
Cellular N-Myristoyl Transferases Are Required for Mammarenavirus Multiplication
 , , , , ,
, , , , ,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Compounds
2.3. Cell Cytotoxicity Assay and CC50 Determination
2.4. EC50 Determination
2.5. Viral Growth Kinetics
2.6. LCMV Minigenome Assay
2.7. Budding Assay
2.8. Western Blotting
2.9. Virus Titration
2.10. RNA Isolation and Characterization
2.11. GP2-Mediated Fusion Assay
2.12. Assessment of NP to Z Ratio
2.13. Immunofluorescence and Subcellular Localization of NP and Z
2.14. Statistical Analyses
3. Results
3.1. Dose-Dependent Effect of the NMT1/2 Specific Inhibitor DDD85646 on LCMV Multiplication
3.2. Effect of DDD85646 on LCMV Multi-Step Growth Kinetics and Peak Titers
3.3. Effect of DDD85646 on Different Steps of the LCMV Life Cycle
3.4. Effect of DDD85646 on GP2-Mediated Fusion
3.5. Effect of DDD85646 on Z Protein Stability
3.6. Effect of DDD85646 on the Subcellular Distribution of Z Protein
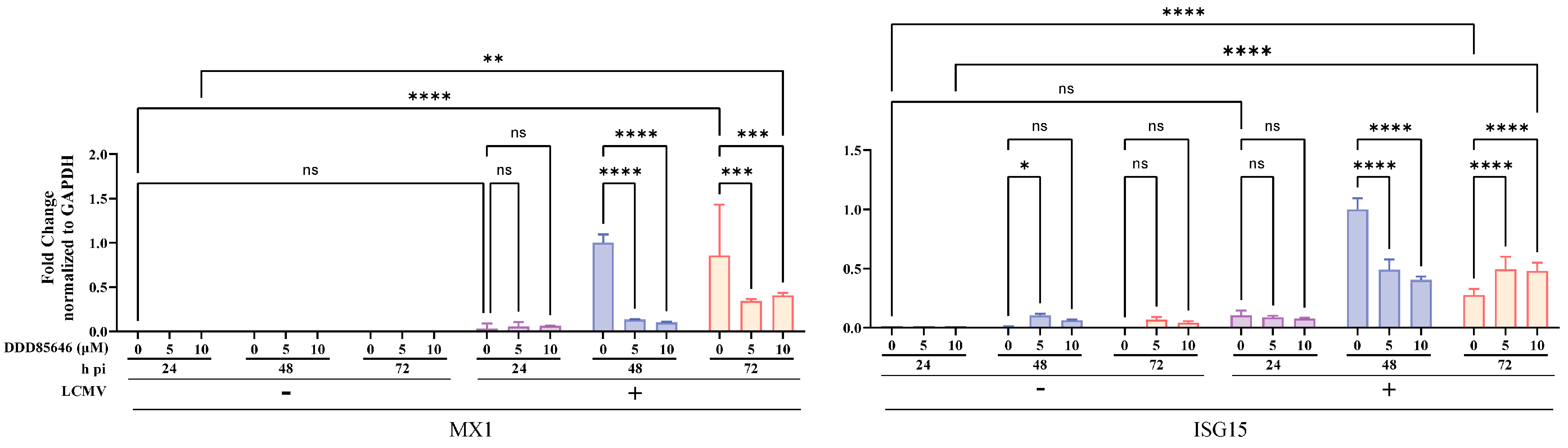
3.7. Contribution of the Type 1 Interferon (T1IFN) Response to DDD85646-Mediated Restricted LCMV Multiplication
3.8. Contribution of NMT1 and NMT2 Isozymes to LCMV Multiplication
3.9. Effect of DDD85646 on the Multiplication of Other Mammarenaviruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Radoshitzky, S.R.; Buchmeier, M.J.; de la Torre, J.C. Fields Virology: Emerging Viruses: Arenaviridae. In Fields Virology, 7th ed.; Vol I: Emerging Viruses; Knipe, D.M., Howley, P.M., Whelan, S., Eds.; Lippincott & Wilkins: Philadelphia, PA, USA, 2022. [Google Scholar]
- Lenz, O.; ter Meulen, J.; Klenk, H.-D.; Seidah, N.G.; Garten, W. The Lassa Virus Glycoprotein Precursor GP-C Is Proteolytically Processed by Subtilase SKI-1/S1P. Proc. Natl. Acad. Sci. USA 2001, 98, 12701–12705. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.R.; Pöpplau, D.; Garten, W.; von Laer, D.; Lenz, O. Endoproteolytic Processing of the Lymphocytic Choriomeningitis Virus Glycoprotein by the Subtilase SKI-1/S1P. J. Virol. 2003, 77, 2866–2872. [Google Scholar] [CrossRef]
- Kunz, S.; Edelmann, K.H.; de la Torre, J.-C.; Gorney, R.; Oldstone, M.B.A. Mechanisms for Lymphocytic Choriomeningitis Virus Glycoprotein Cleavage, Transport, and Incorporation into Virions. Virology 2003, 314, 168–178. [Google Scholar] [CrossRef]
- Rojek, J.M.; Lee, A.M.; Nguyen, N.; Spiropoulou, C.F.; Kunz, S. Site 1 Protease Is Required for Proteolytic Processing of the Glycoproteins of the South American Hemorrhagic Fever Viruses Junin, Machupo, and Guanarito. J. Virol. 2008, 82, 6045–6051. [Google Scholar] [CrossRef] [PubMed]
- Fedeli, C.; Moreno, H.; Kunz, S. Novel Insights into Cell Entry of Emerging Human Pathogenic Arenaviruses. J. Mol. Biol. 2018, 430, 1839–1852. [Google Scholar] [CrossRef] [PubMed]
- Hallam, S.J.; Koma, T.; Maruyama, J.; Paessler, S. Review of Mammarenavirus Biology and Replication. Front. Microbiol. 2018, 9, 1751. [Google Scholar] [CrossRef]
- Cordo, S.M.; Candurra, N.A.; Damonte, E.B. Myristic Acid Analogs Are Inhibitors of Junin Virus Replication. Microbes Infect. 1999, 1, 609–614. [Google Scholar] [CrossRef]
- Perez, M.; Greenwald, D.L.; de La Torre, J.C. Myristoylation of the RING Finger Z Protein Is Essential for Arenavirus Budding. J. Virol. 2004, 78, 11443–11448. [Google Scholar] [CrossRef]
- York, J.; Nunberg, J.H. Role of the Stable Signal Peptide of Junín Arenavirus Envelope Glycoprotein in pH-Dependent Membrane Fusion. J. Virol. 2006, 80, 7775–7780. [Google Scholar] [CrossRef]
- Kallemeijn, W.W.; Lueg, G.A.; Faronato, M.; Hadavizadeh, K.; Goya Grocin, A.; Song, O.-R.; Howell, M.; Calado, D.P.; Tate, E.W. Validation and Invalidation of Chemical Probes for the Human N-Myristoyltransferases. Cell Chem. Biol. 2019, 26, 892–900.e4. [Google Scholar] [CrossRef]
- Corbic Ramljak, I.; Stanger, J.; Real-Hohn, A.; Dreier, D.; Wimmer, L.; Redlberger-Fritz, M.; Fischl, W.; Klingel, K.; Mihovilovic, M.D.; Blaas, D.; et al. Cellular N-Myristoyltransferases Play a Crucial Picornavirus Genus-Specific Role in Viral Assembly, Virion Maturation, and Infectivity. PLoS Pathog. 2018, 14, e1007203. [Google Scholar] [CrossRef] [PubMed]
- Frearson, J.A.; Brand, S.; McElroy, S.P.; Cleghorn, L.A.T.; Smid, O.; Stojanovski, L.; Price, H.P.; Guther, M.L.S.; Torrie, L.S.; Robinson, D.A.; et al. N-Myristoyltransferase Inhibitors as New Leads to Treat Sleeping Sickness. Nature 2010, 464, 728–732. [Google Scholar] [CrossRef] [PubMed]
- York, J.; Romanowski, V.; Lu, M.; Nunberg, J.H. The Signal Peptide of the Junín Arenavirus Envelope Glycoprotein Is Myristoylated and Forms an Essential Subunit of the Mature G1–G2 Complex. J. Virol. 2004, 78, 10783–10792. [Google Scholar] [CrossRef]
- York, J.; Nunberg, J.H. Myristoylation of the Arenavirus Envelope Glycoprotein Stable Signal Peptide Is Critical for Membrane Fusion but Dispensable for Virion Morphogenesis. J. Virol. 2016, 90, 8341–8350. [Google Scholar] [CrossRef]
- Timms, R.T.; Zhang, Z.; Rhee, D.Y.; Harper, J.W.; Koren, I.; Elledge, S.J. A Glycine-Specific N-Degron Pathway Mediates the Quality Control of Protein N-Myristoylation. Science 2019, 365, eaaw4912. [Google Scholar] [CrossRef]
- Andersen, K.G.; Shapiro, B.J.; Matranga, C.B.; Sealfon, R.; Lin, A.E.; Moses, L.M.; Folarin, O.A.; Goba, A.; Odia, I.; Ehiane, P.E.; et al. Clinical Sequencing Uncovers Origins and Evolution of Lassa Virus. Cell 2015, 162, 738–750. [Google Scholar] [CrossRef]
- Richmond, J.K.; Baglole, D.J. Lassa Fever: Epidemiology, Clinical Features, and Social Consequences. BMJ 2003, 327, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Yun, N.E.; Walker, D.H. Pathogenesis of Lassa Fever. Viruses 2012, 4, 2031–2048. [Google Scholar] [CrossRef]
- Radoshitzky, S.R.; Buchmeier, M.J.; de la Torre, J.C. Fields Virology: Emerging Viruses. In Arenaviridae: The Viruses and Their Replication, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D., Lamb, R., Martin, M., Roizman, B., Straus, S., Eds.; Lippincott Williams & Wilkins (LWW): Philadelphia, PA, USA, 2007; Volume II. [Google Scholar]
- Bray, M. Pathogenesis of Viral Hemorrhagic Fever. Curr. Opin. Immunol. 2005, 17, 399–403. [Google Scholar] [CrossRef]
- Grant, A.; Seregin, A.; Huang, C.; Kolokoltsova, O.; Brasier, A.; Peters, C.; Paessler, S. Junín Virus Pathogenesis and Virus Replication. Viruses 2012, 4, 2317–2339. [Google Scholar] [CrossRef]
- Salam, A.P.; Cheng, V.; Edwards, T.; Olliaro, P.; Sterne, J.; Horby, P. Time to Reconsider the Role of Ribavirin in Lassa Fever. PLoS Neglected Trop. Dis. 2021, 15, e0009522. [Google Scholar] [CrossRef]
- Gowen, B.B.; Juelich, T.L.; Sefing, E.J.; Brasel, T.; Smith, J.K.; Zhang, L.; Tigabu, B.; Hill, T.E.; Yun, T.; Pietzsch, C.; et al. Favipiravir (T-705) Inhibits Junín Virus Infection and Reduces Mortality in a Guinea Pig Model of Argentine Hemorrhagic Fever. PLoS Negl. Trop. Dis. 2013, 7, e2614. [Google Scholar] [CrossRef]
- Safronetz, D.; Rosenke, K.; Westover, J.B.; Martellaro, C.; Okumura, A.; Furuta, Y.; Geisbert, J.; Saturday, G.; Komeno, T.; Geisbert, T.W.; et al. The Broad-Spectrum Antiviral Favipiravir Protects Guinea Pigs from Lethal Lassa Virus Infection Post-Disease Onset. Sci. Rep. 2015, 5, 14775. [Google Scholar] [CrossRef]
- Mendenhall, M.; Russell, A.; Smee, D.F.; Hall, J.O.; Skirpstunas, R.; Furuta, Y.; Gowen, B.B. Effective Oral Favipiravir (T-705) Therapy Initiated after the Onset of Clinical Disease in a Model of Arenavirus Hemorrhagic Fever. PLoS Neglected Trop. Dis. 2011, 5, e1342. [Google Scholar] [CrossRef] [PubMed]
- Cashman, K.A.; Smith, M.A.; Twenhafel, N.A.; Larson, R.A.; Jones, K.F.; Allen, R.D.; Dai, D.; Chinsangaram, J.; Bolken, T.C.; Hruby, D.E.; et al. Evaluation of Lassa Antiviral Compound ST-193 in a Guinea Pig Model. Antiviral Res. 2011, 90, 70–79. [Google Scholar] [CrossRef]
- Sangha, R.S.; Jamal, R.; Spratlin, J.L.; Kuruvilla, J.; Sehn, L.H.; Weickert, M.; Berthiaume, L.G.; Mackey, J.R. A First-in-Human, Open-Label, Phase I Trial of Daily Oral PCLX-001, an NMT Inhibitor, in Patients with Relapsed/Refractory B-Cell Lymphomas and Advanced Solid Tumors. JCO 2023, 41, e15094. [Google Scholar] [CrossRef]
- Sangha, R.; Davies, N.M.; Namdar, A.; Chu, M.; Spratlin, J.; Beauchamp, E.; Berthiaume, L.G.; Mackey, J.R. Novel, First-in-Human, Oral PCLX-001 Treatment in a Patient with Relapsed Diffuse Large B-Cell Lymphoma. Curr. Oncol. 2022, 29, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, M.; Minder, P.; Caì, Y.; Kuhn, J.H.; Yates, J.R.; Torbett, B.E.; de la Torre, J.C. Interactome Analysis of the Lymphocytic Choriomeningitis Virus Nucleoprotein in Infected Cells Reveals ATPase Na+/K+ Transporting Subunit Alpha 1 and Prohibitin as Host-Cell Factors Involved in the Life Cycle of Mammarenaviruses. PLoS Pathog. 2018, 14, e1006892. [Google Scholar] [CrossRef] [PubMed]
- Emonet, S.F.; Seregin, A.V.; Yun, N.E.; Poussard, A.L.; Walker, A.G.; de la Torre, J.C.; Paessler, S. Rescue from Cloned cDNAs and In Vivo Characterization of Recombinant Pathogenic Romero and Live-Attenuated Candid #1 Strains of Junin Virus, the Causative Agent of Argentine Hemorrhagic Fever Disease. J. Virol. 2011, 85, 1473–1483. [Google Scholar] [CrossRef]
- Ye, C.; de la Torre, J.C.; Martínez-Sobrido, L. Development of Reverse Genetics for the Prototype New World Mammarenavirus Tacaribe Virus. J. Virol. 2020, 94, e01014-20. [Google Scholar] [CrossRef]
- Lukashevich, I.S.; Paessler, S.; de la Torre, J.C. Lassa Virus Diversity and Feasibility for Universal Prophylactic Vaccine. F1000Research 2019, 8. F1000 Faculty Rev-134. [Google Scholar] [CrossRef] [PubMed]
- Witwit, H.; Khafaji, R.; Salaniwal, A.; Kim, A.S.; Cubitt, B.; Jackson, N.; Ye, C.; Weiss, S.R.; Martinez-Sobrido, L.; de la Torre, J.C. Activation of Protein Kinase Receptor (PKR) Plays a pro-Viral Role in Mammarenavirus-Infected Cells. J. Virol. 2024, 98, e01883-23. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.B.; de la Torre, J.C. Rescue of the Prototypic Arenavirus LCMV Entirely from Plasmid. Virology 2006, 350, 370–380. [Google Scholar] [CrossRef]
- Battegay, M.; Cooper, S.; Althage, A.; Bänziger, J.; Hengartner, H.; Zinkernagel, R.M. Quantification of Lymphocytic Choriomeningitis Virus with an Immunological Focus Assay in 24- or 96-Well Plates. J. Virol. Methods 1991, 33, 191–198. [Google Scholar] [CrossRef]
- Perez, M.; de la Torre, J.C. Characterization of the Genomic Promoter of the Prototypic Arenavirus Lymphocytic Choriomeningitis Virus. J. Virol. 2003, 77, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Capul, A.A.; de la Torre, J.C. A Cell-Based Luciferase Assay Amenable to High-Throughput Screening of Inhibitors of Arenavirus Budding. Virology 2008, 382, 107–114. [Google Scholar] [CrossRef]
- McCausland, M.M.; Crotty, S. Quantitative PCR (QPCR) Technique for Detecting Lymphocytic Choriomeningitis Virus (LCMV) in Vivo. J. Virol. Methods 2008, 147, 167–176. [Google Scholar] [CrossRef]
- Paape, D.; Bell, A.S.; Heal, W.P.; Hutton, J.A.; Leatherbarrow, R.J.; Tate, E.W.; Smith, D.F. Using a Non-Image-Based Medium-Throughput Assay for Screening Compounds Targeting N-Myristoylation in Intracellular Leishmania Amastigotes. PLoS Negl. Trop. Dis. 2014, 8, e3363. [Google Scholar] [CrossRef]
- Perez, M.; Craven, R.C.; de la Torre, J.C. The Small RING Finger Protein Z Drives Arenavirus Budding: Implications for Antiviral Strategies. Proc. Natl. Acad. Sci. USA 2003, 100, 12978–12983. [Google Scholar] [CrossRef] [PubMed]
- Urata, S.; Ngo, N.; de la Torre, J.C. The PI3K/Akt Pathway Contributes to Arenavirus Budding. J. Virol. 2012, 86, 4578–4585. [Google Scholar] [CrossRef]
- Borrow, P.; Oldstone, M.B. Mechanism of Lymphocytic Choriomeningitis Virus Entry into Cells. Virology 1994, 198, 1–9. [Google Scholar] [CrossRef]
- Castilla, V.; Mersich, S.E.; Candurra, N.A.; Damonte, E.B. The Entry of Junin Virus into Vero Cells. Arch. Virol. 1994, 136, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Radoshitzky, S.R.; Abraham, J.; Spiropoulou, C.F.; Kuhn, J.H.; Nguyen, D.; Li, W.; Nagel, J.; Schmidt, P.J.; Nunberg, J.H.; Andrews, N.C.; et al. Transferrin Receptor 1 Is a Cellular Receptor for New World Haemorrhagic Fever Arenaviruses. Nature 2007, 446, 92–96. [Google Scholar] [CrossRef]
- Ma, X.-Z.; Bartczak, A.; Zhang, J.; Khattar, R.; Chen, L.; Liu, M.F.; Edwards, A.; Levy, G.; McGilvray, I.D. Proteasome Inhibition In Vivo Promotes Survival in a Lethal Murine Model of Severe Acute Respiratory Syndrome. J. Virol. 2010, 84, 12419–12428. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, S.; Sabo, Y.; Wang, C.; Tong, L.; Goff, S.P. Heme Oxygenase 2 Binds Myristate to Regulate Retrovirus Assembly and TLR4 Signaling. Cell Host Microbe 2017, 21, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dai, T.; Sun, W.; Wei, Y.; Ren, J.; Zhang, L.; Zhang, M.; Zhou, F. Protein N-Myristoylation: Functions and Mechanisms in Control of Innate Immunity. Cell Mol. Immunol. 2021, 18, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Vlahakis, J.Z.; Vukomanovic, D.; Nakatsu, K.; Szarek, W.A. Selective Inhibition of Heme Oxygenase-2 Activity by Analogs of 1-(4-Chlorobenzyl)-2-(Pyrrolidin-1-Ylmethyl)-1H-Benzimidazole (Clemizole): Exploration of the Effects of Substituents at the N-1 Position. Bioorg. Med. Chem. 2013, 21, 6788–6795. [Google Scholar] [CrossRef]
- Floresta, G.; Fallica, A.N.; Romeo, G.; Sorrenti, V.; Salerno, L.; Rescifina, A.; Pittalà, V. Identification of a Potent Heme Oxygenase-2 (HO-2) Inhibitor by Targeting the Secondary Hydrophobic Pocket of the HO-2 Western Region. Bioorg. Chem. 2020, 104, 104310. [Google Scholar] [CrossRef]
- Kong, X.; Vukomanovic, D.; Nakatsu, K.; Szarek, W.A. Structure-Activity Relationships of 1,2-Disubstituted Benzimidazoles: Selective Inhibition of Heme Oxygenase-2 Activity. ChemMedChem 2015, 10, 1435–1441. [Google Scholar] [CrossRef]
- Intagliata, S.; Salerno, L.; Ciaffaglione, V.; Leonardi, C.; Fallica, A.N.; Carota, G.; Amata, E.; Marrazzo, A.; Pittalà, V.; Romeo, G. Heme Oxygenase-2 (HO-2) as a Therapeutic Target: Activators and Inhibitors. Eur. J. Med. Chem. 2019, 183, 111703. [Google Scholar] [CrossRef]
- Strecker, T.; Maisa, A.; Daffis, S.; Eichler, R.; Lenz, O.; Garten, W. The Role of Myristoylation in the Membrane Association of the Lassa Virus Matrix Protein Z. Virol. J. 2006, 3, 93. [Google Scholar] [CrossRef]
- Thinon, E.; Serwa, R.A.; Broncel, M.; Brannigan, J.A.; Brassat, U.; Wright, M.H.; Heal, W.P.; Wilkinson, A.J.; Mann, D.J.; Tate, E.W. Global Profiling of Co- and Post-Translationally N-Myristoylated Proteomes in Human Cells. Nat. Commun. 2014, 5, 4919. [Google Scholar] [CrossRef]
- Tsumagari, K.; Isobe, Y.; Ishihama, Y.; Seita, J.; Arita, M.; Imami, K. Application of Liquid-Liquid Extraction for N-Terminal Myristoylation Proteomics. Mol. Cell. Proteom. 2023, 22, 100677. [Google Scholar] [CrossRef] [PubMed]
- Takamitsu, E.; Otsuka, M.; Haebara, T.; Yano, M.; Matsuzaki, K.; Kobuchi, H.; Moriya, K.; Utsumi, T. Identification of Human N-Myristoylated Proteins from Human Complementary DNA Resources by Cell-Free and Cellular Metabolic Labeling Analyses. PLoS ONE 2015, 10, e0136360. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Y.; Wang, J.; Qin, D.; Wang, M.; Chai, L.; Fu, Y.; Zhao, C.; Gao, C.; Jia, J.; et al. Myristic Acid as a Checkpoint to Regulate STING-Dependent Autophagy and Interferon Responses by Promoting N-Myristoylation. Nat. Commun. 2023, 14, 660. [Google Scholar] [CrossRef] [PubMed]
- Barr, S.D.; Smiley, J.R.; Bushman, F.D. The Interferon Response Inhibits HIV Particle Production by Induction of TRIM22. PLoS Pathog. 2008, 4, e1000007. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sobrido, L.; Emonet, S.; Giannakas, P.; Cubitt, B.; García-Sastre, A.; de la Torre, J.C. Identification of Amino Acid Residues Critical for the Anti-Interferon Activity of the Nucleoprotein of the Prototypic Arenavirus Lymphocytic Choriomeningitis Virus. J. Virol. 2009, 83, 11330–11340. [Google Scholar] [CrossRef]
- Takamune, N.; Gota, K.; Misumi, S.; Tanaka, K.; Okinaka, S.; Shoji, S. HIV-1 Production Is Specifically Associated with Human NMT1 Long Form in Human NMT Isozymes. Microbes Infect. 2008, 10, 143–150. [Google Scholar] [CrossRef]
- Immerheiser, M.; Zimniak, M.; Hilpert, H.; Geiger, N.; König, E.-M.; Bodem, J. Towards a Broad-Spectrum Antiviral, the Myristoyltransferase Inhibitor IMP-1088 Suppresses Viral Replication—The Yellow Fever NS5 Is Myristoylated. bioRxiv 2021. bioRxiv:2021.03.09.434547. [Google Scholar]
- Mousnier, A.; Bell, A.S.; Swieboda, D.P.; Morales-Sanfrutos, J.; Pérez-Dorado, I.; Brannigan, J.A.; Newman, J.; Ritzefeld, M.; Hutton, J.A.; Guedán, A.; et al. Fragment-Derived Inhibitors of Human N-Myristoyltransferase Block Capsid Assembly and Replication of the Common Cold Virus. Nat. Chem. 2018, 10, 599–606. [Google Scholar] [CrossRef]
- Baillet, N.; Krieger, S.; Carnec, X.; Mateo, M.; Journeaux, A.; Merabet, O.; Caro, V.; Tangy, F.; Vidalain, P.-O.; Baize, S. E3 Ligase ITCH Interacts with the Z Matrix Protein of Lassa and Mopeia Viruses and Is Required for the Release of Infectious Particles. Viruses 2020, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Szulc, N.A.; Stefaniak, F.; Piechota, M.; Soszyńska, A.; Piórkowska, G.; Cappannini, A.; Bujnicki, J.M.; Maniaci, C.; Pokrzywa, W. DEGRONOPEDIA: A Web Server for Proteome-Wide Inspection of Degrons. Nucleic Acids Res. 2024, 52, W221–W232. [Google Scholar] [CrossRef]
- Hickey, C.M.; Breckel, C.; Zhang, M.; Theune, W.C.; Hochstrasser, M. Protein Quality Control Degron-Containing Substrates Are Differentially Targeted in the Cytoplasm and Nucleus by Ubiquitin Ligases. Genetics 2020, 217, iyaa031. [Google Scholar] [CrossRef]
- Veits, G.K.; Henderson, C.S.; Vogelaar, A.; Eron, S.J.; Lee, L.; Hart, A.; Deibler, R.W.; Baddour, J.; Elam, W.A.; Agafonov, R.V.; et al. Development of an AchillesTAG Degradation System and Its Application to Control CAR-T Activity. Curr. Res. Chem. Biol. 2021, 1, 100010. [Google Scholar] [CrossRef]
- Yeh, C.; Huang, W.; Hsu, P.; Yeh, K.; Wang, L.; Hsu, P.W.; Lin, H.; Chen, Y.; Chen, S.; Yeang, C.; et al. The C-degron Pathway Eliminates Mislocalized Proteins and Products of Deubiquitinating Enzymes. EMBO J. 2021, 40, e105846. [Google Scholar] [CrossRef] [PubMed]
- Damhofer, H.; Radzisheuskaya, A.; Helin, K. Generation of Locus-Specific Degradable Tag Knock-Ins in Mouse and Human Cell Lines. STAR Protoc. 2021, 2, 100575. [Google Scholar] [CrossRef]
- Cornu, T.I.; de la Torre, J.C. RING Finger Z Protein of Lymphocytic Choriomeningitis Virus (LCMV) Inhibits Transcription and RNA Replication of an LCMV S-Segment Minigenome. J. Virol. 2001, 75, 9415–9426. [Google Scholar] [CrossRef]
- Thinon, E.; Morales-Sanfrutos, J.; Mann, D.J.; Tate, E.W. N-Myristoyltransferase Inhibition Induces ER-Stress, Cell Cycle Arrest, and Apoptosis in Cancer Cells. ACS Chem. Biol. 2016, 11, 2165–2176. [Google Scholar] [CrossRef]
- Koh, C.; Canini, L.; Dahari, H.; Zhao, X.; Uprichard, S.L.; Haynes-Williams, V.; Winters, M.A.; Subramanya, G.; Cooper, S.L.; Pinto, P.; et al. Oral Prenylation Inhibition with Lonafarnib in Chronic Hepatitis D Infection: A Proof-of-Concept Randomised, Double-Blind, Placebo-Controlled Phase 2A Trial. Lancet Infect. Dis. 2015, 15, 1167–1174. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witwit, H.; Betancourt, C.A.; Cubitt, B.; Khafaji, R.; Kowalski, H.; Jackson, N.; Ye, C.; Martinez-Sobrido, L.; de la Torre, J.C. Cellular N-Myristoyl Transferases Are Required for Mammarenavirus Multiplication. Viruses 2024, 16, 1362. https://doi.org/10.3390/v16091362
Witwit H, Betancourt CA, Cubitt B, Khafaji R, Kowalski H, Jackson N, Ye C, Martinez-Sobrido L, de la Torre JC. Cellular N-Myristoyl Transferases Are Required for Mammarenavirus Multiplication. Viruses. 2024; 16(9):1362. https://doi.org/10.3390/v16091362
Chicago/Turabian StyleWitwit, Haydar, Carlos Alberto Betancourt, Beatrice Cubitt, Roaa Khafaji, Heinrich Kowalski, Nathaniel Jackson, Chengjin Ye, Luis Martinez-Sobrido, and Juan C. de la Torre. 2024. "Cellular N-Myristoyl Transferases Are Required for Mammarenavirus Multiplication" Viruses 16, no. 9: 1362. https://doi.org/10.3390/v16091362