
Screening of Neutralizing Antibodies Targeting Gc Protein of RVFV

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Biosafety Information
2.2. Cells, Viruses, and Animals
2.3. Gc Protein Expression and Purification
2.4. Immunization of Alpaca
2.5. Construction of Single-Domain Antibody Library
2.6. Panning of the Gc-Specific Phage Display Nanobody Library
2.7. Screening of Gc-Specific Positive Clones
2.8. Cloning, Expression, and Purification of VHH-Fc
2.9. Enzyme-Linked Immunosorbent Assays (ELISAs)
2.10. Virus Neutralization Tests
2.11. KD Analysis via the Biacore System
2.12. Reduced SDS-PAGE
2.13. Engineering and Production of Bispecific Antibodies in IgG-VHH Format
2.14. RVFV Challenge Experiments
2.15. Histopathology and Immunohistochemistry
2.16. Quantitative RT-PCR
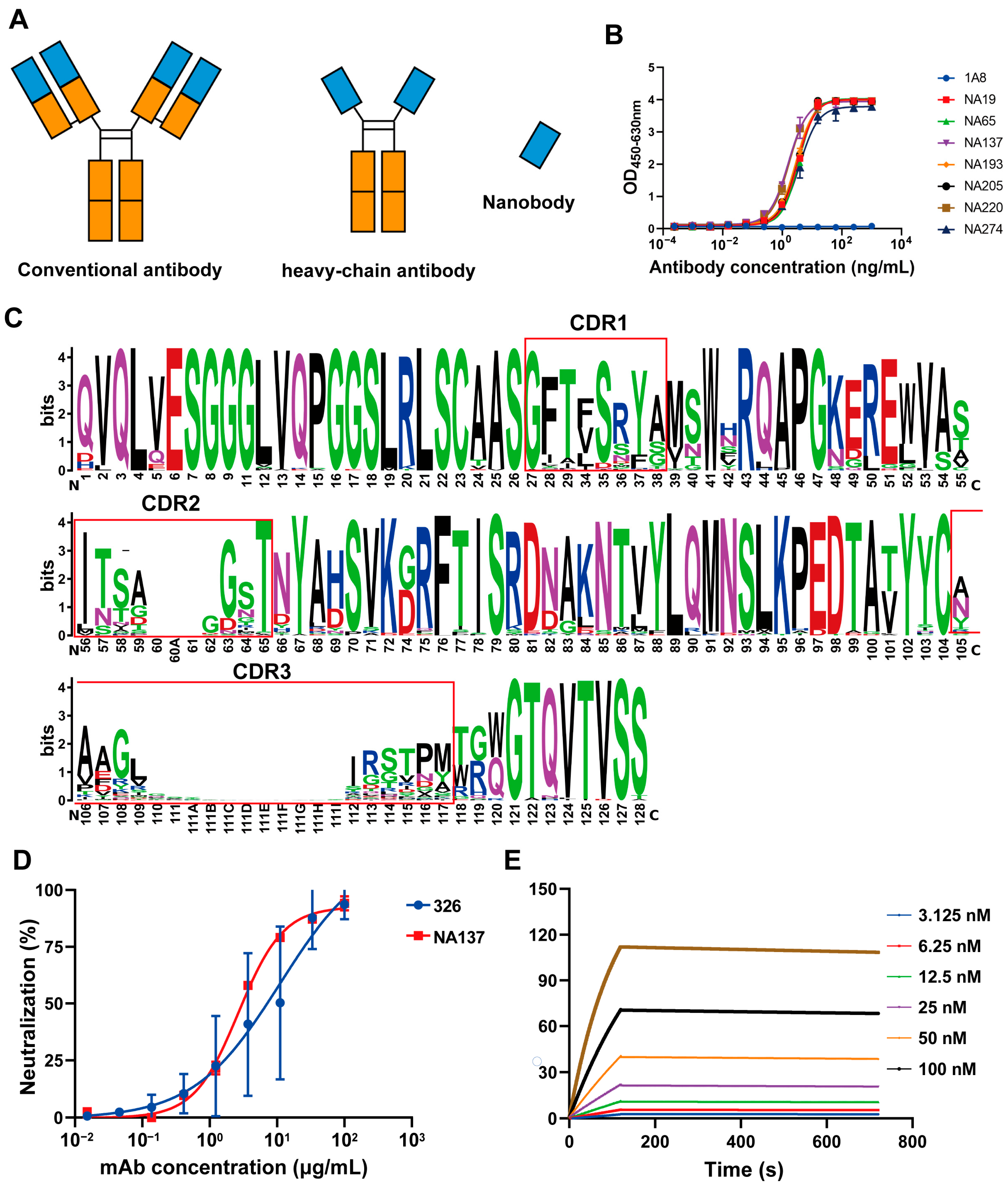
2.17. Analysis of Binding Antibody Sequences
2.18. Simulation of Structure of Antigen–Antibody Complexes Through Alphafold3
2.19. Statistical Analyses
3. Results
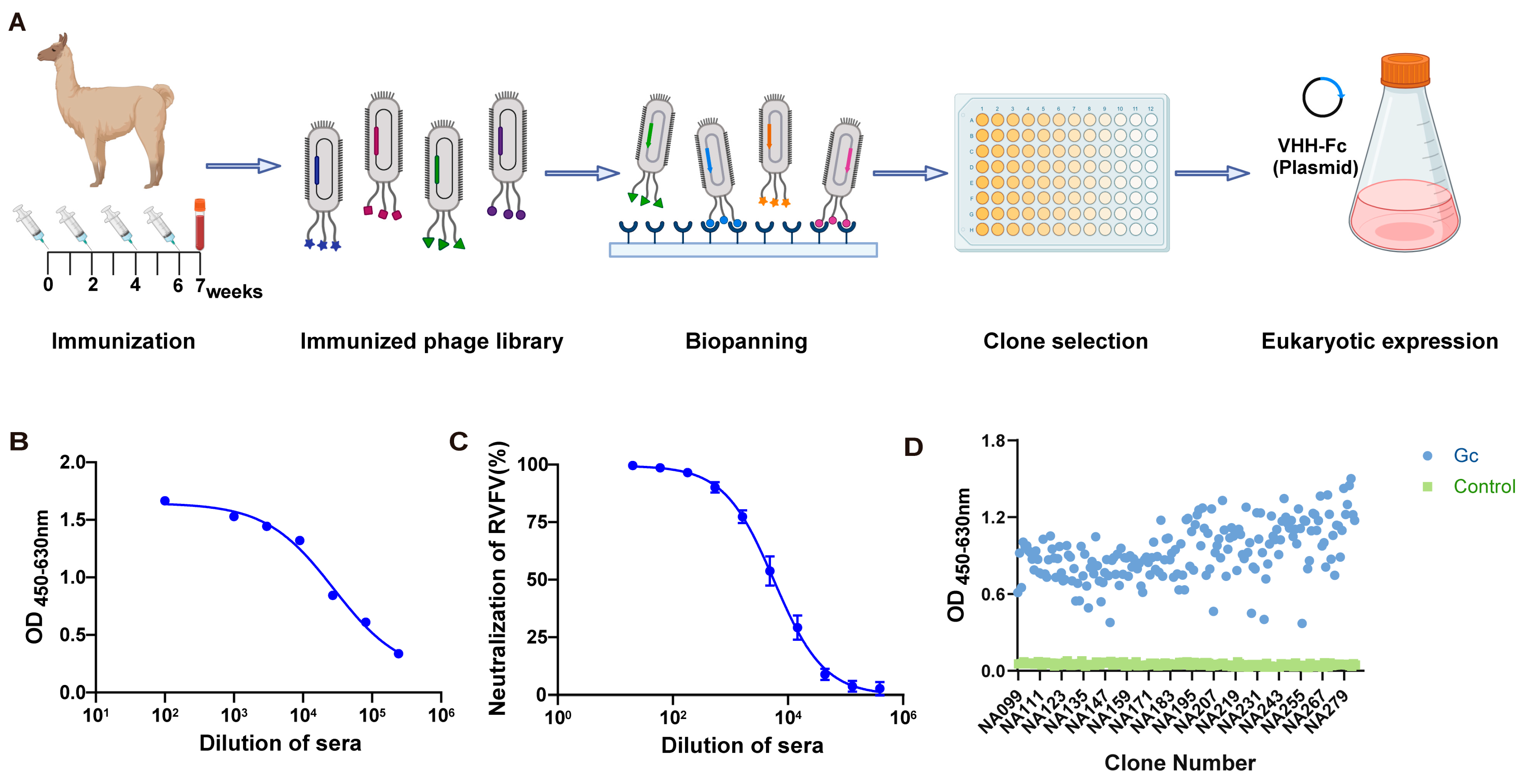
3.1. Construction and Panning of Gc-Specific Phage Display Nanobody Library
3.2. Generation and Screening for Gc-Specific Neutralizing Antibodies
3.3. Design and Enhanced Neutralizing Potency for IgG-VHH
3.4. NA137 and E2-NA137 Prevented Infection When Administered Prior to Challenge
3.5. NA137 and E2-NA137 Prevented Infection When Delivered Post-Challenge
3.6. AlphaFold 3 Analysis of Binding Epitopes of E2-NA137
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, D.; Kortekaas, J.; Bowden, T.A.; Warimwe, G.M. Rift Valley fever: Biology and epidemiology. J. Gen. Virol. 2019, 100, 1187–1199. [Google Scholar] [CrossRef]
- Bird, B.H.; Githinji, J.W.; Macharia, J.M.; Kasiiti, J.L.; Muriithi, R.M.; Gacheru, S.G.; Musaa, J.O.; Towner, J.S.; Reeder, S.A.; Oliver, J.B.; et al. Multiple virus lineages sharing recent common ancestry were associated with a Large Rift Valley fever outbreak among livestock in Kenya during 2006–2007. J. Virol. 2008, 82, 11152–11166. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Makino, S. The pathogenesis of Rift Valley fever. Viruses 2011, 3, 493–519. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lei, C.; Hu, D.; Dimitrov, D.S.; Ying, T. Human monoclonal antibodies as candidate therapeutics against emerging viruses. Front. Med. 2017, 11, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Murithi, R.M.; Munyua, P.; Ithondeka, P.M.; Macharia, J.M.; Hightower, A.; Luman, E.T.; Breiman, R.F.; Njenga, M.K. Rift Valley fever in Kenya: History of epizootics and identification of vulnerable districts. Epidemiol. Infect. 2011, 139, 372–380. [Google Scholar] [CrossRef]
- Madani, T.A.; Al-Mazrou, Y.Y.; Al-Jeffri, M.H.; Mishkhas, A.A.; Al-Rabeah, A.M.; Turkistani, A.M.; Al-Sayed, M.O.; Abodahish, A.A.; Khan, A.S.; Ksiazek, T.G.; et al. Rift Valley fever epidemic in Saudi Arabia: Epidemiological, clinical, and laboratory characteristics. Clin. Infect. Dis. 2003, 37, 1084–1092. [Google Scholar] [CrossRef]
- Samy, A.M.; Peterson, A.T.; Hall, M. Phylogeography of Rift Valley Fever Virus in Africa and the Arabian Peninsula. PLoS Negl. Trop. Dis. 2017, 11, e0005226. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.; Shi, W.; Tan, S.; Pan, Y.; Cui, S.; Zhang, Q.; Dou, X.; Lv, Y.; Li, X.; et al. The first imported case of Rift Valley fever in China reveals a genetic reassortment of different viral lineages. Emerg. Microbes Infect. 2017, 6, e4. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Banyard, A.C.; McElhinney, L.; Johnson, N.; Horton, D.L.; Hernandez-Triana, L.M.; Fooks, A.R. Rift Valley fever virus: A review of diagnosis and vaccination, and implications for emergence in Europe. Vaccine 2015, 33, 5520–5531. [Google Scholar] [CrossRef]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avsic-Zupanc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef]
- Maes, P.; Adkins, S.; Alkhovsky, S.V.; Avsic-Zupanc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; Briese, T.; et al. Taxonomy of the order Bunyavirales: Second update 2018. Arch. Virol. 2019, 164, 927–941. [Google Scholar] [CrossRef]
- Dessau, M.; Modis, Y. Crystal structure of glycoprotein C from Rift Valley fever virus. Proc. Natl. Acad. Sci. USA 2013, 110, 1696–1701. [Google Scholar] [CrossRef] [PubMed]
- Halldorsson, S.; Behrens, A.J.; Harlos, K.; Huiskonen, J.T.; Elliott, R.M.; Crispin, M.; Brennan, B.; Bowden, T.A. Structure of a phleboviral envelope glycoprotein reveals a consolidated model of membrane fusion. Proc. Natl. Acad. Sci. USA 2016, 113, 7154–7159. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, Y.; Chai, Y.; Qi, J.; Peng, R.; Feng, W.H.; Gao, G.F. The Postfusion Structure of the Heartland Virus Gc Glycoprotein Supports Taxonomic Separation of the Bunyaviral Families Phenuiviridae and Hantaviridae. J. Virol. 2018, 92, e01558-17. [Google Scholar] [CrossRef]
- Guardado-Calvo, P.; Atkovska, K.; Jeffers, S.A. A glycerophospholipid-specific pocket in the RVFV class II fusion protein drives target membrane insertion. Science 2017, 358, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Fels, J.M.; Maurer, D.P.; Herbert, A.S.; Wirchnianski, A.S.; Vergnolle, O.; Cross, R.W.; Abelson, D.M.; Moyer, C.L.; Mishra, A.K.; Aguilan, J.T.; et al. Protective neutralizing antibodies from human survivors of Crimean-Congo hemorrhagic fever. Cell 2021, 184, 3486–3501.e21. [Google Scholar] [CrossRef] [PubMed]
- Halldorsson, S.; Li, S.; Li, M.; Harlos, K.; Bowden, T.A.; Huiskonen, J.T. Shielding and activation of a viral membrane fusion protein. Nat. Commun. 2018, 9, 349. [Google Scholar] [CrossRef]
- Faburay, B.; LaBeaud, A.D.; McVey, D.S.; Wilson, W.C.; Richt, J.A. Current Status of Rift Valley Fever Vaccine Development. Vaccines 2017, 5, 29. [Google Scholar] [CrossRef]
- Müller, R.; Poch, O.; Delarue, M.; Bishop, D.H.L.; Bouloy, M. Rift valley fever virus L segment: Correction of the sequence and possible functional role of newly identified regions conserved in RNA-dependent polymerases. J. Gen. Virol. 1994, 75, 1345–1352. [Google Scholar] [CrossRef]
- de Boer, S.M.; Kortekaas, J.; Spel, L.; Rottier, P.J.; Moormann, R.J.; Bosch, B.J. Acid-activated structural reorganization of the Rift Valley fever virus Gc fusion protein. J. Virol. 2012, 86, 13642–13652. [Google Scholar] [CrossRef]
- Freiberg, A.N.; Sherman, M.B.; Morais, M.C.; Holbrook, M.R.; Watowich, S.J. Three-dimensional organization of Rift Valley fever virus revealed by cryoelectron tomography. J. Virol. 2008, 82, 10341–10348. [Google Scholar] [CrossRef] [PubMed]
- Huiskonen, J.T.; Overby, A.K.; Weber, F.; Grunewald, K. Electron cryo-microscopy and single-particle averaging of Rift Valley fever virus: Evidence for GN-GC glycoprotein heterodimers. J. Virol. 2009, 83, 3762–3769. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhu, Y.; Gao, F.; Jiao, Y.; Oladejo, B.O.; Chai, Y.; Bi, Y.; Lu, S.; Dong, M.; Zhang, C.; et al. Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope. Proc. Natl. Acad. Sci. USA 2017, 114, E7564–E7573. [Google Scholar] [CrossRef] [PubMed]
- Besselaar, T.G.; Blackburn, N.K.; Meenehan, G.M. Antigenic analysis of Pdft Valley fever virus isolates: Monoclona|antibodies distinguish between wild-type and neurotropic virus strains. Res. Virol. 1991, 142, 469–474. [Google Scholar] [CrossRef]
- Besselaar, T.G.; Blackburn, N.K. The effect of neutralizing monoclonai antibodies on early events in Rift Valley fever virus infectivity. Res. Virol. 1994, 145, 13–19. [Google Scholar] [CrossRef]
- Besselaar, T.G.; Blackburn, N.K. The synergistic neutralization of Rift Valley fever virus by monoclonal antibodies to the envelope glycoproteins. Arch. Virol. 1992, 125, 239–250. [Google Scholar] [CrossRef]
- Kitandwe, P.K.; McKay, P.F.; Kaleebu, P.; Shattock, R.J. An Overview of Rift Valley Fever Vaccine Development Strategies. Vaccines 2022, 10, 1794. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, T.; Wu, Y.; Chen, Z.; Zeng, H.; Tong, Z.; Gao, F.; Qi, J.; Zhao, Z.; Chai, Y.; et al. Neutralization mechanism of human monoclonal antibodies against Rift Valley fever virus. Nat. Microbiol. 2019, 4, 1231–1241. [Google Scholar] [CrossRef]
- Chapman, N.S.; Zhao, H.; Kose, N.; Westover, J.B.; Kalveram, B.; Bombardi, R.; Rodriguez, J.; Sutton, R.; Genualdi, J.; LaBeaud, A.D.; et al. Potent neutralization of Rift Valley fever virus by human monoclonal antibodies through fusion inhibition. Proc. Natl. Acad. Sci. USA 2021, 118, e2025642118. [Google Scholar] [CrossRef]
- Eden, T.; Menzel, S.; Wesolowski, J.; Bergmann, P.; Nissen, M.; Dubberke, G.; Seyfried, F.; Albrecht, B.; Haag, F.; Koch-Nolte, F. A cDNA Immunization Strategy to Generate Nanobodies against Membrane Proteins in Native Conformation. Front. Immunol. 2017, 8, 1989. [Google Scholar] [CrossRef]
- Wesolowski, J.; Alzogaray, V.; Reyelt, J.; Unger, M.; Juarez, K.; Urrutia, M.; Cauerhff, A.; Danquah, W.; Rissiek, B.; Scheuplein, F.; et al. Single domain antibodies: Promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009, 198, 157–174. [Google Scholar] [CrossRef]
- Muyldermans, S. Nanobodies: Natural single-domain antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [PubMed]
- De Genst, E.; Silence, K.; Decanniere, K.; Conrath, K.; Loris, R.; Kinne, J.; Muyldermans, S.; Wyns, L. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. Proc. Natl. Acad. Sci. USA 2006, 103, 4586–4591. [Google Scholar] [CrossRef]
- Arbabi-Ghahroudi, M. Camelid Single-Domain Antibodies: Historical Perspective and Future Outlook. Front. Immunol. 2017, 8, 1589. [Google Scholar] [CrossRef] [PubMed]
- Farrington, G.K.; Caram-Salas, N.; Haqqani, A.S.; Brunette, E.; Eldredge, J.; Pepinsky, B.; Antognetti, G.; Baumann, E.; Ding, W.; Garber, E.; et al. A novel platform for engineering blood-brain barrier-crossing bispecific biologics. FASEB J. 2014, 28, 4764–4778. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Qi, S.; Unger, M.; Hou, Y.N.; Deng, Q.W.; Liu, J.; Lam, C.M.C.; Wang, X.W.; Xin, D.; Zhang, P.; et al. Immuno-targeting the multifunctional CD38 using nanobody. Sci. Rep. 2016, 6, 27055. [Google Scholar] [CrossRef]
- Herce, H.D.; Schumacher, D.; Schneider, A.F.L.; Ludwig, A.K.; Mann, F.A.; Fillies, M.; Kasper, M.A.; Reinke, S.; Krause, E.; Leonhardt, H.; et al. Cell-permeable nanobodies for targeted immunolabelling and antigen manipulation in living cells. Nat. Chem. 2017, 9, 762–771. [Google Scholar] [CrossRef]
- Bouchet, J.; Basmaciogullari, S.E.; Chrobak, P.; Stolp, B.; Bouchard, N.; Fackler, O.T.; Chames, P.; Jolicoeur, P.; Benichou, S.; Baty, D. Inhibition of the Nef regulatory protein of HIV-1 by a single-domain antibody. Blood 2011, 117, 3559–3568. [Google Scholar] [CrossRef]
- Deng, Q.; Guo, Z.; Hu, H.; Li, Q.; Zhang, Y.; Wang, J.; Liao, C.; Guo, C.; Li, X.; Chen, Z.; et al. Inhibition of Chikungunya virus early replication by intracellular nanoantibodies targeting nsP2 Epitope Rich Region. Antivir. Res. 2022, 208, 105446. [Google Scholar] [CrossRef]
- Zhao, G.; He, L.; Sun, S.; Qiu, H.; Tai, W.; Chen, J.; Li, J.; Chen, Y.; Guo, Y.; Wang, Y.; et al. A Novel Nanobody Targeting Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Receptor-Binding Domain Has Potent Cross-Neutralizing Activity and Protective Efficacy against MERS-CoV. J. Virol. 2018, 92, e00837-18. [Google Scholar] [CrossRef]
- Wu, Y.; Li, C.; Xia, S.; Tian, X.; Kong, Y.; Wang, Z.; Gu, C.; Zhang, R.; Tu, C.; Xie, Y.; et al. Identification of Human Single-Domain Antibodies against SARS-CoV-2. Cell Host Microbe 2020, 27, 891–898.e5. [Google Scholar] [CrossRef] [PubMed]
- Marie Scully, M.D. Caplacizumab Treatment for Acquired Thrombotic Thrombocytopenic Purpura. N. Engl. J. Med. 2019, 380, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.J. Ozoralizumab: First Approval. Drugs 2023, 83, 87–92. [Google Scholar] [CrossRef]
- Labrijn, A.F.; Janmaat, M.L.; Reichert, J.M.; Parren, P. Bispecific antibodies: A mechanistic review of the pipeline. Nat. Rev. Drug Discov. 2019, 18, 585–608. [Google Scholar] [CrossRef]
- Nyakatura, E.K.; Soare, A.Y.; Lai, J.R. Bispecific antibodies for viral immunotherapy. Hum. Vaccin. Immunother. 2017, 13, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, J.L.; Ferrari, G.; Margolis, D.M. Bispecific antibody-derived molecules to target persistent HIV infection. J. Virus Erad. 2022, 8, 100083. [Google Scholar] [CrossRef]
- Frei, J.C.; Nyakatura, E.K.; Zak, S.E.; Bakken, R.R.; Chandran, K.; Dye, J.M.; Lai, J.R. Bispecific Antibody Affords Complete Post-Exposure Protection of Mice from Both Ebola (Zaire) and Sudan Viruses. Sci. Rep. 2016, 6, 19193. [Google Scholar] [CrossRef]
- Wang, R.; Lu, J.; Chen, L.; Yu, Y.; Yang, Z. A human bispecific neutralization antibody against four serotypes of dengue virus. Virology 2021, 558, 49–56. [Google Scholar] [CrossRef]
- Sun, H.; Xia, L.; Li, J.; Zhang, Y.; Zhang, G.; Huang, P.; Wang, X.; Cui, Y.; Fang, T.; Fan, P.; et al. A novel bispecific antibody targeting two overlapping epitopes in RBD improves neutralizing potency and breadth against SARS-CoV-2. Emerg. Microbes Infect. 2024, 13, 2373307. [Google Scholar] [CrossRef]
- Chi, H.; Wang, L.; Liu, C.; Cheng, X.; Zheng, H.; Lv, L.; Tan, Y.; Zhang, N.; Zhao, S.; Wu, M.; et al. An Engineered IgG-VHH Bispecific Antibody against SARS-CoV-2 and Its Variants. Small Methods 2022, 6, e2200932. [Google Scholar] [CrossRef]
- Cho, H.; Gonzales-Wartz, K.K.; Huang, D. Bispecific antibodies targeting distinct regions of the spike protein potently neutralize SARS-CoV-2 variants of concern. Sci. Transl. Med. 2021, 13, eabj5413. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Ma, Y.; Wang, Y.; Zhan, W.; Zheng, Q.; Zhang, M.; Ji, P.; Liu, M.; Liu, Q.; et al. Combating the SARS-CoV-2 Omicron (BA.1) and BA.2 with potent bispecific antibodies engineered from non-Omicron neutralizing antibodies. Cell Discov. 2022, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Won, S.; Peters, C.J.; Makino, S. Rescue of infectious rift valley fever virus entirely from cDNA, analysis of virus lacking the NSs gene, and expression of a foreign gene. J. Virol. 2006, 80, 2933–2940. [Google Scholar] [CrossRef]
- Bian, T.; Hao, M.; Zhao, X.; Zhao, C.; Luo, G.; Zhang, Z.; Fu, G.; Yang, L.; Chen, Y.; Wang, Y.; et al. A Rift Valley fever mRNA vaccine elicits strong immune responses in mice and rhesus macaques. NPJ Vaccines 2023, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Wichgers Schreur, P.J.; Paweska, J.T.; Kant, J.; Kortekaas, J. A novel highly sensitive, rapid and safe Rift Valley fever virus neutralization test. J. Virol. Methods 2017, 248, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Bouhaddou, M.; Memon, D.; Meyer, B.; White, K.M.; Rezelj, V.V.; Correa Marrero, M.; Polacco, B.J.; Melnyk, J.E.; Ulferts, S.; Kaake, R.M.; et al. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 2020, 182, 685–712.e19. [Google Scholar]
- Wichgers Schreur, P.J.; van de Water, S.; Harmsen, M.; Bermudez-Mendez, E.; Drabek, D.; Grosveld, F.; Wernike, K.; Beer, M.; Aebischer, A.; Daramola, O.; et al. Multimeric single-domain antibody complexes protect against bunyavirus infections. eLife 2020, 9, e52716. [Google Scholar] [CrossRef]
- Bird, B.H.; Bawiec, D.A.; Ksiazek, T.G.; Shoemaker, T.R.; Nichol, S.T. Highly sensitive and broadly reactive quantitative reverse transcription-PCR assay for high-throughput detection of Rift Valley fever virus. J. Clin. Microbiol. 2007, 45, 3506–3513. [Google Scholar] [CrossRef]
- Reed, C.; Lin, K.; Wilhelmsen, C.; Friedrich, B.; Nalca, A.; Keeney, A.; Donnelly, G.; Shamblin, J.; Hensley, L.E.; Olinger, G.; et al. Aerosol exposure to Rift Valley fever virus causes earlier and more severe neuropathology in the murine model, which has important implications for therapeutic development. PLoS Negl. Trop. Dis. 2013, 7, e2156. [Google Scholar] [CrossRef]
- Gowen, B.B.; Wong, M.H.; Jung, K.H.; Sanders, A.B.; Mendenhall, M.; Bailey, K.W.; Furuta, Y.; Sidwell, R.W. In vitro and in vivo activities of T-705 against arenavirus and bunyavirus infections. Antimicrob. Agents Chemother. 2007, 51, 3168–3176. [Google Scholar] [CrossRef]
- Caroline, A.L.; Powell, D.S.; Bethel, L.M.; Oury, T.D.; Reed, D.S.; Hartman, A.L. Broad spectrum antiviral activity of favipiravir (T-705): Protection from highly lethal inhalational Rift Valley Fever. PLoS Negl. Trop. Dis. 2014, 8, e2790. [Google Scholar] [CrossRef] [PubMed]
- Scharton, D.; Bailey, K.W.; Vest, Z.; Westover, J.B.; Kumaki, Y.; Van Wettere, A.; Furuta, Y.; Gowen, B.B. Favipiravir (T-705) protects against peracute Rift Valley fever virus infection and reduces delayed-onset neurologic disease observed with ribavirin treatment. Antivir. Res. 2014, 104, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zhang, G.; Zhang, S.; Chen, Z.; Chi, X.; Dong, Y.; Fan, P.; Liu, Y.; Chen, Y.; Song, X.; et al. Characterization of Two Neutralizing Antibodies against Rift Valley Fever Virus Gn Protein. Viruses 2020, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.R.; Krumm, S.A.; Raghwani, J.; Halldorsson, S.; Elliott, A.; Graham, V.A.; Koudriakova, E.; Harlos, K.; Wright, D.; Warimwe, G.M.; et al. A Protective Monoclonal Antibody Targets a Site of Vulnerability on the Surface of Rift Valley Fever Virus. Cell Rep. 2018, 25, 3750–3758.e4. [Google Scholar] [CrossRef]
- Gutjahr, B.; Keller, M.; Rissmann, M.; von Arnim, F.; Jackel, S.; Reiche, S.; Ulrich, R.; Groschup, M.H.; Eiden, M. Two monoclonal antibodies against glycoprotein Gn protect mice from Rift Valley Fever challenge by cooperative effects. PLoS Negl. Trop. Dis. 2020, 14, e0008143. [Google Scholar] [CrossRef]
- Cartwright, H.N.; Barbeau, D.J.; McElroy, A.K. Isotype-Specific Fc Effector Functions Enhance Antibody-Mediated Rift Valley Fever Virus Protection In Vivo. mSphere 2021, 6, e0055621. [Google Scholar] [CrossRef]
- Bird, B.H.; Khristova, M.L.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Complete genome analysis of 33 ecologically and biologically diverse Rift Valley fever virus strains reveals widespread virus movement and low genetic diversity due to recent common ancestry. J. Virol. 2007, 81, 2805–2816. [Google Scholar] [CrossRef]
- Lozach, P.Y.; Kuhbacher, A.; Meier, R.; Mancini, R.; Bitto, D.; Bouloy, M.; Helenius, A. DC-SIGN as a receptor for phleboviruses. Cell Host Microbe 2011, 10, 75–88. [Google Scholar] [CrossRef]
- Sherman, M.B.; Freiberg, A.N.; Holbrook, M.R.; Watowich, S.J. Single-particle cryo-electron microscopy of Rift Valley fever virus. Virology 2009, 387, 11–15. [Google Scholar] [CrossRef]
- Chang, Z.; Gao, D.; Liao, L.; Sun, J.; Zhang, G.; Zhang, X.; Wang, F.; Li, C.; Oladejo, B.O.; Li, S.; et al. Bispecific antibodies targeting two glycoproteins on SFTSV exhibit synergistic neutralization and protection in a mouse model. Proc. Natl. Acad. Sci. USA 2024, 121, e2400163121. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Target | Source | In Vitro Efficacy (IC50) | In Vivo Efficacy | Ref. |
|---|---|---|---|---|---|
| RV-Gn1 | Gn | Rabbit | 3000 ng/mL (Vero) | BALB/c mice (100%) | [64] |
| RV-Gn2 | Gn | Rabbit | 2800 ng/mL (Vero) | BALB/c mice (100%) | [64] |
| RV-Gn3 | Gn | Rabbit | 2100 ng/mL (Vero) | BALB/c mice (100%) | [64] |
| Gn3 | Gn | Mouse | 33,000 ng/mL (Vero 76) | BALB/c mice (58.3%) | [65] |
| Gn3 + Gn32 | Gn | Mouse | 24,600 ng/mL (Vero 76) | BALB/c mice (100%) | [65] |
| 268 | Gn | Human | 0.1 ng/mL (Vero) | C57BL/6 mice (90%) | [29] |
| 142 | Gn | Human | 5.9 ng/mL (Vero) | [29] | |
| 436 | Gn | Human | 2.5 ng/mL (Vero) | [29] | |
| 429 | Gn | Human | 0.9 ng/mL (Vero) | [29] | |
| 379 | Gn | Human | 1.3 ng/mL (Vero) | [29] | |
| 140 | * | Human | 1.1 ng/mL (Vero) | C57BL/6 mice (100%) | [29] |
| 426 | Gn | Human | 9.1 ng/mL (Vero) | [29] | |
| 296 | Gn | Human | 638 ng/mL (Vero) | [29] | |
| 220 | * | Human | 157 ng/mL (Vero) | [29] | |
| 250 | Gc | Human | 51 ng/mL (Vero) | [29] | |
| 401 | Gn | Human | 2120 ng/mL (Vero) | [29] | |
| 128 | Gc | Human | 38 ng/mL (Vero) | [29] | |
| 121 | Gc | Human | 35 ng/mL (Vero) | [29] | |
| 326 | Gc | Human | 70 ng/mL (Vero) | [29] | |
| 144 | * | Human | 1.9 ng/mL (Vero) | [29] | |
| 381 | Gn | Human | 4435 ng/mL (Vero) | [29] | |
| 226 | Gn | Human | 7692 ng/mL (Vero) | C57BL/6 mice (50%) | [29] |
| 229 | NT ** | Human | 3040 ng/mL (Vero) | [29] | |
| 405 | Gn | Human | >(Vero) | [29] | |
| 249 | Gc | Human | 6687 (Vero) | [29] | |
| mAb 1 | Gn | Mouse | 28 ng/mL (Vero-E6) | C57BL/6 mice (100%) | [66] |
| mAb 2 | Gn | Mouse | 1532 ng/mL (Vero-E6) | C57BL/6 mice (100%) | [66] |
| mAb 3 | Gn | Mouse | >(Vero-E6) | C57BL/6 mice (100%) | [66] |
| mAb 4 | Gn | Mouse | >(Vero-E6) | [66] | |
| mAb 5 | Gn | Mouse | >(Vero-E6) | [66] | |
| mAb 6 | Gn | Mouse | >(Vero-E6) | [66] | |
| RVFV-VHHs | Gn | Alpaca | (Vero-E6) | BALB/c mice (20%) | [57] |
| 1332F11 | Gn | Rhesus monkey | 170 ng/mL (Huh7) | [63] | |
| 1331E4 | Gn | Rhesus monkey | 340 ng/mL (Huh7) | [63] | |
| R4 | Gn | Human | 46.8 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R5 | Gc | Human | 1370 ng/mL (Vero) | BALB/c mice (20%) | [28] |
| R12 | Gn | Human | 1.85 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R13 | Gn | Human | 56.2 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R15 | Gn | Human | 0.53 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R16 | Gn | Human | 0.29 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R17 | Gn | Human | 2.53 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R19 | Gn | Human | 51.1 ng/mL (Vero) | BALB/c mice (100%) | [28] |
| R22 | Gn | Human | 73.7 ng/mL (Vero) | BALB/c mice (100%) | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Hao, M.; Bian, T.; Zhao, X.; Chi, X.; Chen, Z.; Fu, G.; Zhu, Z.; Fang, T.; Yu, C.; et al. Screening of Neutralizing Antibodies Targeting Gc Protein of RVFV. Viruses 2025, 17, 559. https://doi.org/10.3390/v17040559
Zhao C, Hao M, Bian T, Zhao X, Chi X, Chen Z, Fu G, Zhu Z, Fang T, Yu C, et al. Screening of Neutralizing Antibodies Targeting Gc Protein of RVFV. Viruses. 2025; 17(4):559. https://doi.org/10.3390/v17040559
Chicago/Turabian StyleZhao, Chuanyi, Meng Hao, Ting Bian, Xiaofan Zhao, Xiangyang Chi, Zhengshan Chen, Guangcheng Fu, Zheng Zhu, Ting Fang, Changming Yu, and et al. 2025. "Screening of Neutralizing Antibodies Targeting Gc Protein of RVFV" Viruses 17, no. 4: 559. https://doi.org/10.3390/v17040559
APA StyleZhao, C., Hao, M., Bian, T., Zhao, X., Chi, X., Chen, Z., Fu, G., Zhu, Z., Fang, T., Yu, C., Li, J., & Chen, W. (2025). Screening of Neutralizing Antibodies Targeting Gc Protein of RVFV. Viruses, 17(4), 559. https://doi.org/10.3390/v17040559