Abstract
Hepatitis B virus (HBV) infection remains a major public health concern worldwide, with approximately 296 million individuals chronically infected. The HBV-encoded X protein (HBx) is a regulatory protein of 17 kDa, reportedly responsible for a broad range of functions, including viral replication and oncogenic processes. In this review, we summarize the state of knowledge on the mechanisms underlying HBx functions in viral replication, the antiviral effect of therapeutics directed against HBx, and the role of HBx in liver cancer development (including a hypothetical model of hepatocarcinogenesis). We conclude by highlighting major unanswered questions in the field and the implications of their answers.
1. Introduction
Infection with the hepatitis B virus (HBV) is one of the most common bloodborne diseases, having infected approximately one third of the world’s population and chronically affecting 296 million people [1]. Viral persistence and the chronic host immune responses against infected cells drive ongoing liver injury, which can result in cirrhosis and liver cancer—disease states that kill ~1 million people per year [2,3]. One of the most long-standing mysteries surrounding HBV is the function of the hepatitis B X protein (HBx), a small trans-activator that has a disordered structure and multiple reported roles in viral replication and cancer development [4]. Here, we review the current state of the art on the roles of functions of this enigmatic protein and potential therapeutics to inhibit it.
3. Hepadnavirus Life Cycle
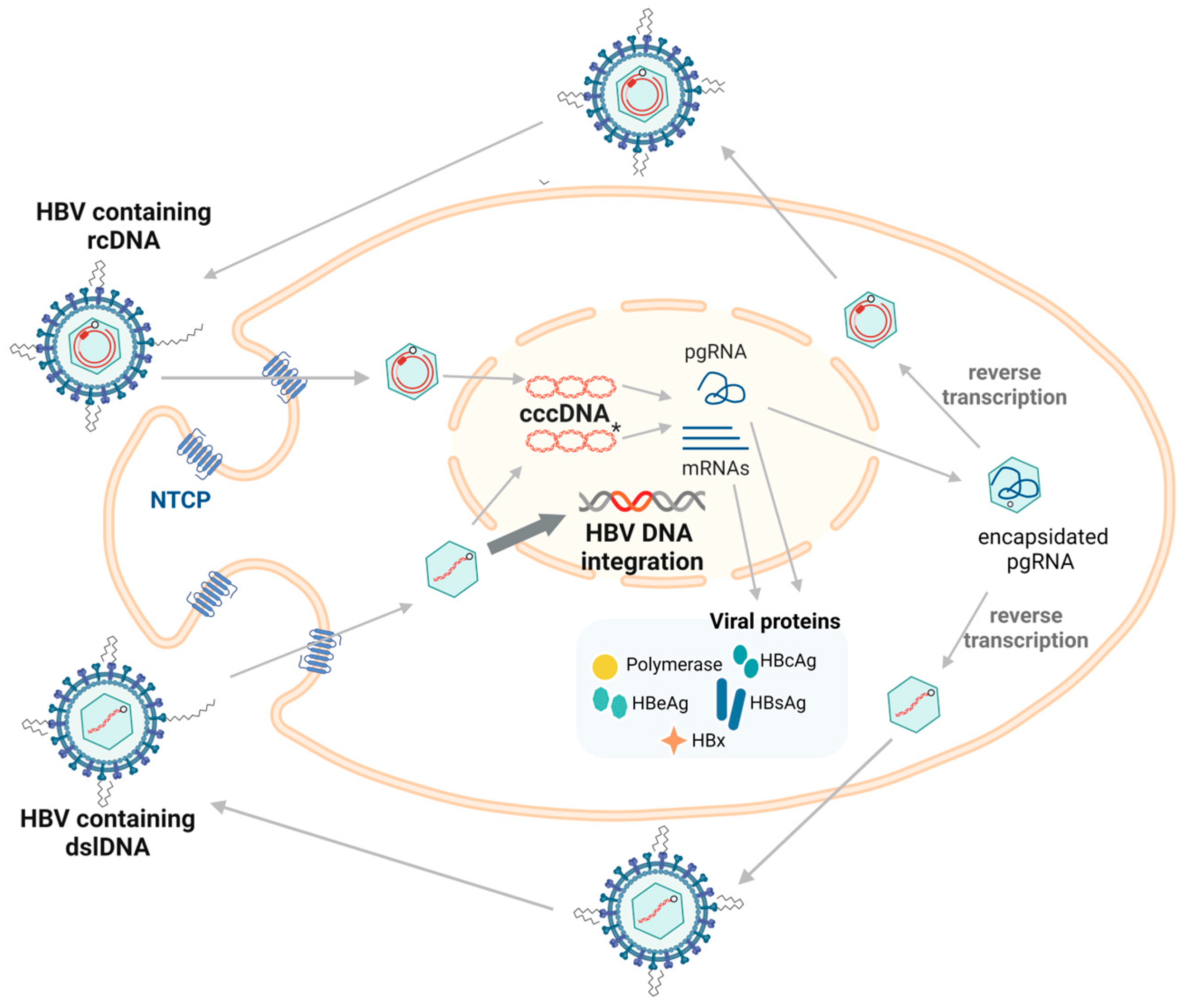
Hepadnaviruses are liver-tropic and mainly infect the hepatocytes (Figure 1). Virions first attach to cell surfaces with low affinity via binding of heparan sulfate proteoglycans. Subsequently, the large envelope protein binds with high affinity to hepatocyte specific receptors (NTCP in the case of human HBV [15], still unknown in many other hepadnaviruses). Following endocytosis and removal of the viral envelope and nucleocapsid, the viral capsid containing the relaxed circular DNA (rcDNA) is transported to the nucleus [16,17], where the genome undergoes repair by host DNA repair enzymes and forms covalently closed circular DNA (cccDNA) [18]. Using cccDNA as the template, viral RNAs are transcribed (among which the HBx transcript is amongst the first in infections by orthohepadnaviruses [19]) and exported into the cytoplasm where they are translated to form the viral proteins. The replicative pre-genomic RNA (pgRNA) is packaged by core protein, forming the viral nucleocapsid [11,12]. The viral DNA-containing nucleocapsids are enveloped at the endoplasmic reticulum and secreted as virions [13]. In avihepadnaviruses, nucleocapsids can also be recycled to the nucleus and contribute to the cccDNA pool of an infected cell [5,8]. In orthohepadnaviruses, this recycling pathway does not appear to be as active, leading to lower numbers of cccDNA molecules per cell (1–10 cccDNA copies per cell, compared to >50 copies in avihepadnavirus infection) [20,21].
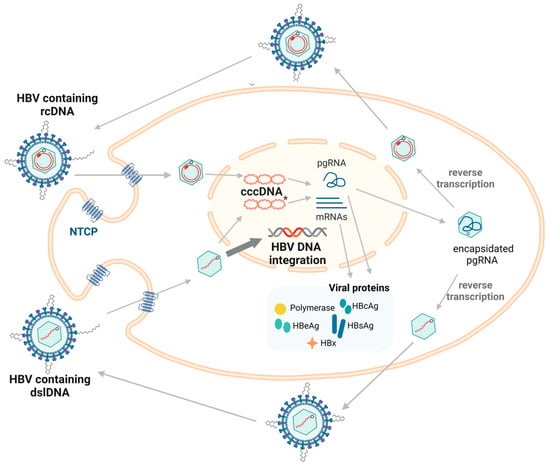
Figure 1.
The replication cycle of HBV. The HBV enters the hepatocyte via endocytosis using the cell surface receptor (NTCP). The capsid containing relaxed circular DNA (rcDNA) is transported to the nucleus where it undergoes repair to form cccDNA, which is the main transcriptional template for both pgRNA and mRNAs that are translated into viral proteins. HBcAg forms the nucleocapsid surrounding pgRNA and initiates reverse transcription to re-form viral DNA. Most of the time, rcDNA is formed, with double-stranded linear DNA (dslDNA) alternatively produced as a by-product (5–10% of synthesized virions) [22]. The mature nucleocapsids containing viral DNA are enveloped and secreted as virions. dslDNA can integrate into the host genome at double stranded DNA breaks (highlighted using bold arrow). dslDNA can also form cccDNA but cannot code for functional rcDNA due to an additional 16nt insertion [23]. Figure adapted from [23] and generated in BioRender (https://biorender.com/, accessed on 20 August 2024).
Besides this replicative cycle, a minority of nucleocapsids contain double stranded linear DNA (dslDNA) rather than rcDNA. These are formed as a by-product during reverse transcription of the pgRNA, which can be released as defective virions, infect surrounding cells, and integrate into the host genome at the site of cellular double-stranded DNA breaks [24,25,26,27]. Integration events are relatively rare, occurring in ~1 per 10,000 infected cells [26,28,29]. Despite their general rarity, integrated forms are strongly associated with liver cancer: ~90% of HBV-associated liver cancers contain HBV DNA integrations [30,31,32,33,34].
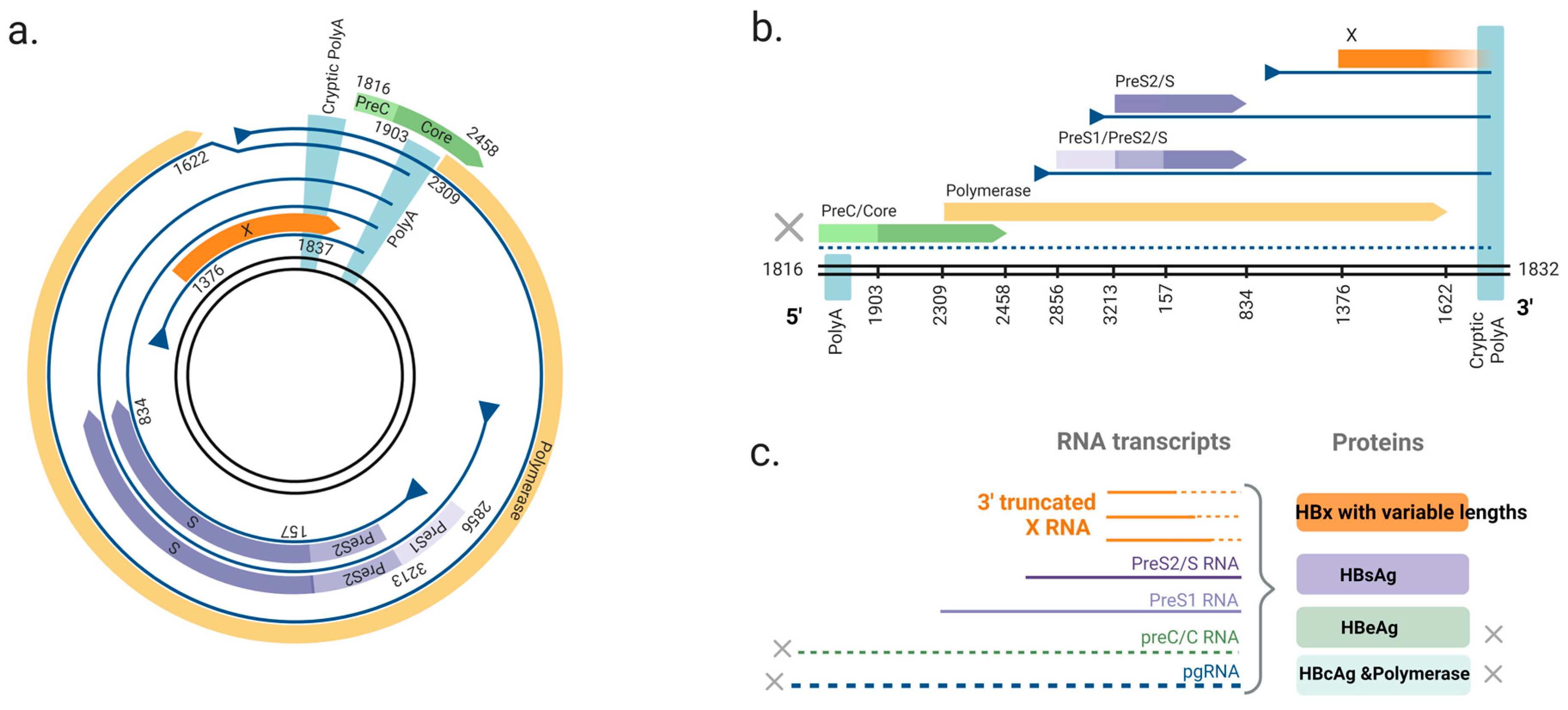
Integrations are amplified when cells with integrations undergo mitosis [30,31,32,35]. This allows HBV DNA integrations to persist and become the major source of HBV antigens in later phases of chronic infection [33,36,37]. Although HBV DNA integration normally fails to transcribe pre-core mRNA and pgRNA due to the loss of the upstream basal core promoter, the promoters of S and X ORF are intact, allowing the synthesis of functional HBsAg and C-terminal truncated HBx proteins [36,38,39] (Figure 2).
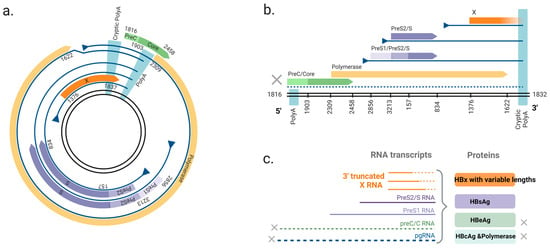
Figure 2.
The ORFs of two forms of HBV DNA and derived viral RNAs. (a) The circular HBV DNA. The HBV genome consists of four overlapping ORFs: polymerase, pre-core/core, surface and X. The viral mRNAs expressed from circular DNA terminate at a polyadenylation signal (polyA) located in the core ORF. (b) dslDNA form HBV DNA. The canonical polyA is located at the 5′ end and mRNAs instead terminate at a non-canonical cryptic polyA. The structural arrangement of the viral genome leads to the separation of pre-c/c and polymerase ORFs from the native promoter (core promoter), leading to loss of mRNA expression (dashed line). The X ORF is truncated at its C-terminus. (c) Viral transcripts from the dslDNA and the viral proteins. Pre-c/c RNA (encoding HBeAg) and pgRNA (encoding HBcAg and polymerase) are not transcribed due to the missing 5′ promoter. 3′ end truncated X RNA transcripts are produced from the dslDNA and transcribed to HBx protein with variable lengths. HBV DNA genome: the double black lines; ORFs: colored arrows (green = pre-c/c, yellow = polymerase, purple = surface, orange = x); transcripts: blue lines; promoters: blue triangles. Nucleotide numbering of ORFs is shown as per GenBank Accession #AB241115. Figure adapted from [23] and generated in BioRender (https://biorender.com/, accessed on 20 August 2024).
4. HBx
A key difference between avihepadnaviruses and orthohepadnaviruses is the presence of HBx ORF (only in the latter), which encodes the HBx protein and appears to be a function acquired by orthohepadnaviruses to adapt to mammalian hosts during evolution [40].
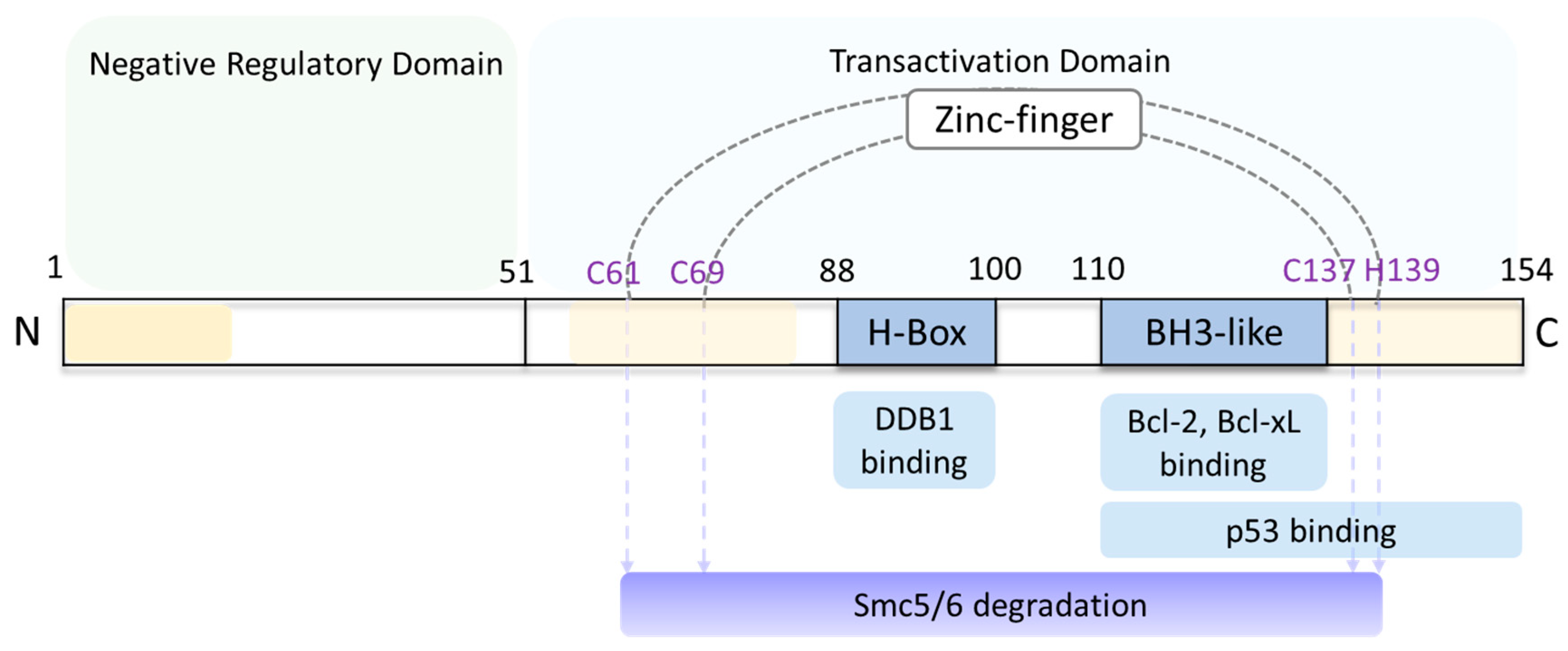
HBx is a small 154 amino acid (aa) 17 kDa protein. Three conserved regions (aa1–20, aa58–84 and aa120–154) have been identified based on the sequence homologies among eight HBV genotypes [41,42]. The N-terminus functions as a negative regulatory domain (aa1–50), while the C-terminus functions as a transactivation domain (aa51–154) [43,44,45] (Figure 3). The N-terminal domain is predicted to be largely unstructured, suggesting high flexibility for binding with diverse interactors [46,47]. The C-terminal domain is relatively well conserved due to the overlap with several regulatory elements [43,45]. The unstructured domain and the poor solubilization of HBx has led to technical difficulties in elucidating the crystal structure of HBx. While small domains of HBx have been crystalized and have solved structures [48,49,50,51,52], the structure of a fully functional HBx remains unknown.
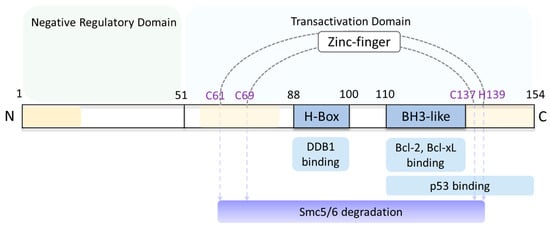
Figure 3.
Scheme of HBx protein functional domains. The 154-amino acid (aa) HBx has a negative regulatory domain located at the N-terminus (aa1–50) that is predicted to be predominantly unstructured, and a transactivation domain located at the C-terminus (aa51–154). H-box motif (aa88–100) binds DDB1 and CCCH (C61, C69, C137 and H139) zinc finger motifs, both of which target the Smc5/6 complex for degradation [41,49]. BH3-like motif (aa110–135) binds with Bcl protein family, promoting HBV replication and cytotoxicity [48,50]. p53 binding motif (aa111–154) represses p53-mediated apoptosis [52]. Three conserved regions (aa1–20, aa58–84 and aa120–154) are colored yellow. The figure is adapted from [53].
HBx is predicted to form homodimers via disulfide bonds and acetylation [54]. Its conformational flexibility may let it develop different secondary structures under specific conditions, such as become folded upon binding to its target molecules (with a wide array of signaling proteins, transcriptional regulators and nucleic acids) [47]. Intracellular localization studies based on HBV patient-derived liver biopsies showed that HBx predominantly accumulates in the cytoplasm when highly expressed, whereas low expression leads to localization primarily in the nucleus [55,56,57]. It has been demonstrated that the localization of HBx to the nucleus is essential for its role in virus replication in vitro and in vivo [13]. Recent discoveries showed that HBx can coordinate a zinc ion (Zn) or an iron-sulfur (Fe–S) cluster through metal-binding cysteine residues located at the C-terminus. This highlights the potential for HBx involvement in metalloprotein-associated functions such as redox regulation, electron transfer, DNA binding and repair mechanisms. This coordination could be important for structural stability, protein–protein interactions, or catalysis, which are key to its various roles in viral replication and the modulation of host cellular processes [49,50,58].
5. Role of HBx in HBV Replication
HBx appears to be essential for viral transcription [19,59]. Chen et al. and Zoulim et al. reported that the orthohepandavirus woodchuck hepatitis virus (WHV) bearing mutations in the X gene was unable to infect or initiate infection in susceptible woodchucks, suggesting that the WHV X gene is important for viral replication [59,60]. Indeed, this has also been shown for human HBV in in vitro infection model, as well as in hydrodynamic mouse models [19,61]. Recent studies have begun to clarify the specific roles of HBx in viral replication (Table 1).

Table 1.
Roles of HBx in Viral Replication.
5.1. HBx and DDB1
HBx appears to overcome cellular restriction factors that inhibit viral transcription. Specific regions of HBx that restore viral transcription have been identified, including a conserved alpha-helical motif (H-box, aa88–100) [51,62,63,79] and a zinc-finger containing transactivating regions (CCCH motif) [41,64] (Figure 3).
In a landmark paper, Decorsiere et al. showed that HBx leads to the degradation of the structural maintenance of the chromosome 5/6 (Smc5/6) complex, which was shown to inhibit the expression of genes from episomal DNA [65]. The Smc5/6 complex bridges DNA molecules through topological entrapment and creates a repressive chromatin structure, therefore silencing viral gene expression [66,80]. Thus, the Smc5/6 complex functions as an HBV restriction factor that blocks viral transcription [65,66].
HBx overcomes this inhibition by binding and recruiting DNA damage-binding protein 1 (DDB1) to the Smc5/6 complex. DDB1 binds Cullin4 (Cul4) as part of an E3 ubiquitin ligase complex that recruits substrate proteins for ubiquitination and degradation [46,51]. By recruiting the DDB1-E3 ligase complex to the Smc5/6 complex, HBx triggers its degradation and therefore allows the transcriptional restriction of cccDNA to be lifted [64,65,66,67]. Indeed, the region of HBx involved in DDB1 binding is the highly conserved H-box domain, suggesting its strong selective advantage and likely role in viral replication of all orthohepadnaviruses.
Other studies support this function of HBx in viral replication. For example, genetic knockdown of Smc5/6 restores the replication of HBx-deficient HBV [66]. A study using clinical samples has shown that the anti-Smc5/6 function can be retained in HBx variants found in patients with hepatocellular carcinoma (HCC) [68]. Cells with either DDB1-binding deficient HBx or Cul4-binding deficient DDB1 exhibited no cccDNA transcription [67]. In the absence of HBx, Smc5/6 anchors cccDNA to nuclear domain 10 (ND10) bodies, nuclear regions of transcriptional repression. This is in contrast to cccDNA in the presence of HBx, which localizes to regions of active transcription [46,67]. Finally, despite the marked diversity of HBx among orthohepadnaviruses (HBx has a percentage identity ranging from 87% compared to orangutan HBV to 30% in Asian grey shrew HBV) [81,82], inhibiting the action of Smc5/6 is a conserved feature in all known members [49,83]. Together, these findings have resulted in a robust model of how HBx controls viral transcription in orthohepadnaviruses.
5.2. HBx and Histone Modification
Other mechanisms by which HBx has been reported to drive HBV transcription include modifying histones. Efficient transcription from cccDNA requires histone binding and transcriptional regulators (e.g., transcription activity is enhanced when the histones are acetylated) [84,85,86]. HBx has been shown to increase HBV transcription through recruiting of histone acetyltransferases such as CREB-binding protein (CBP)/p300 to cccDNA, promoting histone acetylation to maintain active transcription [69]. In the absence of HBx, the viral genome exists in a repressed chromatin state marked by hypoacetylation and histone H3K9 methylation, correlating with the recruitment of histone deacetylases HDAC1 and H3K9 methyltransferase SET domain bifurcated 1 (SETDB1), as well as the recruitment of the heterochromatin protein HP1, and the cccDNA transcribes significantly fewer RNAs [19,69,70,73].
HBx has also been reported to modify the epigenetic landscape of cccDNA and host chromatin through interactions with DNA methyltransferases (DNMTs) to promote transcription [70,87]. HBx binds protein arginine methyltransferase 1 (PRMT1) and the Tudor-domain protein Spindlin-1, blocking the inhibitory activity on HBV transcription [53,71,72]. HBx also can recruit lysine-specific demethylase 1 (LSD1) and lysine methyltransferase Set1A to the viral promoter, which inhibit H3K9me2 demethylation and accumulate activated H3K4me3 to activate viral transcription [74].
5.3. HBx and Cell Signaling Pathways
Some studies have reported that HBx can also directly interact with proteins in the cytoplasm to induce cellular changes. HBx has been found to form a complex with proteins such as MEKK1, SEK1, and SAPK/JNK, resulting in prolonged cell survival and maintaining a cellular environment favorable to HBV replication, reviewed in Schollmeier et al. [54], Diao et al. [88], and Agustiningsih et al. [89]. HBx could increase cytosolic calcium levels through modulation of the mitochondrial permeability transition pore, thereby activating the Pyk2/Src and FAK pathways [75,76]. HBx-mediated Ca2+ signaling also facilitates viral core assembly and pgRNA production, both of which are required for virus replication [76,77]. However, cytosolic HBx localization has only been observed in overexpression models. Most of the interactions of HBx with cellular processes have been studied in many different models, often leading to significant overexpression of HBx and outside the context of natural HBV infection. Thus, it remains unclear whether similar manipulations of the cellular machinery by HBx would also occur in the context of an authentic HBV infection [90].
7. HBx in Hepadnavirus-Associated Liver Cancer
The other major difference between avihepadnaviruses and orthohepadnaviruses is the association with liver cancer (solely with infection with the latter). Hepadnavirus-associated liver cancer has been found in humans [97], woodchucks [98], cats [99], tree shrews [100] and ground squirrels [101], while it is rarely reported in avihepadnavirus-infected birds. Thus, the field has long suspected that HBx contributes to liver pathogenesis and cancer.
The molecular mechanisms regulated by the HBx protein (generally full-length HBx and largely overexpression studies) that drive pathogenesis was reviewed and summarized in a recent comprehensive review [102].
Rather than the cccDNA-encoded HBx being involved in viral replication, it is likely that integrated HBV DNA is the template for the HBx that drives liver cancer. As infected cells undergo mitosis (e.g., during clonal expansion of a cellular clone after acquisition of a driver mutation), they lose cccDNA, resulting in two uninfected daughter cells [103,104]. In contrast, integrated HBV DNA is replicated with the cellular genome and is passed down along the lineage of the clone. Indeed, quantification of specific integrated HBV DNA junctions has been used to measure the level of clonal expansion in infected liver tissues [30,31,32,33]. Moreover, during the dedifferentiation process leading up to liver cancer, hepatocytes lose their susceptibility to HBV infection (due to down-regulation of the receptor NTCP) [105], so no new cccDNA can be formed within the cells. Indeed, this assumption is consistent with the natural history of HBV-associated liver cancer, which occurs in later phases of HBV infection where cccDNA levels are relatively low in the liver (~1/100 to 1/1000 copies per cell) [106] and integrated HBV DNA is the dominant source of viral antigens. Indeed, integrated HBV is also the major form of viral DNA in liver cancers (1–5 copies of integrated HBV DNA vs. 0.00026–1 copies of cccDNA per cell) [107,108,109].
As mentioned above, the integration process leads to terminal deletions of the HBV DNA and subsequent C-terminal truncation of HBx, leading into the cellular sequences. It is unclear whether the HBx derived from integrated HBV DNA may be more pathogenic, but several studies have shown altered phenotypes driven by HBx with C-terminal truncations. C-terminal truncations of HBx decrease HBx steady-state levels, but also impair HBx’s activation of nuclear factor-κB (NF-κB, a master regulator of the cellular stress response) [110]. C-terminal truncations of HBx also abrogate the anti-proliferative and transactivating functions of HBx [111].
On the transcript level, 3′ truncations result in the canonical poly A signal to be cleaved from the HBx RNA. This may result in several types of HBx mRNAs: (1) mRNAs lacking a poly A tail are less stable and more prone to degradation [112]; (2) mRNAs use a poly A signal from the cellular sequence instead, resulting in a chimeric virus–host mRNA; or (3) mRNAs use a cryptic poly A signal in the HBV sequence instead (non-canonical sequence UAUAAA) [113].
Type 1 mRNAs (unstable poly A tail-deficient) are likely to lead to reduced levels of HBx from integrants and therefore mRNAs of this type are unlikely to alter the phenotype of the cell. Type 2 mRNAs (chimeric transcripts) are also unlikely to be a consistent driving factor for liver cancer as integrations are largely randomly distributed across the genome rather than targeting specific oncogenes or tumor suppressor genes. This reduces the likelihood of disrupting or activating genes directly that might promote carcinogenesis. Type 3 mRNAs (cryptic poly A signal) have been investigated as drivers of cancer and their proposed roles will be summarized in the rest of this section (Table 2).
Table 2.
A Summary of C-terminal Truncated HBx in Carcinogenesis.
7.1. HBx and Smc5/6 Complex Disruption
Truncated HBx proteins encoded by Type 3 mRNAs would retain an intact H-box motif and therefore are likely to still bind to DDB1 [64,96], thereby promoting the degradation of Smc5/6 complexes. In addition to silencing transcription from episomal DNA, Smc5/6 complexes are crucial for chromosomal stability [65,66] through several mechanisms: (1) coordination of DNA repair through post-translational modification of host repair proteins to establish DNA-damage-dependent cohesion [125]; (2) stabilization of stalled DNA replication forks, preventing their collapse and allowing proper resumption of DNA synthesis [126,127]; and (3) maintenance of telomeres, the protective ends of chromosomes [128].
Indeed, HBx has been shown to drive DNA damage and genomic instability through Smc5/6 complex degradation in both HBV in vitro and in vivo models [125,129]. HBx has been shown to cause impaired homologous recombination (HR) repair of DNA double-strand breaks [129]. Consequently, DNA damage accumulates within the cell, leading to genomic instability [66,78]. Studies have reported that DNA damage accumulates in the liver tissue of HBV-infected humanized chimeric mice, HBx-transgenic mice and human tissues [129,130]. HBx suppressed the HR repair of DNA double-strand breaks, including that induced by a CRISPR-Cas9 system, which was rescued by restoring the Smc5/6 complex [129]. Smc5/6 degradation may also affect cells to genetic errors under conditions of DNA damage (induced by necroinflammation in chronic hepatitis B), and reduced expression of the NSMCE2 subunit, which is associated with increased cancer incidence in mice [131].
HBx-DBB1 also disrupts the function of transcription factor IIH (TFIIH), an essential component of the DNA repair pathway, making cells more sensitive to ultraviolet light and decreasing their ability to repair their DNA, potentially promoting carcinogenesis [44]. HBx-DDB1 may cause alterations in the function of transcription factor p53, which maintains genomic stability by regulating the cell cycle and promoting DNA repair in response to DNA damage [124]. This may induce cell apoptosis and inhibits nucleotide excision repair, thereby compromising genome integrity [124,132].
7.2. HBx and Cancer-Related Signaling Pathways
The intracellular concentration of HBx likely increases with additional integration events following each bout of hepatitis and regeneration, resulting in outgrowth of HBx-positive hepatocytes. The sustained high levels of HBx block tumor necrosis factor-α (TNFα) and fatty acid synthase (FAS)-mediated apoptosis by activation of NF κB, thus infected hepatocytes survive immune-mediated damage and resistance to apoptosis [133].
Altered cellular signaling pathways play a critical role in the progression of HBV-associated HCC. C-terminal truncated X protein expression can promote hepatocyte proliferation and reprogram cell metabolism by inhibiting thioredoxin-interacting protein (TXNIP) [114]. The accumulated truncated HBx can prevent apoptosis and lead to the progress of stem cell-like characteristics such as self-renewal, tumorigenicity, chemoresistance, and migration, promoting the neoplastic transformation of the hepatocytes [111,115,116,117,118].
Furthermore, C-terminal truncated HBx regulates the transcription of Caveolin-1 and stabilizes LRP6 to maintain the activation of β-catenin, promote the progression of HBV-associated HCC [119], and enhance the invasion and metastasis of HCC cells [116,120]. In addition, integrated viral DNA can lead to the persistent expression of mutated and truncated proteins, which is associated with endoplasmic reticulum and mitochondrial stress responses and may increase the risk of HCC [121,134].
C-terminal truncated HBx is associated with increased reactive oxygen species production and mitochondrial DNA damage, marked by 8-oxoguanine formation, suggesting its role in oxidative stress-induced tumor metastasis [122]. Furthermore, in farnesoid X receptor (FXR)-deficient HCC, C-terminal truncated HBx exacerbates oncogenesis by disrupting cell cycle regulation and glucose metabolism [123]. These findings underscore the importance of targeting metabolic disruptions for therapeutic interventions in HBV-associated HCC.
8. A Hypothetical Model of HBx-Mediated Carcinogenesis
While many studies have shown that overexpression of HBx can drive multiple oncogenic pathways, it is important to consider the context within which liver cancer develops and the template from which HBx is expressed during this development.
Given the asymptomatic nature of liver cancer (and therefore its lack of an impact on spreading the virus infection), the induction of HCC is not likely a feature that is selected for in HBV strains. That is to say, liver cancer induction is likely an “unintended” consequence of the pro-viral functions of HBx.
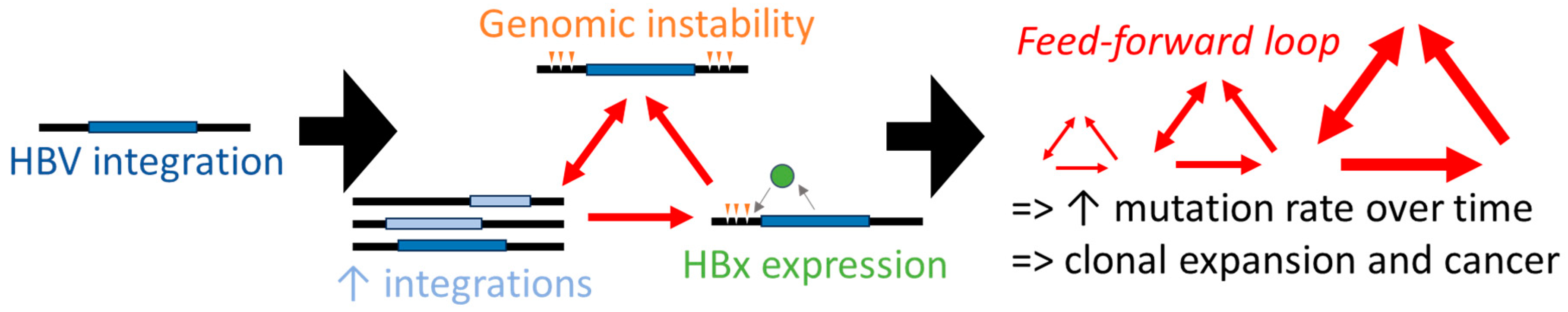
Using the discoveries described in this review, we have developed a hypothetical model with feed-forward loops (Figure 4). We propose that virus integrations drive genomic instability, which in turn may lead to more integrations through the promotion of more double-stranded DNA breaks in the host chromosome. This promotes a self-amplifying process and the resultant exponential increasing genomic instability may fuel the accelerated acquisition of cancer driver mutations.
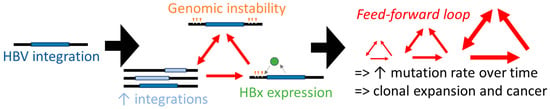
Figure 4.
A hypothetical model where cancer development is accelerated by integrated HBV DNA. Genomic instability is driven by virus integrations, which in turn lead to more integrations through the promotion of more double-stranded DNA breaks in the host chromosome (cells with HBV integrations can express functional HBx and may be more susceptible to additional integrations), promoting a self-amplifying process (red arrows). The resultant exponential increasing genomic instability then fuels the accelerated acquisition of cancer driver mutations.
9. Unanswered Questions in HBx Research
Despite recent progress in understanding HBx and its role in both viral replication and carcinogenesis, many unknowns remain in the field.
9.1. What Is the Structure of HBx in Various Cellular Contexts?
Structure determines the function of a protein. Given the high flexibility of its N-terminus, it is likely that HBx takes various forms with different interacting partners. Novel advances in experimental structural analysis (e.g., cryo-electron microscopy) [135] and in silico modeling (e.g., Alphafold 3) [136] may elucidate this. Solving the structures of HBx with key interactors could open up the field of HBx-targeting antiviral therapeutics.
That it is a metalloprotein capable of coordinating a Zn or an Fe–S cluster adds a new dimension to our understanding of its molecular functions [49,50,58]. Fe–S clusters are essential for a range of cellular activities, including enzymatic reactions, regulatory functions, and protection against oxidative damage. For viruses, which rely on the host machinery for replication, the incorporation of such metallocofactors may be vital for altering host cellular pathways, possibly assisting in viral replication, modulation of immune responses, or influencing cellular redox states. Future studies exploring the structural biology of HBx, including how it coordinates these metal ions and the impact on its interactions with host proteins, could lead to novel therapeutic targets, particularly in the context of inhibiting these metal-binding activities to prevent HBV-related liver diseases.
9.2. Is Transcriptional Regulation of Integrated HBV DNA Genomes Independent of HBx?
Given that Smc5/6 theoretically only regulates episomal DNA, it is possible that transcription of integrated HBV DNA is not under the restriction activity of Smc5/6. Thus, HBx from integrated HBV DNA may be a mechanism by which transcriptionally silenced cccDNA can be rescued to reactivate an inactive infection.
Indeed, given the conservation of DDB1 interacting domains, truncated HBx encoded by integrated HBV DNA likely retains the function of maintaining active transcription of cccDNA [137]. If this is shown, then it may suggest that HBx inhibitors may be an additional mechanism by which to prevent HBV reactivation.
9.3. What Are the Most Appropriate Models to Test HBx Function and Anti-HBx Therapies?
Over-expression of HBx can cause aberrant localization and epiphenomena in vitro. Several models have been utilized to test HBx function and anti-HBx therapies, with specific advantages and limitations. The in vitro NTCP-dependent HBV infection system is widely accepted as a robust model for studying the role of HBx in viral replication within a physiologically relevant setting (such as HepG2-NTCP, HepaRG, other engineered cells, or primary human hepatocytes) [90]. However, it is still unclear if there is an optimal model in which to study the roles of HBx in genomic instability and carcinogenic processes.
9.4. Can Anti-HBx Therapies Prevent or Reduce HBV-Associated Liver Cancer?
By transcriptionally silencing (e.g., using siRNA or epigenetic silencing), mutating (e.g., using CRISPR), or inhibiting the function (e.g., via small molecule inhibitors) of HBx, studies have shown effects on HBx-dependent replication and to some extent DNA damage. However, it is still unclear whether these approaches could induce a reduction of HBV-associated liver cancer, independent of their antiviral effects. Indeed, there is not yet an optimal preclinical model to test this question.
10. Conclusions
HBx has various biological functions and may need this flexibility to mediate its function(s) in the different cellular environments encountered during a decades-long chronic HBV infection. Given the clinical importance of chronic HBV infection, the central role of HBx in HBV replication and in the occurrence and progression of HBV-associated HCC, antiviral strategies targeting HBx may be promising clinical treatment strategies.
Author Contributions
Conceptualization, T.T.; writing—initial draft, Y.H.; writing—full draft preparation, D.L.; writing—review and editing, T.T. and D.L.; figure generation, D.L.; supervision, T.T.; project funding acquisition, T.T. All authors have read and agreed to the published version of the manuscript.
Funding
T.T. was supported by the Paul and Valeria Ainsworth Precision Medicine Fellowship. The APC was funded by the Westmead Institute for Medical Research.
Conflicts of Interest
The authors declare no relevant conflicts of interest.
References
- Hsu, Y.-C.; Huang, D.Q.; Nguyen, M.H. Global burden of hepatitis B virus: Current status, missed opportunities and a call for action. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Global Hepatitis Report 2024: Action for Access in Low- and Middle-Income Countries; States News Service: Washington, DC, USA, 2024.
- Sherman, M. Hepatocellular Carcinoma: Epidemiology, Surveillance, and Diagnosis. Semin. Liver Dis. 2010, 30, 003–016. [Google Scholar] [CrossRef]
- Jin, Y.M.; Yun, C.; Park, C.; Wang, H.; Cho, H. Expression of hepatitis B virus X protein is closely correlated with the high periportal inflammatory activity of liver diseases. J. Viral Hepat. 2001, 8, 322–330. [Google Scholar] [CrossRef]
- Magnius, L.; Mason, W.S.; Taylor, J.; Kann, M.; Glebe, D.; Dény, P.; Sureau, C.; Norder, H.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Hepadnaviridae. J. Gen. Virol. 2020, 101, 571–572. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.J. Hepatitis B: The virus and disease. Hepatology 2009, 49 (Suppl. S5), S13–S21. [Google Scholar] [CrossRef]
- Gavilanes, F.; Gonzalez-Ros, J.M.; Peterson, D.L. Structure of hepatitis B surface antigen. Characterization of the lipid components and their association with the viral proteins. J. Biol. Chem. 1982, 257, 7770–7777. [Google Scholar] [CrossRef]
- Hu, J.; Seeger, C. Hepadnavirus Genome Replication and Persistence. Cold Spring Harb. Perspect. Med. 2015, 5, a021386. [Google Scholar] [CrossRef] [PubMed]
- Gerlich, W.H.; Robinson, W.S. Hepatitis B virus contains protein attached to the 5′ terminus of its complete DNA strand. Cell 1980, 21, 801–809. [Google Scholar] [CrossRef]
- Nassal, M. Hepatitis B viruses: Reverse transcription a different way. Virus Res. 2008, 134, 235–249. [Google Scholar] [CrossRef]
- Bartenschlager, R.; Schaller, H. Hepadnaviral assembly is initiated by polymerase binding to the encapsidation signal in the viral RNA genome. EMBO J. 1992, 11, 3413–3420. [Google Scholar] [CrossRef]
- Hirsch, R.C.; Loeb, D.D.; Pollack, J.R.; Ganem, D. cis-acting sequences required for encapsidation of duck hepatitis B virus pregenomic RNA. J. Virol. 1991, 65, 3309–3316. [Google Scholar] [CrossRef]
- Selzer, L.; Zlotnick, A. Assembly and Release of Hepatitis B Virus. Cold Spring Harb. Perspect. Med. 2015, 5, a021394. [Google Scholar] [CrossRef]
- Chen, M.T.; Billaud, J.-N.; Sällberg, M.; Guidotti, L.G.; Chisari, F.V.; Jones, J.; Hughes, J.; Milich, D.R. A Function of the Hepatitis B Virus Precore Protein Is to Regulate the Immune Response to the Core Antigen. Proc. Natl. Acad. Sci. USA 2004, 101, 14913–14918. [Google Scholar] [CrossRef] [PubMed]
- Wettengel, J.M.; Burwitz, B.J. Innovative HBV Animal Models Based on the Entry Receptor NTCP. Viruses 2020, 12, 828. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne, R.J.; Bagga, S.; Bouchard, M.J. Hepatitis B virus molecular biology and pathogenesis. Hepatoma Res. 2016, 2, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-C.; Huang, E.-Y.; Su, P.-Y.; Wu, S.-Y.; Yang, C.-C.; Lin, Y.-S.; Chang, W.-C.; Shih, C. Nuclear Export and Import of Human Hepatitis B Virus Capsid Protein and Particles. PLoS Pathog. 2010, 6, e1001162. [Google Scholar] [CrossRef]
- Urban, S.; Schulze, A.; Dandri, M.; Petersen, J. The replication cycle of hepatitis B virus. J. Hepatol. 2010, 52, 282–284. [Google Scholar] [CrossRef]
- Lucifora, J.; Arzberger, S.; Durantel, D.; Belloni, L.; Strubin, M.; Levrero, M.; Zoulim, F.; Hantz, O.; Protzer, U. Hepatitis B virus X protein is essential to initiate and maintain virus replication after infection. J. Hepatol. 2011, 55, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Y.; Zhang, B.-H.; Theele, D.; Litwin, S.; Toll, E.; Summers, J. Single-Cell Analysis of Covalently Closed Circular DNA Copy Numbers in a Hepadnavirus-Infected Liver. Proc. Natl. Acad. Sci. USA 2003, 100, 12372–12377. [Google Scholar] [CrossRef]
- Moraleda, G.; Saputelli, J.; Aldrich, C.E.; Averett, D.; Condreay, L.; Mason, W.S. Lack of effect of antiviral therapy in nondividing hepatocyte cultures on the closed circular DNA of woodchuck hepatitis virus. J. Virol. 1997, 71, 9392–9399. [Google Scholar] [CrossRef]
- Tu, T.; Jilbert, A.R. Detection of Hepatocyte Clones Containing Integrated Hepatitis B Virus DNA Using Inverse Nested PCR. In Hepatitis B Virus; Humana Press: New York, NY, USA, 2017; pp. 97–118. [Google Scholar]
- Tu, T.; Zhang, H.; Urban, S. Hepatitis B Virus DNA Integration: In Vitro Models for Investigating Viral Pathogenesis and Persistence. Viruses 2021, 13, 180. [Google Scholar] [CrossRef]
- Tu, T.; Budzinska, M.A.; Shackel, N.A.; Urban, S. HBV DNA Integration: Molecular Mechanisms and Clinical Implications. Viruses 2017, 9, 75. [Google Scholar] [CrossRef]
- Salpini, R.; D’anna, S.; Benedetti, L.; Piermatteo, L.; Gill, U.; Svicher, V.; Kennedy, P.T.F. Hepatitis B virus DNA integration as a novel biomarker of hepatitis B virus-mediated pathogenetic properties and a barrier to the current strategies for hepatitis B virus cure. Front. Microbiol. 2022, 13, 972687. [Google Scholar] [CrossRef]
- Tu, T.; Budzinska, M.A.; Vondran, F.W.R.; Shackel, N.A.; Urban, S. Hepatitis B Virus DNA Integration Occurs Early in the Viral Life Cycle in an In Vitro Infection Model via Sodium Taurocholate Cotransporting Polypeptide-Dependent Uptake of Enveloped Virus Particles. J. Virol. 2018, 92, e02007-17. [Google Scholar] [CrossRef]
- Bill, C.A.; Summers, J. Genomic DNA Double-Strand Breaks Are Targets for Hepadnaviral DNA Integration. Proc. Natl. Acad. Sci. USA 2004, 101, 11135–11140. [Google Scholar] [CrossRef]
- Mason, W.S.; Low, H.-C.; Xu, C.; Aldrich, C.E.; Scougall, C.A.; Grosse, A.; Clouston, A.; Chavez, D.; Litwin, S.; Peri, S.; et al. Detection of Clonally Expanded Hepatocytes in Chimpanzees with Chronic Hepatitis B Virus Infection. J. Virol. 2009, 83, 8396–8408. [Google Scholar] [CrossRef]
- Summers, J.; Jilbert, A.R.; Yang, W.; Aldrich, C.E.; Saputelli, J.; Litwin, S.; Toll, E.; Mason, W.S. Hepatocyte Turnover during Resolution of a Transient Hepadnaviral Infection. Proc. Natl. Acad. Sci. USA 2003, 100, 11652–11659. [Google Scholar] [CrossRef]
- Mason, W.S.; Gill, U.S.; Litwin, S.; Zhou, Y.; Peri, S.; Pop, O.; Hong, M.L.; Naik, S.; Quaglia, A.; Bertoletti, A.; et al. HBV DNA Integration and Clonal Hepatocyte Expansion in Chronic Hepatitis B Patients Considered Immune Tolerant. Gastroenterology 2016, 151, 986–998.e4. [Google Scholar] [CrossRef]
- Tu, T.; Mason, W.S.; Clouston, A.D.; Shackel, N.A.; McCaughan, G.W.; Yeh, M.M.; Schiff, E.R.; Ruszkiewicz, A.R.; Chen, J.W.; Harley, H.A.J.; et al. Clonal expansion of hepatocytes with a selective advantage occurs during all stages of chronic hepatitis B virus infection. J. Viral Hepat. 2015, 22, 737–753. [Google Scholar] [CrossRef]
- Mason, W.S.; Jilbert, A.R.; Summers, J. Clonal Expansion of Hepatocytes during Chronic Woodchuck Hepatitis Virus Infection. Proc. Natl. Acad. Sci. USA 2005, 102, 1139–1144. [Google Scholar] [CrossRef]
- Mason, W.S.; Liu, C.; Aldrich, C.E.; Litwin, S.; Yeh, M.M. Clonal Expansion of Normal-Appearing Human Hepatocytes during Chronic Hepatitis B Virus Infection. J. Virol. 2010, 84, 8308–8315. [Google Scholar] [CrossRef]
- Sung, W.-K.; Zheng, H.; Li, S.; Chen, R.; Liu, X.; Li, Y.; Lee, N.P.; Lee, W.H.; Ariyaratne, P.N.; Tennakoon, C.; et al. Genome-wide survey of recurrent HBV integration in hepatocellular carcinoma. Nat. Genet. 2012, 44, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Summers, J. Integration of hepadnavirus DNA in infected liver: Evidence for a linear precursor. J. Virol. 1999, 73, 9710–9717. [Google Scholar] [CrossRef] [PubMed]
- Podlaha, O.; Wu, G.; Downie, B.; Ramamurthy, R.; Gaggar, A.; Subramanian, M.; Ye, Z.; Jiang, Z. Genomic modeling of hepatitis B virus integration frequency in the human genome. PLoS ONE 2019, 14, e0220376. [Google Scholar] [CrossRef] [PubMed]
- Wooddell, C.I.; Yuen, M.-F.; Chan, H.L.-Y.; Gish, R.G.; Locarnini, S.A.; Chavez, D.; Ferrari, C.; Given, B.D.; Hamilton, J.; Kanner, S.B.; et al. RNAi-based treatment of chronically infected patients and chimpanzees reveals that integrated hepatitis B virus DNA is a source of HBsAg. Sci. Transl. Med. 2017, 9, eaan0241. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhang, Y.; Gu, W.; Wang, Z.; Li, D.; Zhang, F.; Qiu, G.; Xie, K. Integration of the hepatitis B virus X fragment in hepatocellular carcinoma and its effects on the expression of multiple molecules: A key to the cell cycle and apoptosis. Int. J. Oncol. 2005, 26, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Shaul, Y. Enhancer I Predominance in Hepatitis B Virus Gene Expression. Mol. Cell. Biol. 2004, 24, 1799–1808. [Google Scholar] [CrossRef]
- Lauber, C.; Seitz, S.; Mattei, S.; Suh, A.; Beck, J.; Herstein, J.; Börold, J.; Salzburger, W.; Kaderali, L.; Briggs, J.A.; et al. Deciphering the Origin and Evolution of Hepatitis B Viruses by Means of a Family of Non-enveloped Fish Viruses. Cell Host Microbe 2017, 22, 387–399.e6. [Google Scholar] [CrossRef]
- Kumar, V.; Jayasuryan, N.; Kumar, R. A Truncated Mutant (Residues 58-140) of the Hepatitis B Virus X Protein Retains Transactivation Function. Proc. Natl. Acad. Sci. USA 1996, 93, 5647–5652. [Google Scholar] [CrossRef]
- Kidd-Ljunggren, K.; Oberg, M.; Kidd, A.H. The hepatitis B virus X gene: Analysis of functional domain variation and gene phylogeny using multiple sequences. J. Gen. Virol. 1995, 76, 2119–2130. [Google Scholar] [CrossRef]
- Tang, H.; Oishi, N.; Kaneko, S.; Murakami, S. Molecular functions and biological roles of hepatitis B virus x protein. Cancer Sci. 2006, 97, 977–983. [Google Scholar] [CrossRef]
- Bouchard, M.J.; Schneider, R.J. The enigmatic X gene of hepatitis B virus. J. Virol. 2004, 78, 12725–12734. [Google Scholar] [CrossRef] [PubMed]
- Schuster, R.; Hildt, E.; Chang, S.-F.; Terradillos, O.; Pollicino, T.; Lanford, R.; Gerlich, W.H.; Will, H.; Schaefer, S. Conserved transactivating and pro-apoptotic functions of hepadnaviral X protein in ortho- and avihepadnaviruses. Oncogene 2002, 21, 6606–6613. [Google Scholar] [CrossRef]
- Prescott, N.A.; Bram, Y.; Schwartz, R.E.; David, Y. Targeting Hepatitis B Virus Covalently Closed Circular DNA and Hepatitis B Virus X Protein: Recent Advances and New Approaches. ACS Infect. Dis. 2019, 5, 1657–1667. [Google Scholar] [CrossRef]
- Rui, E.; de Moura, P.R.; Gonçalves, K.d.A.; Kobarg, J. Expression and spectroscopic analysis of a mutant hepatitis B virus onco-protein HBx without cysteine residues. J. Virol. Methods 2005, 126, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.-Y.; Chen, H.-Y.; Cao, J.-L.; Xiong, H.-L.; Mo, X.-B.; Li, T.-L.; Kang, X.-Z.; Zhao, J.-H.; Yin, B.; Zhao, X.; et al. Structural and functional analyses of hepatitis B virus X protein BH3-like domain and Bcl-xL interaction. Nat. Commun. 2019, 10, 3192. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, D.; Xing, W.; Beran, R.K.; Chemuru, S.; Rohrs, H.; Niedziela-Majka, A.; Marchand, B.; Mehra, U.; Zábranský, A.; Doležal, M.; et al. Hepatitis B Virus X Protein Function Requires Zinc Binding. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, M.; Wu, J.; Shi, Y. Structural and biochemical analysis of Bcl-2 interaction with the hepatitis B virus protein HBx. Proc. Natl. Acad. Sci. USA 2016, 113, 2074–2079. [Google Scholar] [CrossRef]
- Li, T.; Robert, E.I.; van Breugel, P.C.; Strubin, M.; Zheng, N. A promiscuous α-helical motif anchors viral hijackers and substrate receptors to the CUL4-DDB1 ubiquitin ligase machinery. Nat. Struct. Mol. Biol. 2010, 17, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Elmore, L.W.; Hancock, A.R.; Chang, S.-F.; Wang, X.W.; Chang, S.; Callahan, C.P.; Geller, D.A.; Will, H.; Harris, C.C. Hepatitis B Virus X Protein and p53 Tumor Suppressor Interactions in the Modulation of Apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 14707–14712. [Google Scholar] [CrossRef]
- Liu, W.; Yao, Q.; Su, X.; Deng, Y.; Yang, M.; Peng, B.; Zhao, F.; Du, C.; Zhang, X.; Zhu, J.; et al. Molecular insights into Spindlin1-HBx interplay and its impact on HBV transcription from cccDNA minichromosome. Nat. Commun. 2023, 14, 4663. [Google Scholar] [CrossRef] [PubMed]
- Schollmeier, A.; Glitscher, M.; Hildt, E. Relevance of HBx for Hepatitis B Virus-Associated Pathogenesis. Int. J. Mol. Sci. 2023, 24, 4964. [Google Scholar] [CrossRef] [PubMed]
- Martin-Vilchez, S.; Lara-Pezzi, E.; Trapero-Marugán, M.; Moreno-Otero, R.; Sanz-Cameno, P. The molecular and pathophysiological implications of hepatitis B X antigen in chronic hepatitis B virus infection. Rev. Med. Virol. 2011, 21, 315–329. [Google Scholar] [CrossRef]
- Cha, M.-Y.; Ryu, D.-K.; Jung, H.-S.; Chang, H.-E.; Ryu, W.-S. Stimulation of hepatitis B virus genome replication by HBx is linked to both nuclear and cytoplasmic HBx expression. J. Gen. Virol. 2009, 90, 978–986. [Google Scholar] [CrossRef]
- Henkler, F.; Hoare, J.; Waseem, N.; Goldin, R.D.; McGarvey, M.J.; Koshy, R.; King, I.A. Intracellular localization of the hepatitis B virus HBx protein. J. Gen. Virol. 2001, 82, 871–882. [Google Scholar] [CrossRef]
- Ueda, C.; Langton, M.; Chen, J.; Pandelia, M.-E. The HBx protein from hepatitis B virus coordinates a redox-active Fe-S cluster. J. Biol. Chem. 2022, 298, 101698. [Google Scholar] [CrossRef]
- Zoulim, F.; Saputelli, J.; Seeger, C. Woodchuck hepatitis virus X protein is required for viral infection in vivo. J. Virol. 1994, 68, 2026–2030. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.S.; Kaneko, S.; Girones, R.; Anderson, R.W.; Hornbuckle, W.E.; Tennant, B.C.; Cote, P.J.; Gerin, J.L.; Purcell, R.H.; Miller, R.H. The woodchuck hepatitis virus X gene is important for establishment of virus infection in woodchucks. J. Virol. 1993, 67, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Keasler, V.V.; Hodgson, A.J.; Madden, C.R.; Slagle, B.L. Hepatitis B virus HBx protein localized to the nucleus restores HBx-deficient virus replication in HepG2 cells and in vivo in hydrodynamically-injected mice. Virology 2009, 390, 122–129. [Google Scholar] [CrossRef]
- Lin-Marq, N.; Bontron, S.; Leupin, O.; Strubin, M. Hepatitis B Virus X Protein Interferes with Cell Viability through Interaction with the p127-kDa UV-Damaged DNA-Binding Protein. Virology 2001, 287, 266–274. [Google Scholar] [CrossRef]
- Leupin, O.; Bontron, S.; Schaeffer, C.; Strubin, M. Hepatitis B Virus X Protein Stimulates Viral Genome Replication via a DDB1-Dependent Pathway Distinct from That Leading to Cell Death. J. Virol. 2005, 79, 4238–4245. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.J.; Hyser, J.M.; Keasler, V.V.; Cang, Y.; Slagle, B.L. Hepatitis B virus regulatory HBx protein binding to DDB1 is required but is not sufficient for maximal HBV replication. Virology 2012, 426, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Decorsière, A.; Mueller, H.; Van Breugel, P.C.; Abdul, F.; Gerossier, L.; Beran, R.K.; Livingston, C.M.; Niu, C.; Fletcher, S.P.; Hantz, O.; et al. Hepatitis B virus X protein identifies the Smc5/6 complex as a host restriction factor. Nature 2016, 531, 386–389. [Google Scholar] [CrossRef]
- Murphy, C.M.; Xu, Y.; Li, F.; Nio, K.; Reszka-Blanco, N.; Li, X.; Wu, Y.; Yu, Y.; Xiong, Y.; Su, L. Hepatitis B Virus X Protein Promotes Degradation of SMC5/6 to Enhance HBV Replication. Cell Rep. 2016, 16, 2846–2854. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Livingston, C.M.; Li, L.; Beran, R.K.; Daffis, S.; Ramakrishnan, D.; Burdette, D.; Peiser, L.; Salas, E.; Ramos, H.; et al. The Smc5/6 Complex Restricts HBV when Localized to ND10 without Inducing an Innate Immune Response and Is Counteracted by the HBV X Protein Shortly after Infection. PLoS ONE 2017, 12, e0169648. [Google Scholar] [CrossRef]
- Rivière, L.; Quioc-Salomon, B.; Fallot, G.; Halgand, B.; Féray, C.; Buendia, M.A.; Neuveut, C. Hepatitis B virus replicating in hepatocellular carcinoma encodes HBx variants with preserved ability to antagonize restriction by Smc5/6. Antivir. Res. 2019, 172, 104618. [Google Scholar] [CrossRef]
- Belloni, L.; Pollicino, T.; De Nicola, F.; Guerrieri, F.; Raffa, G.; Fanciulli, M.; Raimondo, G.; Levrero, M. Nuclear HBx binds the HBV minichromosome and modifies the epigenetic regulation of cccDNA function. Proc. Natl. Acad. Sci. USA 2009, 106, 19975–19979. [Google Scholar] [CrossRef]
- Chong, C.K.; Cheng, C.Y.S.; Tsoi, S.Y.J.; Huang, F.-Y.; Liu, F.; Fung, J.; Seto, W.-K.; Lai, K.K.-Y.; Lai, C.-L.; Yuen, M.-F.; et al. HBV X protein mutations affect HBV transcription and association of histone-modifying enzymes with covalently closed circular DNA. Sci. Rep. 2020, 10, 802. [Google Scholar] [CrossRef]
- Benhenda, S.; Ducroux, A.; Rivière, L.; Sobhian, B.; Ward, M.D.; Dion, S.; Hantz, O.; Protzer, U.; Michel, M.-L.; Benkirane, M.; et al. Methyltransferase PRMT1 Is a Binding Partner of HBx and a Negative Regulator of Hepatitis B Virus Transcription. J. Virol. 2013, 87, 4360–4371. [Google Scholar] [CrossRef]
- Ducroux, A.; Benhenda, S.; Rivière, L.; Semmes, O.J.; Benkirane, M.; Neuveut, C. The Tudor Domain Protein Spindlin1 Is Involved in Intrinsic Antiviral Defense against Incoming Hepatitis B Virus and Herpes Simplex Virus Type 1. PLoS Pathog. 2014, 10, e1004343. [Google Scholar] [CrossRef]
- Rivière, L.; Gerossier, L.; Ducroux, A.; Dion, S.; Deng, Q.; Michel, M.-L.; Buendia, M.-A.; Hantz, O.; Neuveut, C. HBx relieves chromatin-mediated transcriptional repression of hepatitis B viral cccDNA involving SETDB1 histone methyltransferase. J. Hepatol. 2015, 63, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, V.; Hernández, S.; Rubio, L.; Alvarez, F.; Flores, Y.; Varas-Godoy, M.; De Ferrari, G.V.; Kann, M.; Villanueva, R.A.; Loyola, A. The enzymes LSD1 and Set1A cooperate with the viral protein HBx to establish an active hepatitis B viral chromatin state. Sci. Rep. 2016, 6, 25901. [Google Scholar] [CrossRef]
- Bouchard, M.J.; Wang, L.-H.; Schneider, R.J. Calcium Signaling by HBx Protein in Hepatitis B Virus DNA Replication. Science 2001, 294, 2376–2378. [Google Scholar] [CrossRef] [PubMed]
- McClain, S.L.; Clippinger, A.J.; Lizzano, R.; Bouchard, M.J. Hepatitis B Virus Replication Is Associated with an HBx-Dependent Mitochondrion-Regulated Increase in Cytosolic Calcium Levels. J. Virol. 2007, 81, 12061–12065. [Google Scholar] [CrossRef]
- Choi, Y.; Park, S.G.; Yoo, J.-H.; Jung, G. Calcium ions affect the hepatitis B virus core assembly. Virology 2005, 332, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, M.; Hiraga, N.; Akiyama, R.; Tanaka, S.; Matsushita, M.; Mitsui, F.; Abe, H.; Kitamura, S.; Hatakeyama, T.; Kimura, T.; et al. HBx protein is indispensable for development of viraemia in human hepatocyte chimeric mice. J. Gen. Virol. 2010, 91 Pt 7, 1854–1864. [Google Scholar] [CrossRef]
- Sitterlin, D.; Lee, T.H.; Prigent, S.; Tiollais, P.; Butel, J.S.; Transy, C. Interaction of the UV-damaged DNA-binding protein with hepatitis B virus X protein is conserved among mammalian hepadnaviruses and restricted to transactivation-proficient X-insertion mutants. J. Virol. 1997, 71, 6194–6199. [Google Scholar] [CrossRef]
- Kanno, T.; Berta, D.G.; Sjögren, C. The Smc5/6 Complex Is an ATP-Dependent Intermolecular DNA Linker. Cell Rep. 2015, 12, 1471–1482. [Google Scholar] [CrossRef]
- Choonnasard, A.; Shofa, M.; Okabayashi, T.; Saito, A. Conserved Functions of Orthohepadnavirus X Proteins to Inhibit Type-I Interferon Signaling. Int. J. Mol. Sci. 2024, 25, 3753. [Google Scholar] [CrossRef]
- Al-Qahtani, A.A.; Al-Anazi, M.R.; Nazir, N.; Ghai, R.; Abdo, A.A.; Sanai, F.M.; Al-Hamoudi, W.K.; Alswat, K.A.; Al-Ashgar, H.I.; Khan, M.Q.; et al. Hepatitis B virus (HBV) X gene mutations and their association with liver disease progression in HBV-infected patients. Oncotarget 2017, 8, 105115–105125. [Google Scholar] [CrossRef]
- Abdul, F.; Filleton, F.; Gerossier, L.; Paturel, A.; Hall, J.; Strubin, M.; Etienne, L. Smc5/6 Antagonism by HBx Is an Evolutionarily Conserved Function of Hepatitis B Virus Infection in Mammals. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Bock, C.T.; Schwinn, S.; Locarnini, S.; Fyfe, J.; Manns, M.P.; Trautwein, C.; Zentgraf, H. Structural organization of the hepatitis B virus minichromosome. J. Mol. Biol. 2001, 307, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Pollicino, T.; Belloni, L.; Raffa, G.; Pediconi, N.; Squadrito, G.; Raimondo, G.; Levrero, M. Hepatitis B Virus Replication Is Regulated by the Acetylation Status of Hepatitis B Virus cccDNA-Bound H3 and H4 Histones. Gastroenterology 2006, 130, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Newbold, J.E.; Xin, H.; Tencza, M.; Sherman, G.; Dean, J.; Bowden, S.; Locarnini, S. The covalently closed duplex form of the hepadnavirus genome exists in situ as a heterogeneous population of viral minichromosomes. J. Virol. 1995, 69, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Levrero, M.; Zucman-Rossi, J. Mechanisms of HBV-induced hepatocellular carcinoma. J. Hepatol. 2016, 64, S84–S101. [Google Scholar] [CrossRef]
- Diao, J.; Garces, R.; Richardson, C.D. X protein of hepatitis B virus modulates cytokine and growth factor related signal transduction pathways during the course of viral infections and hepatocarcinogenesis. Cytokine Growth Factor Rev. 2001, 12, 189–205. [Google Scholar] [CrossRef]
- Agustiningsih, A.; Rasyak, M.R.; Turyadi; Jayanti, S.; Sukowati, C. The oncogenic role of hepatitis B virus X gene in hepatocarcinogenesis: Recent updates. Explor. Target. Anti-Tumor Ther. 2024, 5, 120–134. [Google Scholar] [CrossRef]
- Slagle, B.L.; Andrisani, O.M.; Bouchard, M.J.; Lee, C.G.; Ou, J.J.; Siddiqui, A. Technical standards for hepatitis B virus X protein (HBx) research. Hepatology 2015, 61, 1416–1424. [Google Scholar] [CrossRef]
- Allweiss, L.; Giersch, K.; Pirosu, A.; Volz, T.; Muench, R.C.; Beran, R.K.; Urban, S.; Javanbakht, H.; Fletcher, S.P.; Lütgehetmann, M.; et al. Therapeutic shutdown of HBV transcripts promotes reappearance of the SMC5/6 complex and silencing of the viral genome in vivo. Gut 2022, 71, 372–381. [Google Scholar] [CrossRef]
- Han, Q.; Hou, Z.; Yin, C.; Zhang, C.; Zhang, J. 5′-triphosphate siRNA targeting HBx elicits a potent anti-HBV immune response in pAAV-HBV transfected mice. Antivir. Res. 2019, 161, 36–45. [Google Scholar] [CrossRef]
- Rossignol, J.F.; Bréchot, C. A Pilot Clinical Trial of Nitazoxanide in the Treatment of Chronic Hepatitis B. Hepatol. Commun. 2019, 3, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-T.; Hu, J.-L.; Ren, J.-H.; Yu, H.-B.; Zhong, S.; Wong, V.K.W.; Law, B.Y.K.; Chen, W.-X.; Xu, H.-M.; Zhang, Z.-Z.; et al. Dicoumarol, an NQO1 inhibitor, blocks cccDNA transcription by promoting degradation of HBx. J. Hepatol. 2021, 74, 522–534. [Google Scholar] [CrossRef]
- Tan, G.; Yi, Z.; Song, H.; Xu, F.; Li, F.; Aliyari, R.; Zhang, H.; Du, P.; Ding, Y.; Niu, J.; et al. Type-I-IFN-Stimulated Gene TRIM5γ Inhibits HBV Replication by Promoting HBx Degradation. Cell Rep. 2019, 29, 3551–3563.e3. [Google Scholar] [CrossRef]
- Bergametti, F.; Sitterlin, D.; Transy, C. Turnover of Hepatitis B Virus X Protein Is Regulated by Damaged DNA-Binding Complex. J. Virol. 2002, 76, 6495–6501. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.S.; Klote, L.; Aoki, N. Hepadnaviruses in cirrhotic liver and hepatocellular carcinoma. J. Med. Virol. 1990, 31, 18–32. [Google Scholar] [CrossRef]
- Gerin, J.L.; Cote, P.J.; Korba, B.E.; Tennant, B.C. Hepadnavirus-induced liver cancer in woodchucks. Cancer Detect. Prev. 1989, 14, 227–229. [Google Scholar]
- Pesavento, P.A.; Jackson, K.; Scase, T.; Tse, T.; Hampson, B.; Munday, J.S.; Barrs, V.R.; Beatty, J.A. A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses 2019, 11, 969. [Google Scholar] [CrossRef]
- Yang, C.; Ruan, P.; Ou, C.; Su, J.; Cao, J.; Luo, C.; Tang, Y.; Wang, Q.; Qin, H.; Sun, W.; et al. Chronic hepatitis B virus infection and occurrence of hepatocellular carcinoma in tree shrews (Tupaia belangeri chinensis). Virol. J. 2015, 12, 26. [Google Scholar] [CrossRef]
- Marion, P.L.; Van Davelaar, M.J.; Knight, S.S.; Salazar, F.H.; Garcia, G.; Popper, H.; Robinson, W.S. Hepatocellular Carcinoma in Ground Squirrels Persistently Infected with Ground Squirrel Hepatitis Virus. Proc. Natl. Acad. Sci. USA 1986, 83, 4543–4546. [Google Scholar] [CrossRef]
- Sivasudhan, E.; Blake, N.; Lu, Z.; Meng, J.; Rong, R. Hepatitis B Viral Protein HBx and the Molecular Mechanisms Modulating the Hallmarks of Hepatocellular Carcinoma: A Comprehensive Review. Cells 2022, 11, 741. [Google Scholar] [CrossRef] [PubMed]
- Allweiss, L.; Volz, T.; Giersch, K.; Kah, J.; Raffa, G.; Petersen, J.; Lohse, A.W.; Beninati, C.; Pollicino, T.; Urban, S.; et al. Proliferation of primary human hepatocytes and prevention of hepatitis B virus reinfection efficiently deplete nuclear cccDNA in vivo. Gut 2018, 67, 542–552. [Google Scholar] [CrossRef]
- Tu, T.; Zehnder, B.; Wettengel, J.M.; Zhang, H.; Coulter, S.; Ho, V.; Douglas, M.W.; Protzer, U.; George, J.; Urban, S. Mitosis of hepatitis B virus-infected cells in vitro results in uninfected daughter cells. JHEP Rep. 2022, 4, 100514. [Google Scholar] [CrossRef]
- Yan, Y.; Allweiss, L.; Yang, D.; Kang, J.; Wang, J.; Qian, X.; Zhang, T.; Liu, H.; Wang, L.; Liu, S.; et al. Down-regulation of cell membrane localized NTCP expression in proliferating hepatocytes prevents hepatitis B virus infection. Emerg. Microbes Infect. 2019, 8, 879–894. [Google Scholar] [CrossRef]
- Werle-Lapostolle, B.; Bowden, S.; Locarnini, S.; Wursthorn, K.; Petersen, J.; Lau, G.; Trepo, C.; Marcellin, P.; Goodman, Z.; Delaney, W.E., IV; et al. Persistence of cccDNA during the natural history of chronic hepatitis B and decline during adefovir dipivoxil therapy: cccDNA levels in chronic hepatitis B patients. Gastroenterology 2004, 126, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Bréchot, C.; Gozuacik, D.; Murakami, Y.; Paterlini-Bréchot, P. Molecular bases for the development of hepatitis B virus (HBV)-related hepatocellular carcinoma (HCC). Semin. Cancer Biol. 2000, 10, 211–231. [Google Scholar] [CrossRef]
- Hayashi, S.; Isogawa, M.; Kawashima, K.; Ito, K.; Chuaypen, N.; Morine, Y.; Shimada, M.; Higashi-Kuwata, N.; Watanabe, T.; Tangkijvanich, P.; et al. Droplet digital PCR assay provides intrahepatic HBV cccDNA quantification tool for clinical application. Sci. Rep. 2022, 12, 2133. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Deng, H.; Zhen, X.; Xiong, J.; Hu, Y. Quantification of intrahepatic cccDNA in HBV associated hepatocellular carcinoma by improved ddPCR method. J. Virol. Methods 2022, 299, 114334. [Google Scholar] [CrossRef]
- Lizzano, R.A.; Yang, B.; Clippinger, A.J.; Bouchard, M.J. The C-terminal region of the hepatitis B virus X protein is essential for its stability and function. Virus Res. 2011, 155, 231–239. [Google Scholar] [CrossRef]
- Ma, N.-F.; Lau, S.H.; Hu, L.; Xie, D.; Wu, J.; Yang, J.; Wang, Y.; Wu, M.-C.; Fung, J.; Bai, X.; et al. COOH-Terminal Truncated HBV X Protein Plays Key Role in Hepatocarcinogenesis. Clin. Cancer Res. 2008, 14, 5061–5068. [Google Scholar] [CrossRef]
- Sachs, A. The role of poly(A) in the translation and stability of mRNA. Curr. Opin. Cell Biol. 1990, 2, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Russnak, R.; Ganem, D. Sequences 5’ to the polyadenylation signal mediate differential poly(A) site use in hepatitis B viruses. Genes Dev. 1990, 4, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, Q.; Gong, L.; Xu, H.; Liu, B.; Fang, X.; Yu, D.; Li, L.; Wei, T.; Wang, Y.; et al. C-terminal truncated HBx initiates hepatocarcinogenesis by downregulating TXNIP and reprogramming glucose metabolism. Oncogene 2021, 40, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.-Y.; Chai, S.; Tong, M.; Guan, X.-Y.; Lin, C.-H.; Ching, Y.P.; Xie, D.; Cheng, A.S.L.; Ma, S.K.Y. C-terminal truncated hepatitis B virus X protein promotes hepatocellular carcinogenesis through induction of cancer and stem cell-like properties. Oncotarget 2016, 7, 24005–24017. [Google Scholar] [CrossRef]
- Sze, K.M.F.; Chu, G.K.; Lee, J.M.; Ng, I.O. C-terminal truncated hepatitis B virus x protein is associated with metastasis and enhances invasiveness by c-jun/matrix metalloproteinase protein 10 activation in hepatocellular carcinoma. Hepatology 2013, 57, 131–139. [Google Scholar] [CrossRef]
- Ching, R.H.H.; Sze, K.M.F.; Lau, E.Y.T.; Chiu, Y.-T.; Lee, J.M.F.; Ng, I.O.L.; Lee, T.K.W. C-terminal truncated hepatitis B virus X protein regulates tumorigenicity, self-renewal and drug resistance via STAT3/Nanog signaling pathway. Oncotarget 2017, 8, 23507–23516. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, W.; Liu, Q.; Zhang, X.; Lv, N.; Ye, L.; Zhang, X. A Mutant of Hepatitis B Virus X Protein (HBxΔ127) Promotes Cell Growth through A Positive Feedback Loop Involving 5-Lipoxygenase and Fatty Acid Synthase. Neoplasia 2010, 12, 103–115, IN1–IN3. [Google Scholar] [CrossRef]
- Mao, X.; Tey, S.K.; Ko, F.C.; Kwong, E.M.; Gao, Y.; Ng, I.O.; Cheung, S.T.; Guan, X.Y.; Yam, J.W. C-terminal truncated HBx protein activates caveolin-1/LRP6/β-catenin/FRMD5 axis in promoting hepatocarcinogenesis. Cancer Lett. 2019, 444, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, M.; Liao, D.; Lu, X.; Gu, X.; Zhang, Q.; Zhang, Z.; Li, H. Carboxyl-terminal truncated HBx contributes to invasion and metastasis via deregulating metastasis suppressors in hepatocellular carcinoma. Oncotarget 2016, 7, 55110–55127. [Google Scholar] [CrossRef]
- Montalbano, R.; Honrath, B.; Wissniowski, T.T.; Elxnat, M.; Roth, S.; Ocker, M.; Quint, K.; Churin, Y.; Roederfeld, M.; Schroeder, D.; et al. Exogenous hepatitis B virus envelope proteins induce endoplasmic reticulum stress: Involvement of cannabinoid axis in liver cancer cells. Oncotarget 2016, 7, 20312–20323. [Google Scholar] [CrossRef]
- Jung, S.-Y.; Kim, Y.-J. C-terminal region of HBx is crucial for mitochondrial DNA damage. Cancer Lett. 2013, 331, 76–83. [Google Scholar] [CrossRef]
- Wu, X.; Ni, Z.; Song, T.; Lv, W.; Chen, Y.; Huang, D.; Xie, Y.; Huang, W.; Niu, Y. C-Terminal Truncated HBx Facilitates Oncogenesis by Modulating Cell Cycle and Glucose Metabolism in FXR-Deficient Hepatocellular Carcinoma. Int. J. Mol. Sci. 2023, 24, 5174. [Google Scholar] [CrossRef]
- Becker, S.A.; Lee, T.-H.; Butel, J.S.; Slagle, B.L. Hepatitis B Virus X Protein Interferes with Cellular DNA Repair. J. Virol. 1998, 72, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.M.; Murray, J.M. Smc5/6: A link between DNA repair and unidirectional replication? Nature reviews. Mol. Cell Biol. 2008, 9, 177–182. [Google Scholar]
- Atkins, A.; Xu, M.J.; Li, M.; Rogers, N.P.; Pryzhkova, M.V.; Jordan, P.W. SMC5/6 is required for replication fork stability and faithful chromosome segregation during neurogenesis. eLife 2020, 9, e61171. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Jordan, P.W. SMC5/6 Promotes Replication Fork Stability via Negative Regulation of the COP9 Signalosome. Int. J. Mol. Sci. 2024, 25, 952. [Google Scholar] [CrossRef]
- Moradi-Fard, S.; Sarthi, J.; Tittel-Elmer, M.; Lalonde, M.; Cusanelli, E.; Chartrand, P.; Cobb, J.A. Smc5/6 Is a Telomere-Associated Complex that Regulates Sir4 Binding and TPE. PLoS Genet. 2016, 12, e1006268. [Google Scholar] [CrossRef]
- Sekiba, K.; Otsuka, M.; Funato, K.; Miyakawa, Y.; Tanaka, E.; Seimiya, T.; Yamagami, M.; Tsutsumi, T.; Okushin, K.; Miyakawa, K.; et al. HBx-induced degradation of Smc5/6 complex impairs homologous recombination-mediated repair of damaged DNA. J. Hepatol. 2022, 76, 53–62. [Google Scholar] [CrossRef]
- Lee, T.H.; Finegold, M.J.; Shen, R.F.; DeMayo, J.L.; Woo, S.L.; Butel, J.S. Hepatitis B virus transactivator X protein is not tumorigenic in transgenic mice. J. Virol. 1990, 64, 5939–5947. [Google Scholar] [CrossRef] [PubMed]
- Jacome, A.; Gutierrez-Martinez, P.; Schiavoni, F.; Tenaglia, E.; Martinez, P.; Rodríguez-Acebes, S.; Lecona, E.; Murga, M.; Méndez, J.; Blasco, M.A.; et al. NSMCE2 suppresses cancer and aging in mice independently of its SUMO ligase activity. EMBO J. 2015, 34, 2604–2619. [Google Scholar] [CrossRef]
- Groisman, I.J.; Koshy, R.; Henkler, F.; Groopman, J.D.; Alaoui-Jamali, M.A. Downregulation of DNA excision repair by the hepatitis B virus-x protein occurs in p53-proficient and p53-deficient cells. Carcinogenesis 1999, 20, 479–483. [Google Scholar] [CrossRef]
- Arzumanyan, A.; Reis, H.M.G.P.V.; Feitelson, M.A. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat. Rev. Cancer 2013, 13, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-M.; Lee, S.-Y.; Kim, B.-J. Naturally Occurring Hepatitis B Virus Mutations Leading to Endoplasmic Reticulum Stress and Their Contribution to the Progression of Hepatocellular Carcinoma. Int. J. Mol. Sci. 2019, 20, 597. [Google Scholar] [CrossRef]
- Yip, K.M.; Fischer, N.; Paknia, E.; Chari, A.; Stark, H. Atomic-resolution protein structure determination by cryo-EM. Nature 2020, 587, 157–161. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Iavarone, M.; Trabut, J.-B.; Delpuech, O.; Carnot, F.; Colombo, M.; Kremsdorf, D.; Bréchot, C.; Thiers, V. Characterisation of hepatitis B virus X protein mutants in tumour and non-tumour liver cells using laser capture microdissection. J. Hepatol. 2003, 39, 253–261. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).