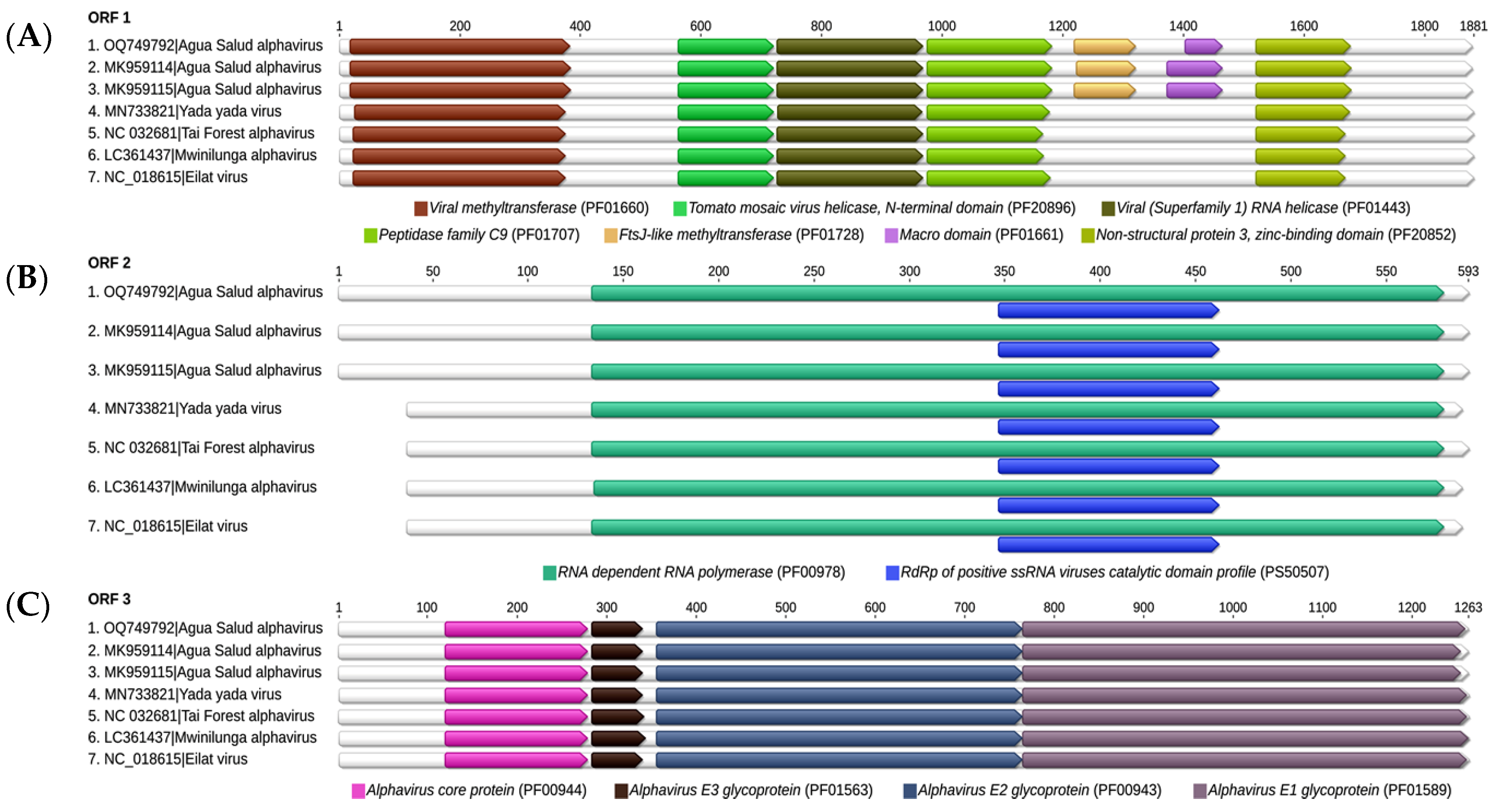
3.2. Nucleotide Sequencing and Phylogenetic Inference
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrade, R.P. “Uma floresta cheia de vírus!” Ciência e desenvolvimento nas fronteiras amazônicas. RBH 2019, 39, 19–42. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Dégallier, N.; Pinheiro, F.P.; Sá Filho, G.C. Epidemiologia das encefalites por arbovírus na Amazônia Brasileira. Rev. Inst. Med. Trop. São Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef]
- Ramos, B.A.; Chagas, L.L.; Silva, F.A.; Santos, E.B.; Chiang, J.O.; Nunes-Neto, J.P.; Vieira, D.B.R.; Rosa-Junior, J.W.; Silva, E.V.P.; Freitas, M.N.O.; et al. Arboviruses in free-ranging birds and hematophagous arthropods (Diptera, Nematocera) from forest remnants and urbanized areas of an Environmental Protect Area in the Amazon biome. Viruses 2022, 14, 2101. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dégallier, N.; Travassos da Rosa, J.F.S. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saúde Pública Rio Janeiro 2001, 17, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Instituto de Desenvolvimento Florestal e da Biodiversidade do Estado do Pará (IDEFLOR-BIO). Região Administrativa de Belém 2021. Available online: https://ideflorbio.pa.gov.br/unidades-de-conservacao/regiao-administrativa-belem/ (accessed on 5 January 2024).
- Nunes, M.R.T.; Silva, S.P.; Carvalho, V.L.; Vasconcelos, J.M.; Silva, D.E.A.; Oliveira, L.F.; Nunes-Neto, J.P.; Rodrigues, S.G.; Azevedo, R.S.S.; Monteiro, H.A.O.; et al. Emergence of new insect restrictive virus in Amazon region. Genome Announc. 2015, 2, e00131-15. [Google Scholar]
- Nunes, M.R.T.; Contreras-Gutierrez, M.A.; Guzman, H.; Martins, L.C.; Barbirato, M.F.; Savit, C.; Balta, V.; Uribe, S.; Viveiro, R.; Suaza, J.D.; et al. Genetic characterization molecular epidemiology and phylogenetic relationships of insect-specific viruses in the táxon Negevirus. Virology 2017, 504, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.S.; Martins, L.C.; Silva, S.P.; Medeiros, D.B.A.; Miranda, K.C.P.; Nunes-Neto, J.P.; Monteiro, H.A.O.; Nascimento, B.L.S.; Rosa-Junior, J.W.; Cruz, A.C.R.; et al. Negeviruses isolated from mosquitoes in the Brazilian Amazon. J. Virol. 2022, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Stollar, V.; Thomas, V.L. An agent in the Aedes aegypti cell line (Peleg) which causes fusion of Aedes albopictus cells. Virology 1975, 64, 367–377. [Google Scholar] [CrossRef]
- Bolling, B.G.; Weaver, S.C.; Tesh, R.B.; Vasilakis, N. Insect-specific virus discovery: Significance for the arbovirus Community. Viruses 2015, 7, 4911–4928. [Google Scholar] [CrossRef]
- Altinli, M.; Schnettler, E.; Sicard, M. Synbiontic interactions between mosquitoes and mosquito viruses. Front. Cell. Infect. Microbiol. 2021, 11, 694020. [Google Scholar] [CrossRef]
- Carvalho, V.L.; Long, M.T. Insect-specific viruses: An overview and their relationship to arboviruses of concern to humans and animals. Virology 2021, 557, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Nasar, F.; Palacios, G.; Gorchakov, R.V.; Guzman, H.; Travassos da Rosa, A.P.A.; Savji, N.; Popov, V.L.; Sherman, M.B.; Lipkin, W.I.; Tesh, R.B.; et al. Eilat virus, a unique alphavirus with host range restricted to insects by RNA replication. Proc. Natl. Acad. Sci. USA 2012, 36, 14622–14627. [Google Scholar] [CrossRef]
- Hermanns, K.; Zirkel, F.; Kopp, A.; Marklewitz, M.; Rwego, I.B.; Estrada, A.; Gillespie, T.R.; Drosten, C.; Junglen, S. Discovery of a novel alphavirus related to Eilat vírus. J. Gen. Virol. 2017, 98, 43–49. [Google Scholar] [CrossRef]
- Torii, S.; Orba, Y.; Hang’Ombe, B.M.; Mweene, A.S.; Wada, Y.; Anindita, P.D.; Phongphaew, W.; Qiu, Y.; Kajihara, M.; Mori-Kajhara, A.; et al. Discovery of Mwinilunga alphavirus: A novel alphavirus in Culex mosquitoes in Zambia. Virus Res. 2018, 250, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Batovska, J.; Buchmann, J.P.; Holmes, E.C.; Lynch, S.E. Coding-complete genome sequencing of Yada yada virus, a novel Alphavirus detected in Australian mosquitoes. Microbiol. Resour. Announc. 2020, 9, e01476-19. [Google Scholar]
- Hermanns, K.; Marklewitz, M.; Zirkel, F.; Overheul, G.J.; Page, R.A.; Loaiza, J.R.; Drosten, C.; Van Rij, R.P.; Junglen, S. Agua Salud alphavirus defines a novel lineage of insect-specific alphaviruses discovered in the New World. J. Gen. Virol. 2020, 101, 96–104. [Google Scholar] [CrossRef]
- Brasil. Guia para planejamento de capturas de anofelinos pela técnica de Atração por Humano Protegido (TAHP) e acompanhamento dos riscos à saúde do profissional capturador; Secretaria de Vigilância em Saúde, Departamento de Imunização e Doenças Transmissíveis, Ministério da Saúde: Brasília, Brasil, 2019. Available online: https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/svsa/malaria/vigilancia-entomologica-e-controle-vetorial-da-malaria/guia-para-o-planejamento-das-acoes-de-captura-de-anofelinos-pela-tecnica-de-atracao-por-humano-protegido-tahp-e-acompanhamento-dos-riscos-a-saude-do-profissional-capturador.pdf (accessed on 8 September 2022).
- Consoli, R.A.G.B.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Fiocruz: Rio de Janeiro, Brasil, 1994; p. 228. [Google Scholar]
- Igarashi, A. Isolation of a singh’s Aedes albopictus cell clone sensitive to Dengue and Chikungunya viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef]
- Rhim, J.S.; Schell, K.; Creasy, B.; Case, W. Biological characteristics and susceptibility of an African green monkey kidney cell line (VERO). Proc. Soc. Exp. Biol. Med. 1962, 132, 670–678. [Google Scholar] [CrossRef]
- Gubler, D.J.; Kuno, G.; Sather, G.E.; Oliver, V.L.A. Mosquito cell culture specific monoclonal antibodies in surveillance for dengue viruses. Am. J. Trop. Med. Hyg. 1984, 1, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.; Yiu, S.M.; Chin, F.Y. IDBA-UD: A DE NOVO assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Myung, I.J. Tutorial in maximum likelihood estimation. J. Math. Psychol. 2003, 47, 90–100. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the Bootstrap. Evolution 1985, 4, 783–791. [Google Scholar] [CrossRef]
- Bennouna, A.; Gil, P.; El Rhaffouli, H.; Exbrayat, A.; Loire, E.; Belenghien, T.; Chlyeh, G.; Gutierrez, S.; Fihri, O.F. Identification of Eilat virus and prevalence of infecction among Culex pipiens L. populations, Marroco, 2016. Virology 2019, 530, 85–88. [Google Scholar] [CrossRef]
- Öhlund, P.; Lunden, H.; Blomström, A.L. Insect-specific virus evolution and potential effects on vector competence. Virus Genes 2019, 55, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Pang, R.; Cheng, T.; Xue, L.; Zeng, H.; Lei, T.; Chen, M.; Wu, S.; Ding, Y.; Zhang, J.; et al. Abundant and diverse RNA viruses in insects revealed by RNA-seq analysis: Ecological and evolutionary implications. MSystems 2020, 5, e00039-20. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C. Febre amarela. Rev. Soc. Bras. Med. Trop. 2003, 36, 275–293. [Google Scholar] [CrossRef]
- Wilkerson, R.C.; Linton, Y.M.; Stickman, D. Mosquitoes of the World, 2nd ed.; Hopkins University Press: Baltimore, MD, USA, 2020; p. 1332. [Google Scholar]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef]
- Faria, I.J.S.; Almeida, J.P.P.; Marques, J.T. Impact of symbiotic insect-specific viruses on mosquito vector competence for arboviruses. Curr. Opin. Insect. Sci. 2024, 63, 101194. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Hanh, C.S.; Lustig, S.; Strauss, E.G.; Strauss, J.H. Western equine encephalitis virus is a recombinant virus. Proc. Natl. Acad. Sci. USA 1988, 85, 5997–6001. [Google Scholar]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribuition. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef] [PubMed]
- Kallies, R.; Kopp, A.; Zirkel, F.; Estrada, A.; Gillespie, T.R.; Drosten, C.; Junglen, S. Genetic characterization of Goutanap virus, a novel virus related to negeviruses, cileviruses and higreviruses. Viruses 2014, 6, 4346–4357. [Google Scholar] [CrossRef]
- Chen, R.; Mukhopadhway, S.; Mertis, A.; Bolling, B.; Nasar, F.; Coffey, L.C.; Powers, A.; Weaver, S.C.; Ictv Report Consortiom. ICTV virus taxonomy profile: Togaviridae. J. Gen. Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef]
- Ferron, F.; Etienne, D.; Selisko, B.; Canard, B. The viral RNA capping machinery as a target for antiviral drugs. Antivir. Res. 2012, 96, 21–31. [Google Scholar] [CrossRef]
- Russo, A.T.; White, M.A.; Watowich, S.J. The crystal structure of the Venezuelan equine encephalitis alphavirus nsP2 protease. Structure 2006, 14, 1449–1458. [Google Scholar] [CrossRef]
- Göetz, G.P.; McNally, K.L.; Robertson, S.J.; Best, S.M.; Pijlman, G.P.; Fros, J.J. The Methyltransferase-like domains of Chikungunya virus nsP2 inhibits the interferon response by promoting the nuclear export of STAT1. J. Virol. 2018, 92, e01008-18. [Google Scholar]
- Powers, A.M.; Brault, A.C.; Shirako, Y.; Strauss, E.G.; Kang, W.; Strauss, J.H.; Weaver, S.C. Evolutionary relationships and systematics of the alphaviruses. J. Virol. 2001, 75, 10118–10131. [Google Scholar] [CrossRef]
- Barbosa, M.G.V.; Fé, N.F.; Marcião, A.H.R.; Silva, A.P.T.; Monteiro, W.M.; Guerra, M.V.F.; Guerra, J.A.O. Registro de Culicidae de importância epidemiológica na área rural de Manaus, Amazonas. Rev. Soc. Bras. Med. Trop. 2008, 41, 658–663. [Google Scholar] [CrossRef]
- Nouri, S.; Matsumura, E.E.; Kuo, Y.W.; Falk, B.W. Insect-specific viruses: From discovery to potential translational applications. Curr. Opin. Virol. 2018, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Nasar, F.; Erasmus, J.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus induces both homologous and heterologous interference. Virology 2015, 484, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Kautz, T.F.; Contreras-Gutierrez, M.A.; Guzman, H.; Tesh, R.B.; Hughes, G.L.; Forrester, N.L. Negeviruses reduces replication of alphaviruses during coinfection. J. Virol. 2021, 95, e00433-21. [Google Scholar] [CrossRef] [PubMed]
- Altinli, M.; Leggewie, M.; Badusche, M.; Gyanwali, R.; Scherer, C.; Schulze, J.; Sreenu, V.B.; Fegebank, M.; Bemhard, Z.; Fuss, J.; et al. Antiviral RNAi response Against insect-specific Agua Salud alphavirus. mSphere 2022, 7, e01003-21. [Google Scholar] [CrossRef]
- Jagtap, S.V.; Brink, J.; Frank, S.C.; Badusche, M.; Leggewie, M.; Sreenu, V.B.; Fuss, J.; Schnettler, E.; Altinli, M. Agua Salud alphavirus infection, dissemination and transmission in Aedes aegypti mosquitões. Viruses 2023, 15, 1113. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Antigenic Group | Virus |
|---|---|---|
| Alphavirus | A | Aura virus (AURAV), EEEV, Mayaro virus (MAYV), MUCV, PIXV, Una virus (UNAV), WEEV, Chikungunya virus (CHIKV), Trocara virus (TROV). |
| Orthoflavivirus | B | Orthoflavivirus denguei (1-4), Orthoflavivirus flavi, Bussuquara virus, Orthoflavivirus cacipacoreense, Orthoflavivirus ilheusenses, Orthoflavivirus nilense, Orthoflavivirus louisense, Orthoflavivirus zikaense. |
| Orthobunyavirus | Guamá | Orthobunyavirus ananindeuaense, Bimiti orthobunyavirus, Orthobunyavirus catuense, Orthobunyavirus guamaense, Orthobunyavirus mirimense, Moju orthobunyavirus, Orthobunyavirus timboteuaense. |
| Capim | Orthobunyavirus acaraense, Benevides orthobunyavirus, Orthobunyavirus benficaense, Orthobunyavirus capimense, Orthobunyavirus guajaraense, Orthobunyavirus bushbushense, Moriche orthobunyavirus. | |
| Bunyamwera | Orthobunyavirus iacoense, Kairi orthobunyavirus, Orthobunyavirus macauaense, Orthobunyavirus maguariense, Sorocaba orthobunyavirus, Tucunduba orthobunyavirus, Taiassui orthobunyavirus, Xingu orthobunyavirus. | |
| Simbu | Orthobunyavirus oropoucheense | |
| Phlebovirus | Phlebotomus | Phlebovirus alenquerense, Phlebovirus ambeense, Ariquemes phlebovirus, Belterra phlebovirus, Phlebovirus bujaruense, Phlebovirus candiruense, Phlebovirus icoaraciense, Joá phlebovirus, Phlebovirus itaporanguense, Jacundá phlebovirus, Morumbi phlebovirus, Mucura phlebovirus, Phlebovirus mugumbaense, Phlebovirus oriximinaense, Pacuí phlebovirus, Phlebovirus saloboense, Phlebovirus taparaense, Phlebovirus turunaense, Phlebovirus uriuranaense, Phlebovirus urucuriense. |
| Orbivirus | Changuinola | Acatinga virus, Acurené virus, Almeirim virus, Altamira virus, Anapú virus, Araçaí virus, Aratau virus, Aruana virus, Arawetê virus, Assurinis virus, Bacajaí virus, Bacuri virus, Balbina virus, Barcarena virus, Breves virus, Canindé virus, Canoal virus, Catetê virus, Orbivirus changuinolaense, Coari virus, Gorotire virus, Gurupi virus, Iopaka virus, Ipixaia virus, Irituia virus, Iruana virus, Itaboca virus, Jamanxi virus, Jandaia virus, Jari virus, Jatuarana virus, Jutaí virus, Kararaô virus, Melgaço virus, Monte Dourado virus, Ourém virus, Pacajá virus, Parakanã virus, Poranati virus, Parauapebas virus, Parú virus, Pependana virus, Pindobai virus, Piratuba virus, Purus virus, Rio Mutapi virus, Saracá virus, Serra Sul virus, Surubim virus, Tapiropé virus, Tekupeú virus, Timbozal virus, Tocantins virus, Tocaxá virus, Tuerê virus, Tumucumaque virus, Uatamã virus, Uxituba virus, Xaraíra virus, Xiwanga virus. |
| Insect-Specific Alphavirus Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | Eilat virus/NC_018615 | 72.27 | 71.87 | 62.05 | 45.12 | 45.12 | 44.72 | Amino acid | |
| 2 | Mwinilunga alphavirus/LC361437 | 69.18 | 75.93 | 63.33 | 46.12 | 46.12 | 46.04 | ||
| 3 | Tai Forest alphavirus/NC_032681 | 66.85 | 68.83 | 62.45 | 46.32 | 46.32 | 46.40 | ||
| 4 | Yada yada virus/MN733821 | 61.55 | 61.88 | 61.62 | 44.56 | 44.56 | 44.24 | ||
| 5 | Agua Salud alphavirus/MK959114 | 53.06 | 52.61 | 53.71 | 53.29 | 91.06 | 96.96 | ||
| 6 | Agua Salud alphavirus/MK959115 | 53.03 | 52.58 | 53.71 | 53.27 | 99.97 | 96.96 | ||
| 7 | Agua Salud alphavirus/OQ749792 | 52.85 | 52.42 | 53.68 | 52.89 | 91.06 | 91.03 | ||
| Nucleotide | |||||||||
| Insect-Specific Alphavirus Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | Eilat virus /NC_018615 | 83.25 | 81.80 | 75.58 | 62.56 | 62.47 | 62.56 | Amino acid | |
| 2 | Mwinilunga alphavirus/LC361437 | 72.87 | 80.62 | 74.33 | 62.55 | 62.51 | 62.59 | ||
| 3 | Tai Forest alphavirus/NC_032681 | 72.32 | 71.39 | 74.19 | 62.15 | 62.15 | 62.34 | ||
| 4 | Yada yada virus/MN733821 | 67.93 | 68.08 | 67.33 | 60.87 | 60.87 | 60.83 | ||
| 5 | Agua Salud alphavirus/MK959114 | 62.17 | 61.42 | 60.94 | 60.15 | 99.88 | 98.19 | ||
| 6 | Agua Salud alphavirus/MK959115 | 62.14 | 61.42 | 60.94 | 60.15 | 99.96 | 98.06 | ||
| 7 | Agua Salud alphavirus/OQ749792 | 62.09 | 61.38 | 60.89 | 60.23 | 89.77 | 89.73 | ||
| Nucleotide | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, B.; Carvalho, V.; da Silva, E.; Freitas, M.; Barros, L.J.; Santos, M.; Pantoja, J.A.; Gonçalves, E.; Nunes Neto, J.; Junior, J.W.; et al. The First Isolation of Insect-Specific Alphavirus (Agua Salud alphavirus) in Culex (Melanoconion) Mosquitoes in the Brazilian Amazon. Viruses 2024, 16, 1355. https://doi.org/10.3390/v16091355
Ramos B, Carvalho V, da Silva E, Freitas M, Barros LJ, Santos M, Pantoja JA, Gonçalves E, Nunes Neto J, Junior JW, et al. The First Isolation of Insect-Specific Alphavirus (Agua Salud alphavirus) in Culex (Melanoconion) Mosquitoes in the Brazilian Amazon. Viruses. 2024; 16(9):1355. https://doi.org/10.3390/v16091355
Chicago/Turabian StyleRamos, Bruna, Valéria Carvalho, Eliana da Silva, Maria Freitas, Landeson Junior Barros, Maissa Santos, Jamilla Augusta Pantoja, Ercília Gonçalves, Joaquim Nunes Neto, José Wilson Junior, and et al. 2024. "The First Isolation of Insect-Specific Alphavirus (Agua Salud alphavirus) in Culex (Melanoconion) Mosquitoes in the Brazilian Amazon" Viruses 16, no. 9: 1355. https://doi.org/10.3390/v16091355