
Production and Characterization of Self-Assembled Virus-like Particles Comprising Capsid Proteins from Genotypes 3 and 4 Hepatitis E Virus (HEV) and Rabbit HEV Expressed in Escherichia coli

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and Truncated HEV ORF2 Protein Expression
2.2. Purification of Recombinant HEV ORF2 Proteins
2.3. SDS-PAGE and Western Blot Analysis
2.4. Analysis of Purified HEV VLPs with the Bioanalyzer
2.5. Transmission Electron Microscopy (TEM)
2.6. Immunoelectron Microscopy (IEM)
2.7. Enzyme-Linked Immunosorbent Assays (ELISAs) for the Determination of Binding with Monoclonal Antibodies (MAbs) against ORF2 Protein of Human HEV (HEV-3 and HEV-4), Rabbit HEV (HEV-3ra), or Rat HEV (HEV-C1)
2.8. Immunization of HEV VLPs in Mice
3. Results
3.1. The Expression and Purification of Truncated HEV ORF2 Proteins
3.2. Characterization of the Purified VLPs of HEV-3, HEV-3ra, and HEV-4
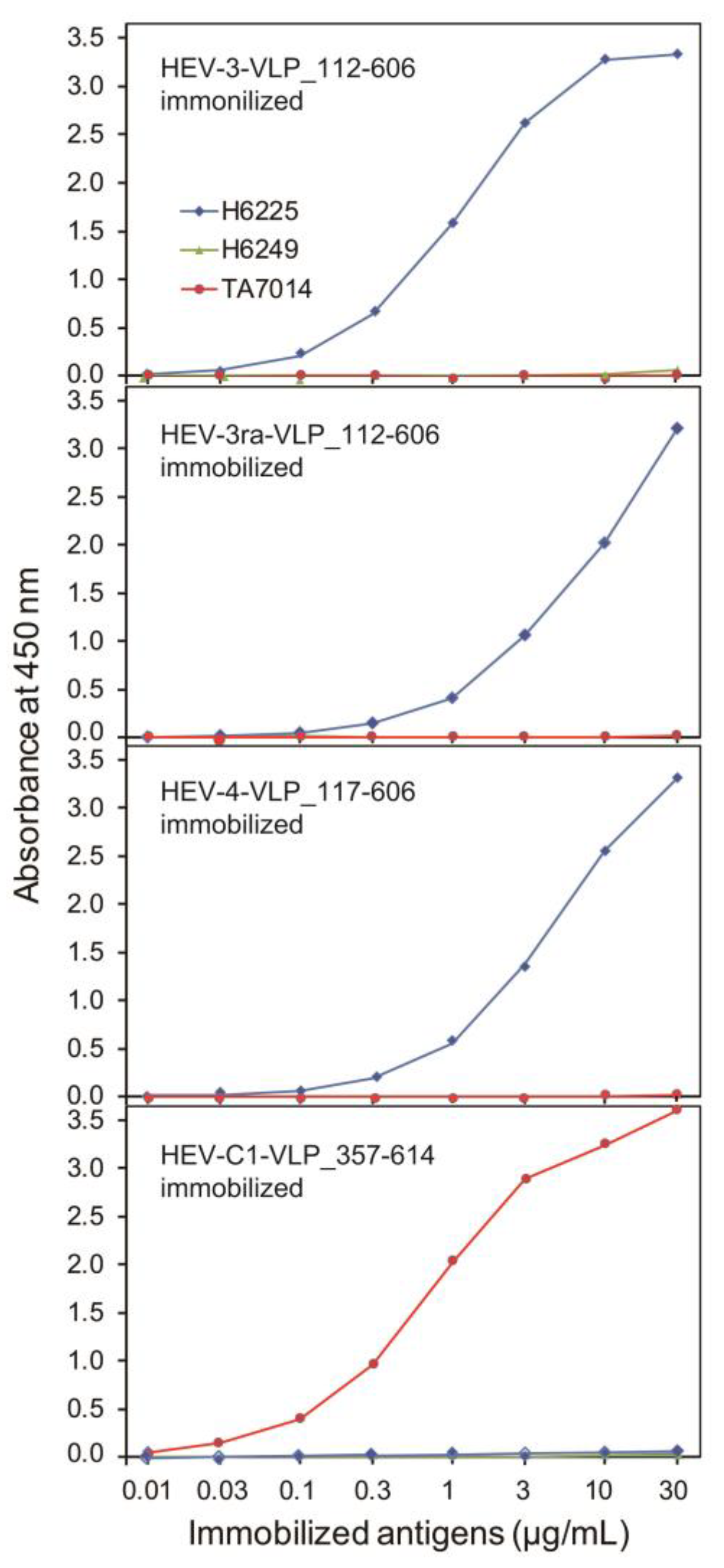
3.3. The Antigenicity of the Purified VLPs of HEV-3, HEV-3ra, and HEV-4
3.4. Immune Response of the Balb/cAJcl Mice to the Purified VLPs of HEV-3, HEV-3ra, and HEV-4
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef]
- Li, P.; Liu, J.; Li, Y.; Su, J.; Ma, Z.; Bramer, W.M.; Cao, W.; de Man, R.A.; Peppelenbosch, M.P.; Pan, Q. The global epidemiology of hepatitis E virus infection: A systematic review and meta-analysis. Liver Int. 2020, 40, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Damiris, K.; Aghaie Meybodi, M.; Niazi, M.; Pyrsopoulos, N. Hepatitis E in immunocompromised individuals. World J. Hepatol. 2022, 14, 482–494. [Google Scholar] [CrossRef]
- Pischke, S.; Hartl, J.; Pas, S.D.; Lohse, A.W.; Jacobs, B.C.; Van der Eijk, A.A. Hepatitis E virus: Infection beyond the liver? J. Hepatol. 2017, 66, 1082–1095. [Google Scholar] [CrossRef] [PubMed]
- Berglov, A.; Hallager, S.; Weis, N. Hepatitis E during pregnancy: Maternal and foetal case-fatality rates and adverse outcomes-A systematic review. J. Viral Hepat. 2019, 26, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gracia, M.T.; Suay-Garcia, B.; Mateos-Lindemann, M.L. Hepatitis E and pregnancy: Current state. Rev. Med. Virol. 2017, 27, e1929. [Google Scholar] [CrossRef]
- Grange, Z.L.; Goldstein, T.; Johnson, C.K.; Anthony, S.; Gilardi, K.; Daszak, P.; Olival, K.J.; O’Rourke, T.; Murray, S.; Olson, S.H.; et al. Ranking the risk of animal-to-human spillover for newly discovered viruses. Proc. Natl. Acad. Sci. USA 2021, 118, e2002324118. [Google Scholar] [CrossRef]
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Characterization of the Quasi-Enveloped Hepatitis E Virus Particles Released by the Cellular Exosomal Pathway. J. Virol. 2017, 91, e00822-17. [Google Scholar] [CrossRef]
- Purdy, M.A.; Drexler, J.F.; Meng, X.J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus Taxonomy Profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Structural and molecular biology of hepatitis E virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Nimgaonkar, I.; Ding, Q.; Schwartz, R.E.; Ploss, A. Hepatitis E virus: Advances and challenges. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 96–110. [Google Scholar] [CrossRef]
- Nan, Y.; Zhang, Y.J. Molecular Biology and Infection of Hepatitis E Virus. Front. Microbiol. 2016, 7, 1419. [Google Scholar] [CrossRef]
- Graff, J.; Torian, U.; Nguyen, H.; Emerson, S.U. A bicistronic subgenomic mRNA encodes both the ORF2 and ORF3 proteins of hepatitis E virus. J. Virol. 2006, 80, 5919–5926. [Google Scholar] [CrossRef]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef]
- Zhou, Y. Immunobiology and Host Response to HEV. Adv. Exp. Med. Biol. 2016, 948, 113–141. [Google Scholar] [PubMed]
- Glitscher, M.; Hildt, E. Hepatitis E virus egress and beyond—The manifold roles of the viral ORF3 protein. Cell Microbiol. 2021, 23, e13379. [Google Scholar] [CrossRef]
- Nagashima, S.; Jirintai, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Kouki, T.; Yashiro, T.; Okamoto, H. Hepatitis E virus egress depends on the exosomal pathway, with secretory exosomes derived from multivesicular bodies. J. Gen. Virol. 2014, 95, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.J.; Renou, C. Zoonotic hepatitis E: Animal reservoirs and emerging risks. Vet. Res. 2010, 41, 46. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Hepatitis E virus: Host tropism and zoonotic infection. Curr. Opin. Microbiol. 2020, 59, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar]
- Takahashi, M.; Nishizawa, T.; Nagashima, S.; Jirintai, S.; Kawakami, M.; Sonoda, Y.; Suzuki, T.; Yamamoto, S.; Shigemoto, K.; Ashida, K.; et al. Molecular characterization of a novel hepatitis E virus (HEV) strain obtained from a wild boar in Japan that is highly divergent from the previously recognized HEV strains. Virus Res. 2014, 180, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nishizawa, T.; Sato, Y.; Miyazaki, S.; Aikawa, T.; Ashida, K.; Tamaru, T.; Oguro, K.; Hayakawa, F.; Matsuoka, H.; et al. Prevalence and genotype/subtype distribution of hepatitis E virus (HEV) among wild boars in Japan: Identification of a genotype 5 HEV strain. Virus Res. 2020, 287, 198106. [Google Scholar] [CrossRef]
- Primadharsini, P.P.; Takahashi, M.; Nishizawa, T.; Sato, Y.; Nagashima, S.; Murata, K.; Okamoto, H. The Full-Genome Analysis and Generation of an Infectious cDNA Clone of a Genotype 6 Hepatitis E Virus Variant Obtained from a Japanese Wild Boar: In Vitro Cultivation in Human Cell Lines. Viruses 2024, 16, 842. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef]
- Nishizawa, T.; Takahashi, M.; Tsatsralt-Od, B.; Nyamdavaa, K.; Dulmaa, N.; Osorjin, B.; Tseren-Ochir, E.O.; Sharav, T.; Bayasgalan, C.; Sukhbaatar, B.; et al. Identification and a full genome analysis of novel camel hepatitis E virus strains obtained from Bactrian camels in Mongolia. Virus Res. 2021, 299, 198355. [Google Scholar] [CrossRef]
- Hara, Y.; Terada, Y.; Yonemitsu, K.; Shimoda, H.; Noguchi, K.; Suzuki, K.; Maeda, K. High prevalence of hepatitis E virus in wild boar (Sus scrofa) in Yamaguchi Prefecture, Japan. J. Wildl. Dis. 2014, 50, 378–383. [Google Scholar] [CrossRef]
- Nakano, T.; Takahashi, K.; Arai, M.; Okano, H.; Kato, H.; Ayada, M.; Okamoto, H.; Mishiro, S. Identification of European-type hepatitis E virus subtype 3e isolates in Japanese wild boars: Molecular tracing of HEV from swine to wild boars. Infect. Genet. Evol. 2013, 18, 287–298. [Google Scholar] [CrossRef]
- Okamoto, H. Genetic variability and evolution of hepatitis E virus. Virus Res. 2007, 127, 216–228. [Google Scholar] [CrossRef]
- Sato, Y.; Sato, H.; Naka, K.; Furuya, S.; Tsukiji, H.; Kitagawa, K.; Sonoda, Y.; Usui, T.; Sakamoto, H.; Yoshino, S.; et al. A nationwide survey of hepatitis E virus (HEV) infection in wild boars in Japan: Identification of boar HEV strains of genotypes 3 and 4 and unrecognized genotypes. Arch. Virol. 2011, 156, 1345–1358. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Wang, L. An overview: Rabbit hepatitis E virus (HEV) and rabbit providing an animal model for HEV study. Rev. Med. Virol. 2018, 28, e1961. [Google Scholar] [CrossRef] [PubMed]
- Jirintai, S.; Jinshan; Tanggis; Manglai, D.; Mulyanto; Takahashi, M.; Nagashima, S.; Kobayashi, T.; Nishizawa, T.; Okamoto, H. Molecular analysis of hepatitis E virus from farm rabbits in Inner Mongolia, China and its successful propagation in A549 and PLC/PRF/5 cells. Virus Res. 2012, 170, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Abravanel, F.; Lhomme, S.; El Costa, H.; Schvartz, B.; Peron, J.M.; Kamar, N.; Izopet, J. Rabbit Hepatitis E Virus Infections in Humans, France. Emerg. Infect. Dis. 2017, 23, 1191–1193. [Google Scholar] [CrossRef]
- Izopet, J.; Dubois, M.; Bertagnoli, S.; Lhomme, S.; Marchandeau, S.; Boucher, S.; Kamar, N.; Abravanel, F.; Guerin, J.L. Hepatitis E virus strains in rabbits and evidence of a closely related strain in humans, france. Emerg. Infect. Dis. 2012, 18, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.Y.; Wu, S.; Cai, J.; Zhang, A.J.; Leung, K.H.; Chung, T.W.H.; Chan, J.F.W.; Chan, W.M.; Teng, J.L.L.; et al. Rat Hepatitis E Virus as Cause of Persistent Hepatitis after Liver Transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W.; et al. Transmission of Rat Hepatitis E Virus Infection to Humans in Hong Kong: A Clinical and Epidemiological Analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef]
- Caballero-Gomez, J.; Pereira, S.; Rivero-Calle, I.; Perez, A.B.; Viciana, I.; Casares-Jimenez, M.; Rios-Munoz, L.; Rivero-Juarez, A.; Aguilera, A.; Rivero, A. Acute Hepatitis in Children Due to Rat Hepatitis E Virus. J. Pediatr. 2024, 273, 114125. [Google Scholar] [CrossRef]
- Johne, R.; Heckel, G.; Plenge-Bönig, A.; Kindler, E.; Maresch, C.; Reetz, J.; Schielke, A.; Ulrich, R.G. Novel hepatitis E virus genotype in Norway rats, Germany. Emerg. Infect. Dis. 2010, 16, 1452–1455. [Google Scholar] [CrossRef]
- Mulyanto; Wibawa, I.D.; Suparyatmo, J.B.; Amirudin, R.; Ohnishi, H.; Takahashi, M.; Nishizawa, T.; Okamoto, H. The complete genomes of subgenotype IA hepatitis A virus strains from four different islands in Indonesia form a phylogenetic cluster. Arch. Virol. 2014, 159, 935–945. [Google Scholar] [CrossRef]
- Purcell, R.H.; Engle, R.E.; Rood, M.P.; Kabrane-Lazizi, Y.; Nguyen, H.T.; Govindarajan, S.; St Claire, M.; Emerson, S.U. Hepatitis E virus in rats, Los Angeles, California, USA. Emerg. Infect. Dis. 2011, 17, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Schofield, D.J.; Glamann, J.; Emerson, S.U.; Purcell, R.H. Identification by phage display and characterization of two neutralizing chimpanzee monoclonal antibodies to the hepatitis E virus capsid protein. J. Virol. 2000, 74, 5548–5555. [Google Scholar] [CrossRef] [PubMed]
- Mazalovska, M.; Kouokam, J.C. Progress in the Production of Virus-Like Particles for Vaccination against Hepatitis E Virus. Viruses 2020, 12, 826. [Google Scholar] [CrossRef]
- Hadj Hassine, I.; Ben M’hadheb, M.; Almalki, M.A.; Gharbi, J. Virus-like particles as powerful vaccination strategy against human viruses. Rev. Med. Virol. 2024, 34, e2498. [Google Scholar] [CrossRef]
- Cao, Y.F.; Tao, H.; Hu, Y.M.; Shi, C.B.; Wu, X.; Liang, Q.; Chi, C.P.; Li, L.; Liang, Z.L.; Meng, J.H.; et al. A phase 1 randomized open-label clinical study to evaluate the safety and tolerability of a novel recombinant hepatitis E vaccine. Vaccine 2017, 35, 5073–5080. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Takeda, N.; Miyamura, T.; Matsuura, Y.; Wang, J.C.; Engvall, H.; Hammar, L.; Xing, L.; Cheng, R.H. Essential elements of the capsid protein for self-assembly into empty virus-like particles of hepatitis E virus. J. Virol. 2005, 79, 12999–13006. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Zhao, Q.; Wu, T.; Chen, S.; Zhang, J.; Xia, N.S. The development of a recombinant hepatitis E vaccine HEV 239. Hum. Vaccin. Immunother. 2015, 11, 908–914. [Google Scholar] [CrossRef]
- Li, T.C.; Yamakawa, Y.; Suzuki, K.; Tatsumi, M.; Razak, M.A.; Uchida, T.; Takeda, N.; Miyamura, T. Expression and self-assembly of empty virus-like particles of hepatitis E virus. J. Virol. 1997, 71, 7207–7213. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, M.; Pan, H.; Lin, Z.; Wang, K.; Weng, Z.; Zhu, Y.; Xin, L.; Zhang, J.; Li, S.; et al. Robust manufacturing and comprehensive characterization of recombinant hepatitis E virus-like particles in Hecolin((R)). Vaccine 2014, 32, 4039–4050. [Google Scholar] [CrossRef]
- Zheng, M.; Jiang, J.; Zhang, X.; Wang, N.; Wang, K.; Li, Q.; Li, T.; Lin, Q.; Wang, Y.; Yu, H.; et al. Characterization of capsid protein (p495) of hepatitis E virus expressed in Escherichia coli and assembling into particles in vitro. Vaccine 2018, 36, 2104–2111. [Google Scholar] [CrossRef]
- Kobayashi, T.; Takahashi, M.; Ohta, S.; Nagashima, S.; Primadharsini, P.P.; Mulyanto; Kunita, S.; Murata, K.; Okamoto, H. Production of capsid proteins of rat hepatitis E virus in Escherichia coli and characterization of self-assembled virus-like particles. Virus Res. 2021, 302, 198483. [Google Scholar] [CrossRef] [PubMed]
- Abravanel, F.; Lhomme, S. Hecolin vaccine: Long-term efficacy against HEV for a three-dose regimen. Lancet 2024, 403, 782–783. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for Hepatitis E virus. J. Gen. Virol. 2007, 88, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Takahashi, M.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Construction of an infectious cDNA clone of hepatitis E virus strain JE03-1760F that can propagate efficiently in cultured cells. J. Gen. Virol. 2009, 90, 457–462. [Google Scholar] [CrossRef]
- Yamashita, T.; Mori, Y.; Miyazaki, N.; Cheng, R.H.; Yoshimura, M.; Unno, H.; Shima, R.; Moriishi, K.; Tsukihara, T.; Li, T.C.; et al. Biological and immunological characteristics of hepatitis E virus-like particles based on the crystal structure. Proc. Natl. Acad. Sci. USA 2009, 106, 12986–12991. [Google Scholar] [CrossRef]
- Tanaka, T.; Takahashi, M.; Takahashi, H.; Ichiyama, K.; Hoshino, Y.; Nagashima, S.; Mizuo, H.; Okamoto, H. Development and Characterization of a Genotype 4 Hepatitis E Virus Cell Culture System Using a HE-JF5/15F Strain Recovered from a Fulminant Hepatitis Patient. J. Clin. Microbiol. 2009, 47, 1906–1910. [Google Scholar] [CrossRef]
- Takahashi, M.; Hoshino, Y.; Tanaka, T.; Takahashi, H.; Nishizawa, T.; Okamoto, H. Production of monoclonal antibodies against hepatitis E virus capsid protein and evaluation of their neutralizing activity in a cell culture system. Arch. Virol. 2008, 153, 657–666. [Google Scholar] [CrossRef]
- Nishiyama, T.; Umezawa, K.; Yamada, K.; Takahashi, M.; Kunita, S.; Mulyanto; Kii, I.; Okamoto, H. The Capsid (ORF2) Protein of Hepatitis E Virus in Feces Is C-Terminally Truncated. Pathogens 2022, 11, 24. [Google Scholar] [CrossRef]
- Kobayashi, T.; Takahashi, M.; Tanggis; Mulyanto; Jirintai, S.; Nagashima, S.; Nishizawa, T.; Okamoto, H. Characterization and epitope mapping of monoclonal antibodies raised against rat hepatitis E virus capsid protein: An evaluation of their neutralizing activity in a cell culture system. J. Virol. Methods 2016, 233, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Mizuo, H.; Suzuki, K.; Takikawa, Y.; Sugai, Y.; Tokita, H.; Akahane, Y.; Itoh, K.; Gotanda, Y.; Takahashi, M.; Nishizawa, T.; et al. Polyphyletic strains of hepatitis E virus are responsible for sporadic cases of acute hepatitis in Japan. J. Clin. Microbiol. 2002, 40, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tang, X.; Seetharaman, J.; Yang, C.; Gu, Y.; Zhang, J.; Du, H.; Shih, J.W.; Hew, C.L.; Sivaraman, J.; et al. Dimerization of hepatitis E virus capsid protein E2s domain is essential for virus-host interaction. PLoS. Pathog. 2009, 5, e1000537. [Google Scholar] [CrossRef]
- Zhang, M.; Emerson, S.U.; Nguyen, H.; Engle, R.E.; Govindarajan, S.; Gerin, J.L.; Purcell, R.H. Immunogenicity and protective efficacy of a vaccine prepared from 53 kDa truncated hepatitis E virus capsid protein expressed in insect cells. Vaccine 2001, 20, 853–857. [Google Scholar] [CrossRef]
- Liu, Z.; Behloul, N.; Baha, S.; Wei, W.; Tao, W.; Zhang, T.; Li, W.; Shi, R.; Meng, J. Role of the C-terminal cysteines in virus-like particle formation and oligomerization of the hepatitis E virus ORF2 truncated proteins. Virology 2020, 544, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Zhang, Z.; Li, S.; Zhang, J.; Xia, N.; Zhao, Q. Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation. Viruses 2020, 12, 109. [Google Scholar] [CrossRef]
- Vormittag, P.; Klamp, T.; Hubbuch, J. Ensembles of Hydrophobicity Scales as Potent Classifiers for Chimeric Virus-Like Particle Solubility—An Amino Acid Sequence-Based Machine Learning Approach. Front. Bioeng. Biotechnol. 2020, 8, 395. [Google Scholar] [CrossRef] [PubMed]
- Thrane, S.; Janitzek, C.M.; Matondo, S.; Resende, M.; Gustavsson, T.; de Jongh, W.A.; Clemmensen, S.; Roeffen, W.; van de Vegte-Bolmer, M.; van Gemert, G.J.; et al. Bacterial superglue enables easy development of efficient virus-like particle based vaccines. J. Nanobiotechnol. 2016, 14, 30. [Google Scholar] [CrossRef]
- Li, T.C.; Yoshimatsu, K.; Yasuda, S.P.; Arikawa, J.; Koma, T.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Mai le, T.Q.; Hoa, N.T.; et al. Characterization of self-assembled virus-like particles of rat hepatitis E virus generated by recombinant baculoviruses. J. Gen. Virol. 2011, 92, 2830–2837. [Google Scholar] [CrossRef]
- Li, T.C.; Yang, T.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Ishii, K.; Haga, K.; Nakamura, T.; Ochiai, S.; Takaji, W.; et al. Construction and characterization of an infectious cDNA clone of rat hepatitis E virus. J. Gen. Virol. 2015, 96, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Kishida, N.; Shirakura, M.; Imai, M.; Asanuma, H.; Takeda, N.; Wakita, T.; et al. Characterization of self-assembled virus-like particles of ferret hepatitis E virus generated by recombinant baculoviruses. J. Gen. Virol. 2013, 94, 2647–2656. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Kataoka, M.; Liu, Z.; Takeda, N.; Wakita, T.; Li, T.C. Characterization of self-assembled virus-like particles of dromedary camel hepatitis e virus generated by recombinant baculoviruses. Virus Res. 2015, 210, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.C. Immunogenicity and Antigenicity of Rabbit Hepatitis E Virus-Like Particles Produced by Recombinant Baculoviruses. Viruses 2021, 13, 1573. [Google Scholar] [CrossRef] [PubMed]
- Emerson, S.U.; Clemente-Casares, P.; Moiduddin, N.; Arankalle, V.A.; Torian, U.; Purcell, R.H. Putative neutralization epitopes and broad cross-genotype neutralization of Hepatitis E virus confirmed by a quantitative cell-culture assay. J. Gen. Virol. 2006, 87, 697–704. [Google Scholar] [CrossRef]
- Engle, R.E.; Yu, C.; Emerson, S.U.; Meng, X.J.; Purcell, R.H. Hepatitis E virus (HEV) capsid antigens derived from viruses of human and swine origin are equally efficient for detecting anti-HEV by enzyme immunoassay. J. Clin. Microbiol. 2002, 40, 4576–4580. [Google Scholar] [CrossRef]
- Li, T.C.; Kataoka, M.; Takahashi, K.; Yoshizaki, S.; Kato, T.; Ishii, K.; Takeda, N.; Mishiro, S.; Wakita, T. Generation of hepatitis E virus-like particles of two new genotypes G5 and G6 and comparison of antigenic properties with those of known genotypes. Vet. Microbiol. 2015, 178, 150–157. [Google Scholar] [CrossRef]
- Mulyanto; Suparyatmo, J.B.; Andayani, I.; Khalid; Takahashi, M.; Ohnishi, H.; Jirintai, S.; Nagashima, S.; Nishizawa, T.; Okamoto, H. Marked genomic heterogeneity of rat hepatitis E virus strains in Indonesia demonstrated on a full-length genome analysis. Virus Res. 2014, 179, 102–112. [Google Scholar] [CrossRef]
- Sridhar, S.; Situ, J.; Cai, J.P.; Yip, C.C.; Wu, S.; Zhang, A.J.; Wen, L.; Chew, N.F.; Chan, W.M.; Poon, R.W.; et al. Multimodal investigation of rat hepatitis E virus antigenicity: Implications for infection, diagnostics, and vaccine efficacy. J. Hepatol. 2021, 74, 1315–1324. [Google Scholar] [CrossRef]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Muramatsu, M.; Li, T.C. Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus. Viruses 2022, 14, 1125. [Google Scholar] [CrossRef]
- Xu, L.D.; Zhang, F.; Chen, C.; Peng, L.; Luo, W.T.; Chen, R.; Xu, P.; Huang, Y.W. Revisiting the Mongolian Gerbil Model for Hepatitis E Virus by Reverse Genetics. Microbiol. Spectr. 2022, 10, e0219321. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Note |
|---|---|---|
| hHEV_G3 Nde-ORF2 aa12 Fw | CATATG GTGCTTCTGCCTATGCTGCC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa14 Fw | CATATG CTGCCTATGCTGCCCGCGCC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa61 Fw | CATATG CCCTTTGCCGCCGATGTCG | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa89 Fw | CATATG GACCAGTCCCAGCGCCCCTC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa107 Fw | CATATG GCTGCGCCGTTGACTGCTATC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa112 Fw | CATATG GCTATCTCACCAGCCCCTGAC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa117 Fw | CATATG CCTGACACAGCCCCTGTACC | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa122 Fw | CATATG GTACCTGATGTTGATTCACG | Nde I site (underlined) |
| hHEV_G3 Nde-ORF2 aa368 Fw | CATATG ATTGCTCTGACACTGTTCAAC | Nde I site (underlined) |
| rbHEV_G3 Nde-ORF2 aa112 Fw | CATATG GCTGTTTCACCTGCACCTG | Nde I site (underlined) |
| rbHEV_G3 Nde-ORF2 aa368 Fw | CATATG ATAGCCCTGACGCTGTTTAAC | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa112 Fw | CATATG GCTGTGGCCCCGGCCCCCGA | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa114 Fw | CATATG GCCCCGGCCCCCGATACTGC | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa117 Fw | CATATG CCCGATACTGCTCCTGTTCC | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa366 Fw | CATATG GTCGGTCGTGGTATAGCGC | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa368 Fw | CATATG CGTGGTATAGCGCTAACTCTG | Nde I site (underlined) |
| hHEV_G4 Nde-ORF2 aa370 Fw | CATATG ATAGCGCTAACTCTGTTCAATC | Nde I site (underlined) |
| hHEV_G3 ORF2-Bam aa606 Rev | GGATCC CTA AGCCGAGTGCGGGGCTAGTAC | BamH I site (underlined) |
| rbHEV_G3 ORF2-Bam aa606 Rev | GGATCC CTA AACTGAGTGCGGAGCAAGC | BamH I site (underlined) |
| hHEV_G4 ORF2-Bam aa606 Rev | GGATCC CTA CGCAGAGTGAGGTGCGAGGAC | BamH I site (underlined) |
| hHEV_G4 ORF2-Bam aa609 Rev | GGATCC CTA AGCGGCCAGCGCAGAGTGAG | BamH I site (underlined) |
| hHEV_G4 ORF2-Bam aa612 Rev | GGATCC CTA GTCCTCTAAAGCGGCCAGCG | BamH I site (underlined) |
| rbHEV_G3 T6686C Fw | TACGGCTCGTCAACTAACCC | nt 6684-6703 a (mutated nt underlined) |
| rbHEV_G3 T6686C Rev | TGTAGTTTGATCATACTCTG | nt 6664-6683 a |
| Purified VLPs | Amount (mL) Recovered after Gel Filtration | Concentration (Mean ± SD, mg/mL) a | Molecular Weight (Mean ± SD, kDa) b | Yield (Mean ± SD, mg) | Purity (Mean ± SD, %) b |
|---|---|---|---|---|---|
| HEV-3-VLP_89-606 (p518) | 48 | 0.5 ± 0.1 | 63.4 ± 0.7 | 24.0 ± 6.8 | 89.5 ± 2.8 |
| HEV-3-VLP_107-606 (p500) | 48 | 1.4 ± 0.1 | 61.1 ± 0.4 | 66.0 ± 6.0 | 90.7 ± 4.1 |
| HEV-3-VLP_112-606 (p495) | 48 | 1.9 ± 0.3 | 61.0 ± 0.5 | 89.6 ± 15.4 | 95.3 ± 1.4 |
| HEV-3-VLP_117-606 (p490) | 48 | 1.3 ± 0.1 | 60.3 ± 0.6 | 64.0 ± 5.5 | 91.9 ± 2.3 |
| HEV-3-VLP_122-606 (p485) | 48 | 1.6 ± 0.4 | 58.4 ± 1.1 | 76.8 ± 17.3 | 95.0 ± 1.6 |
| HEV-3-VLP_368-606 (p239) | 12 | 1.4 ± 0.6 | 27.1 ± 0.5 | 16.2 ± 7.6 | 93.1 ± 0.1 |
| HEV-3ra-VLP_112-606 (p495) | 16 | 0.9 ± 0.2 | 62.7 ± 0.6 | 13.9 ± 3.7 | 94.9 ± 1.9 |
| HEV-4-VLP_114-606 (p493) | 48 | 0.8 ± 0.1 | 61.9 ± 0.7 | 39.2 ± 5.0 | 74.7 ± 5.8 |
| HEV-4-VLP_117-606 (p490) | 48 | 1.3 ± 0.4 | 60.7 ± 0.3 | 62.4 ± 20.4 | 90.1 ± 8.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, T.; Takahashi, M.; Ohta, S.; Hoshino, Y.; Yamada, K.; Jirintai, S.; Primadharsini, P.P.; Nagashima, S.; Murata, K.; Okamoto, H. Production and Characterization of Self-Assembled Virus-like Particles Comprising Capsid Proteins from Genotypes 3 and 4 Hepatitis E Virus (HEV) and Rabbit HEV Expressed in Escherichia coli. Viruses 2024, 16, 1400. https://doi.org/10.3390/v16091400
Kobayashi T, Takahashi M, Ohta S, Hoshino Y, Yamada K, Jirintai S, Primadharsini PP, Nagashima S, Murata K, Okamoto H. Production and Characterization of Self-Assembled Virus-like Particles Comprising Capsid Proteins from Genotypes 3 and 4 Hepatitis E Virus (HEV) and Rabbit HEV Expressed in Escherichia coli. Viruses. 2024; 16(9):1400. https://doi.org/10.3390/v16091400
Chicago/Turabian StyleKobayashi, Tominari, Masaharu Takahashi, Satoshi Ohta, Yu Hoshino, Kentaro Yamada, Suljid Jirintai, Putu Prathiwi Primadharsini, Shigeo Nagashima, Kazumoto Murata, and Hiroaki Okamoto. 2024. "Production and Characterization of Self-Assembled Virus-like Particles Comprising Capsid Proteins from Genotypes 3 and 4 Hepatitis E Virus (HEV) and Rabbit HEV Expressed in Escherichia coli" Viruses 16, no. 9: 1400. https://doi.org/10.3390/v16091400
APA StyleKobayashi, T., Takahashi, M., Ohta, S., Hoshino, Y., Yamada, K., Jirintai, S., Primadharsini, P. P., Nagashima, S., Murata, K., & Okamoto, H. (2024). Production and Characterization of Self-Assembled Virus-like Particles Comprising Capsid Proteins from Genotypes 3 and 4 Hepatitis E Virus (HEV) and Rabbit HEV Expressed in Escherichia coli. Viruses, 16(9), 1400. https://doi.org/10.3390/v16091400