Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Geographical Distribution, Host Range and Epidemics
2. Genome Structure and Protein Functions
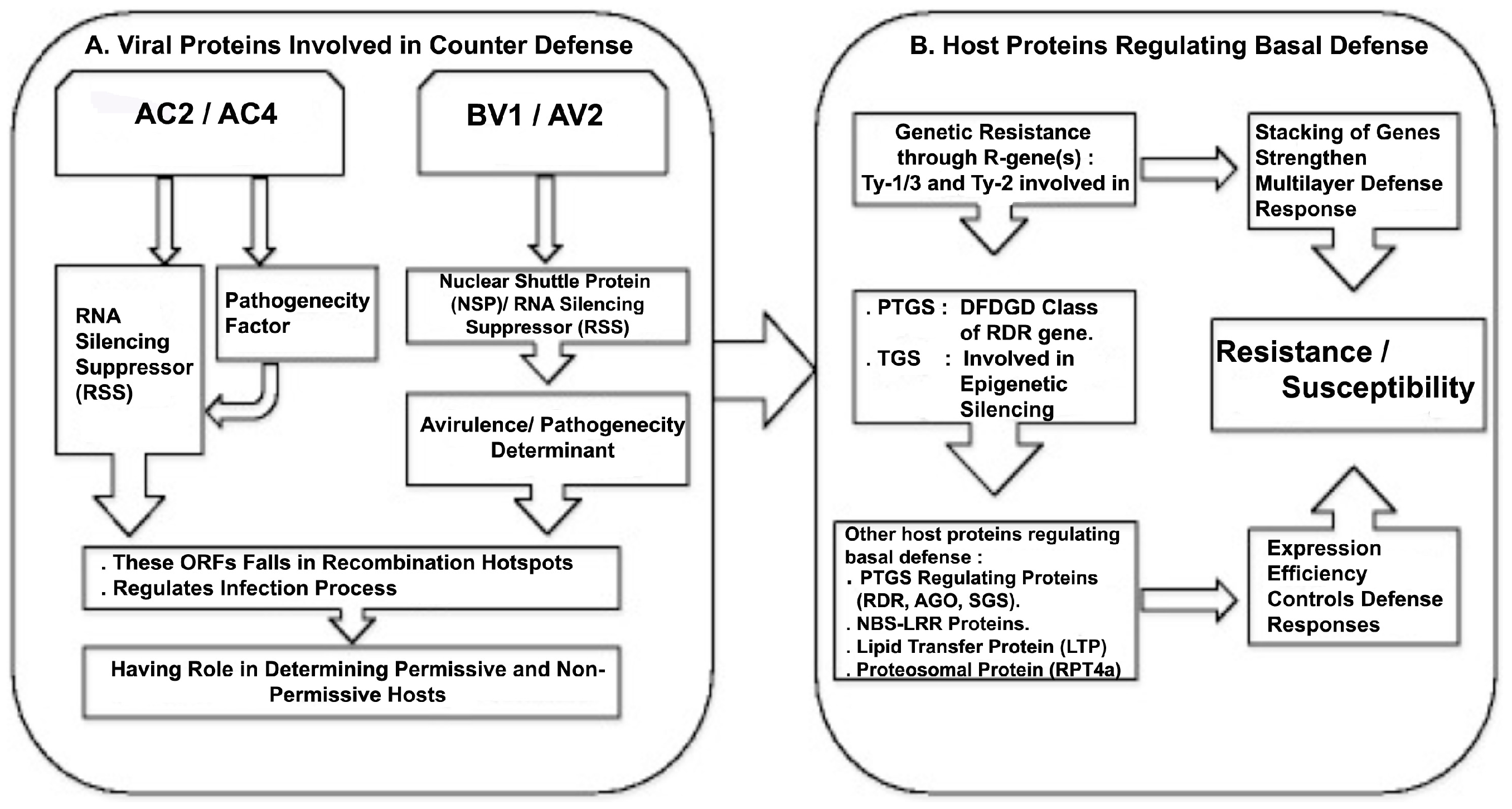
2.1. Viral Genes and Proteins
2.2. Suppression of Plant Defenses
3. Genetic Diversity, Host Adaptation and Evolution
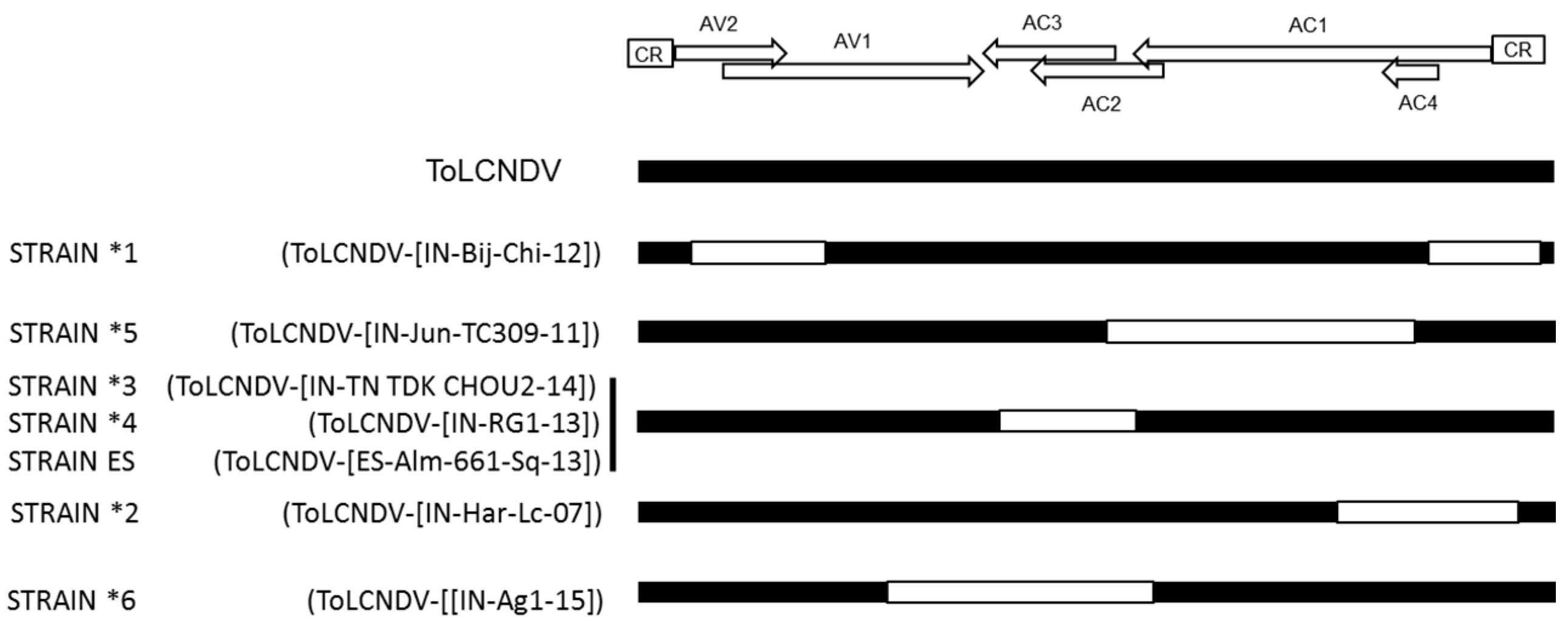
3.1. Variants of ToLCNDV
3.2. Host Adaption
3.3. Recombination and Pseudo-Recombination as Motors of Diversification
4. Biology and Interaction of Satellite Molecules with ToLCNDV
4.1. Disease Severity
4.2. Extending Host Range
5. Virus–Vector Interactions
6. Control Strategies to Reduce Disease Damage
6.1. Management Practices
6.2. Host Plant Resistance
6.3. Genetic Engineering Approaches
7. Concluding Remarks
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brown, J.K.; Fauquet, C.M.; Briddon, R.W.; Zerbini, F.M.; Moriones, E.; Navas-Castillo, J. Family Geminiviridae. In Virus Taxonomy 9th Report, Proceedings of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: London, UK, 2012; pp. 351–373. [Google Scholar]
- Varma, A.; Malathi, V.G. Emerging geminivirus problems: A serious threat to crop production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Zaidi, S.S.S.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato leaf curl New Delhi virus: A widespread bipartite begomovirus in the territory of monopartite begomoviruses. Mol. Plant Pathol. 2016, 18, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S. Tomato leaf curl viruses from India (Geminiviridae). In Enciclopedia of Virology; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Elsevier: London, UK, 2008; pp. 124–133. [Google Scholar]
- Mansoor, S.; Khan, S.H.; Saeed, M.; Bashir, A.; Zafar, Y.; Malik, K.A. Evidence for the association of a bipartite geminivirus with tomato leaf curl disease in Pakistan. Plant Dis. 1997, 81, 958. [Google Scholar] [CrossRef]
- Yazdani-Khameneh, S.; Aboutorabi, S.; Shoori, M.; Aghazadeh, A.; Jahanshahi, P.; Golnaraghi, A.; Maleki, M. Natural occurrence of tomato leaf curl New Delhi virus in Iranian cucurbit crops. Plant Pathol. J. 2016, 32, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Fortes, I.M.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, A.J.; Navas-Castillo, J.; Moriones, E. A novel strain of tomato leaf curl New Delhi virus has spread to the mediterranean basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.; Doña, D.B.; Ben Nahia, E.; Hajlaoui, M. First report of tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33, 6. [Google Scholar] [CrossRef]
- Ito, T.; Sharma, P.; Kittipakorn, K.; Ikegami, M. Complete nucleotide sequence of a new isolate of tomato leaf curl New Delhi virus infecting cucumber, bottle gourd and muskmelon in Thailand. Arch. Virol. 2008, 153, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.V.; Singh, A.K.; Singh, A.K.; Yadav, T.; Basu, S.; Kushwaha, N.; Chattopadhyay, B.; Chakraborty, S. Complexity of begomovirus and betasatellite populations associated with chilli leaf curl disease in India. J. Gen. Virol. 2015, 96, 3157–3172. [Google Scholar] [CrossRef] [PubMed]
- Pratap, D.; Kashikar, A.; Mukherjee, S. Molecular characterization and infectivity of a tomato leaf curl New Delhi virus variant associated with newly emerging yellow mosaic disease of eggplant in India. Virol. J. 2011, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, P.N.; Sumiya, K.V.; Malathi, V.G. Carrot as a new host for a begomovirus: Yellow mosaic disease of carrot reported in India. New Dis. Rep. 2011, 23, 34. [Google Scholar] [CrossRef]
- Tahir, M.; Haider, M.S. First report of tomato leaf curl New Delhi virus infecting bitter gourd in Pakistan. Plant Pathol. 2005, 54, 807. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Lakshminarayana, R.C.; Jalali, S.; Krishna, R.M. Association of tomato leaf curl New Delhi virus DNA-B with bhendi yellow vein mosaic virus in okra showing yellow vein mosaic disease symptoms. Acta Virol. 2014, 59, 125–139. [Google Scholar] [CrossRef]
- Roy, A.; Spoorthi, P.; Panwar, G.; Bag, M.K.; Prasad, T.; Kumar, G.; Gangopadhyay, K.; Dutta, M. Molecular evidence for occurrence of tomato leaf curl New Delhi virus in ash gourd (Benincasa hispida) germplasm showing a severe yellow stunt disease in India. Ind. J. Virol. 2013, 24, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.L.; Tsai, W.S.; Lee, L.M.; Kenyon, L. Molecular characterization of begomoviruses infecting Sauropus androgynus in Thailand. J. Phytopathol. 2013, 161, 78–85. [Google Scholar] [CrossRef]
- Zaidi, S.S.A.; Shafiq, M.; Amin, I.; Scheffler, B.E.; Scheffler, J.A.; Briddon, R.W.; Mansoor, S. Frequent occurrence of tomato leaf curl New Delhi virus in cotton leaf curl disease affected cotton in Pakistan. PLoS ONE 2016, 11, e0155520. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, R.; Heydarnejad, J.; Massumi, H.; Shaabanian, M.; Varsani, A. Genetic diversity and distribution of tomato-infecting begomoviruses in Iran. Virus Genes 2009, 38, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Mansoor, S.; Iram, S.; Zafar, Y.; Briddon, R.W. First report of tomato leaf curl New Delhi virus affecting chilli pepper in Pakistan. Plant Pathol. 2004, 53, 794. [Google Scholar] [CrossRef]
- Hameed, A.; Tahir, M.N.; Amin, I.; Mansoor, S. First report of tomato leaf curl New Delhi virus and a tomato yellow leaf curl Thailand betasatellite causing severe leaf curl disease of potato in Pakistan. Plant Dis. 2017, 101, 1065. [Google Scholar] [CrossRef]
- Usharani, K.S.; Surendranath, B.; Paul-Khurana, S.M.; Garg, I.D.; Malathi, V.G. Potato leaf curl—A new disease of potato in northern India caused by a strain of tomato leaf curl New Delhi virus. Plant Pathol. 2004, 53, 235. [Google Scholar] [CrossRef]
- Singh, A.K.; Mishra, K.K.; Chattopadhyay, B.; Chakraborty, S. Biological and molecular characterization of a begomovirus associated with yellow mosaic vein mosaic disease of pumpkin from Northern India. Virus Genes 2009, 39, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Akhter, A.; Akhtar, S.; Saeed, M.; Mansoor, S. Chili leaf curl betasatellite enhances symptoms induced by tomato leaf curl New Delhi virus, a bipartite begomovirus. Int. J. Agric. Biol. 2014, 16, 1225–1228. [Google Scholar]
- Jyothsna, P.; Haq, Q.M.I.; Singh, P.; Sumiya, K.V.; Praveen, S.; Rawat, R.; Briddon, R.W.; Malathi, V.G. Infection of tomato leaf curl New Delhi virus (ToLCNDV), a bipartite begomovirus with betasatellites, results in enhanced level of helper virus components and antagonistic interaction between DNA B and betasatellites. Appl. Microbiol. Biotechnol. 2013, 97, 5457–5471. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Li, G.; Wang, D.; Hu, D.; Zhou, X. A begomovirus DNA β-encoded protein binds DNA, functions as a suppressor of RNA silencing, and targets the cell nucleus. J. Virol. 2005, 79, 10764–10775. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. MicroRNA-binding viral protein interferes with Arabidopsis development. Proc. Natl. Acad. Sci. USA. 2005, 2, 10381–10386. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Phaneendra, C.; Rao, K.R.S.S.; Jain, R.K.; Mandal, B. Tomato leaf curl New Delhi virus is associated with pumpkin leaf curl: A new disease in Northern India. Ind. J. Virol. 2012, 23, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.V.; Choudhury, N.R.; Mukherjee, S.K. Transgenic tomato plants expressing artificial microRNAs for silencing the pre-coat and coat proteins of a begomovirus, tomato leaf curl New Delhi virus, show tolerance to virus infection. Virus Res. 2013, 172, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Mansoor, S.; Iram, S.; Fatima, A.N.; Mansoor, Y.Z. The nuclear shuttle protein of tomato leaf curl New Delhi virus is a pathogenicity determinant. J. Virol. 2005, 79, 4434–4439. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, H.C.; Sinha, D.P.; Rai, G.K.; Krishna, R.; Kashyap, S.P.; Singh, N.K.; Singh, M.; Malathi, V.G. Pyramiding Ty-2 and Ty-3 genes for resistance to monopartite and bipartite tomato leaf curl viruses of India. Plant Pathol. 2015, 64, 256–264. [Google Scholar] [CrossRef]
- Butterbach, P.; Verlaan, M.G.; Dullemans, A.; Lohuis, D.; Visser, R.G.F.; Bai, Y.; Kormelink, R. Tomato yellow leaf curl virus resistance by Ty-1 involves increased cytosine methylation of viral genomes and is compromised by cucumber mosaic virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 12942–12947. [Google Scholar] [CrossRef] [PubMed]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.F.; Scott, J.W.; Edwards, J.D.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD class RNA dependent RNA polymerase. PLoS Genet. 2013, 9, e1003300. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olive, E.; Briddon, R.W.; Hernandez-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Fraile, A.; Malpica, J.M.; Aranda, M.A.; Rodríguez-Cerezo, E.; García-Arenal, F. Genetic diversity in tobacco mild green mosaic tobamovirus infecting Nicotiana glauca. Virology 1996, 223, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Campos, S.; Díaz, J.A.; Monci, F.; Bejarano, E.R.; Reina, J.; Navas-Castillo, J.; Aranda, M.A.; Moriones, E. High genetic stability of the begomovirus tomato yellow leaf curl Sardinia virus in southern Spain over an 8-year period. Phytopathology 2002, 92, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, N.; Singh, A.; Basu, S.; Chakraborty, S. Differential response of diverse solanaceous hosts to tomato leaf curl New Delhi virus infection indicates coordinated action of NBS-LRR and RNAi-mediated host defense. Arch. Virol. 2015, 160, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Maule, A.J.; Leh, V.; Lederer, C. The dialogue between viruses and hosts in compatible interactions. Curr. Opin. Plant Biol. 2002, 5, 279–284. [Google Scholar] [CrossRef]
- Gillette, W.K.; Meade, T.J.; Jeffrey, J.L.; Petty, I.T.D. Genetic determinants of host-specificity in bipartite geminivirus DNA A components. Virology 1998, 251, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Petty, I.T.D.; Carter, S.C.; Morra, M.R.; Jeffrey, J.L.; Olivey, H.E. Bipartite geminivirus host adaptation determined cooperatively by coding and noncoding sequences of the genome. Virology 2000, 277, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Bisaro, D.M. Silencing suppression by geminivirus proteins. Virology 2006, 344, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Tomás, D.M.; Cañizares, C.; Abad, J.; Fernández-Muñoz, R.; Moriones, E. Resistance to tomato yellow leaf curl virus accumulation in the tomato wild relative Solanum habrochaites associated with the C4 viral protein. Mol. Plant Microbe Interact. 2011, 24, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Jeevalatha, A.; Siddappa, S.; Kumar, A.; Kaundal, P.; Guleria, A.; Sharma, S.; Nagesh, M.; Singh, B.P. An insight into differentially regulated genes in resistant and susceptible genotypes of potato in response to tomato leaf curl New Delhi virus-[potato] infection. Virus Res. 2017, 232, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Li, C.; Shi, Y.; Ryabov, E.; Huang, J.; Wu, Z.; Fan, Z.; Hong, Y. A single amino acid change in a geminiviral Rep protein differentiates between triggering a plant defence response and initiating viral DNA replication. J. Gen. Virol. 2008, 89, 2636–2641. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.C.; Garrido-Ramírez, E.R.; Sudarshana, M.R.; Yendluri, S.; Gilbertson, R.L. The N-terminus of the begomovirus nuclear shuttle protein (BV1) determines virulence or avirulence in Phaseolus vulgaris. Mol. Plant Microbe Interact. 2007, 20, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S. Distinct association of an alphasatellite and a betasatellite with tomato leaf curl New Delhi virus in field-infected cucurbit. J. Gen. Plant Pathol. 2017, 83, 185–188. [Google Scholar] [CrossRef]
- Saeed, M.; Zafar, Y.; Randles, J.W.; Rezaian, M.A. A monopartite begomovirus-associated DNA β satellite substitutes for the DNA B of a bipartite begomovirus to permit systemic infection. J. Gen. Virol. 2007, 88, 2881–2889. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, P.; Varma, A. Role of betasatellite in the pathogenesis of a bipartite begomovirus affecting tomato in India. Arch. Virol. 2012, 157, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.P.; Sharma, N.; Puranik, S.; Prasad, M. Post-transcriptional and epigenetic arms of RNA silencing: A defense machinery of naturally tolerant tomato plant against tomato leaf curl New Delhi virus. Plant Mol. Biol. Rep. 2014, 32, 1015–1029. [Google Scholar] [CrossRef]
- Singh, A.K.; Kushwaha, N.; Chakraborty, S. Synergistic interaction among begomoviruses leads to the suppression of host defense-related gene expression and breakdown of resistance in chilli. Appl. Microbiol. Biotechnol. 2016, 100, 4035–4049. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Moriones, E. Recombination as a motor of host switches and virus emergence: Geminiviruses as case studies. Curr. Opin. Virol. 2015, 10, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Wylie, S.J.; Jones, R.A.C. Role of recombination in the evolution of host specialization within bean yellow mosaic virus. Phytopathology 2009, 99, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Van der Walt, E.; Rybicki, E.P.; Varsani, A.; Polston, J.E.; Billharz, R.; Donaldson, L.; Monjane, A.L.; Martin, D.P. Rapid host adaptation by extensive recombination. J. Gen. Virol. 2009, 90, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Monci, F.; Sánchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses tomato yellow leaf curl Sardinia virus and tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.P.; Liu, Y.L.; Calvert, L.; Muñoz, C.; Otim-Nape, G.W.; Robinson, D.J.; Harrison, B.D. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol. 1997, 78, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Vanitharani, R.; Chattopadhyay, B.; Fauquet, C.M. Supervirulent pseudorecombination and asymmetric synergism between genomic components of two distinct species of begomovirus associated with severe tomato leaf curl disease in India. J. Gen. Virol. 2008, 89, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Kanakala, S.; Jyothsna, P.; Shukla, R.; Tiwari, N.; Veer, B.S.; Swarnalatha, P.; Krishnareddy, M.; Malathi, V.G. Asymmetric synergism and heteroencapsidation between two bipartite begomoviruses, tomato leaf curl New Delhi virus and tomato leaf curl Palampur virus. Virus Res. 2013, 174, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Singh, A.K.; Sharma, V.K.; Chattopadhyay, B.; Chakraborty, S. A novel recombinant tomato-infecting begomovirus capable of transcomplementing heterologous DNA-B components. Arch. Virol. 2011, 156, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.V.; Singh, D.; Singh, A.K.; Chakraborty, S. Molecular diversity, recombination and population structure of alphasatellites associated with begomovirus disease complexes. Infect. Genet. Evol. 2017, 49, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.V.; Prasanna, H.C.; Singh, A.K.; Ragunathan, D.; Garg, G.K.; Chakraborty, S. Molecular genetic analysis and evolution of begomoviruses and betasatellites causing yellow mosaic disease of bhendi. Virus Genes 2017, 53, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Mansoor, S.; Bedford, I.D.; Rishi, N.; Siwatch, S.S.; Zafar, Y.; Abdel-Salam, A.M.; Markham, P.G. Diversity of DNA 1: A satellite-like molecule associated with monopartite begomovirus-DNA β complexes. Virology 2004, 324, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, M.; Asad, S.; Zafar, Y.; Briddon, R.W.; Mansoor, S. Pepper leaf curl Lahore virus requires the DNA B component of tomato leaf curl New Delhi virus to cause leaf curl symptoms. Virol. J. 2010, 7, 367. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, P.; Singh, A.K.; Kumar, R.V.; Basu, S.; Chakraborty, S. Host specific adaptation of diverse betasatellites associated with distinct Indian tomato-infecting begomoviruses. Virus Genes 2014, 48, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Chattopadhyay, B.; Chakraborty, S. Biology and interactions of two distinct monopartite begomoviruses and betasatellites associated with radish leaf curl disease in India. Virol. J. 2012, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- De Barro, P.J.; Ahmed, M.Z. Genetic networking of the Bemisia tabaci cryptic species complex reveals pattern of biological invasions. PLoS ONE 2011, 6, e25579. [Google Scholar] [CrossRef] [PubMed]
- Janssen, D.; Simón, A.; Crespo, O.; Ruiz, L. Genetic population structure of Bemisia tabaci in Spain associated with tomato leaf curl New Delhi virus. Plant Protect. Sci. 2017, 53, 25–31. [Google Scholar]
- Nariani, T.K.; Vasudeva, R.S. Reaction of Lycopersicon. spp. to tomato leaf curl virus. Indian Phytopathol. 1963, 16, 238–239. [Google Scholar]
- Butter, N.S.; Rataul, H.S. The virus vector relationship of the tomato leaf curl virus (TLCV) and its vector Bemisia tabaci Gennadius (Homoptera: Aleyrodidae.). Phytoparasitica 1977, 5, 173–186. [Google Scholar] [CrossRef]
- Ghanim, M.; Czosnek, H. Interactions between the whitefly Bemisia tabaci and begomoviruses: Biological and genomic perspectives. In Management of Insect Pests to Agriculture: Lessons Learned from Deciphering Their Genome, Transcriptome and Proteome; Czosnek, H., Ghanim, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 181–200. [Google Scholar]
- Rana, V.S.; Popli, S.; Saurav, G.K.; Raina, H.S.; Chaubey, R.; Ramamurthy, V.V.; Rajagopal, R. A Bemisia tabaci midgut protein interacts with begomoviruses and plays a role in virus transmission. Cell. Microbiol. 2015, 18, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Legg, J.P.; Wintermantel, W.M.; Polston, J.E. Management of whitefly-transmitted viruses in open-field production systems. Control Plant Virus Dis. Seed-Propag. Crops 2014, 90, 147–206. [Google Scholar]
- Jones, R.A.C. Using epidemiological information to develop effective integrated virus disease management strategies. Virus Res. 2004, 100, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, M.; Rileyt, D.G.; Srinivasan, R. Insecticidal effects on the spatial progression of tomato yellow leaf curl virus and movement of its whitefly vector in tomato. J. Econ. Entomol. 2017, 110, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Jeger, M.; Chen, Z.; Cunningham, E.; Martin, G.; Powell, G. Population biology and epidemiology of plant virus epidemics: From tripartite to tritrophic interactions. Eur. J. Plant Pathol. 2012, 133, 3–23. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M. Breeding for resistance to whitefly-transmitted geminiviruses. Ann. Appl. Biol. 2002, 140, 109–127. [Google Scholar] [CrossRef]
- García-Andrés, S.; Tomás, D.M.; Navas-Castillo, J.; Moriones, E. Resistance-driven selection of begomoviruses associated with the tomato yellow leaf curl disease. Virus Res. 2009, 146, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Seal, S.E.; van den Bosch, F.; Jeger, M.J. Factors influencing begomovirus evolution and their increasing global significance: Implications for sustainable control. Crit. Rev. Plant Sci. 2006, 25, 23–46. [Google Scholar] [CrossRef]
- Rodriguez-López, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar] [PubMed]
- Tellez, M.D.M.; Simón, A.; Rodríguez, E.; Janssen, D. Control of tomato leaf curl New Delhi virus in zucchini using the predatory mite Amblyseius swirskii. Biol. Control 2017, 114, 106–113. [Google Scholar] [CrossRef]
- Sahu, P.P.; Sharma, N.; Puranik, S.; Chakraborty, S.; Prasad, M. Tomato 26S proteasome subunit RPT4a regulates ToLCNDV transcription and activates hypersensitive response in tomato. Sci. Rep. 2016, 6, 27078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, N.; Sahu, P.; Gupta, S.; Reddy, M.; Ravishankar, K.; Singh, M.; Sadashiva, A.; Prasad, M. Identification and validation of an ISSR marker linked to tomato leaf curl New Delhi virus resistant gene in a core set of tomato accessions. Veg. Sci. 2013, 40, 1–6. [Google Scholar]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Islam, S.; Das Munshi, A.; Verma, M.; Arya, L.; Mandal, B.; Behera, T.K.; Kumar, R.; Lal, S.K. Screening of Luffa cylindrica roem. for resistance against tomato leaf curl New Delhi virus, inheritance of resistance, and identification of SRAP markers linked to the single dominant resistance gene. J. Hortic. Sci. Biotechnol. 2011, 86, 661–667. [Google Scholar] [CrossRef]
- Saha, D.; Rana, R.S.; Sureja, A.K.; Verma, M.; Arya, L.; Das Munshi, A. Cloning and characterization of NBS-LRR encoding resistance gene candidates from tomato leaf curl New Delhi virus resistant genotype of Luffa cylindrica Roem. Physiol. Mol. Plant Pathol. 2013, 81, 107–117. [Google Scholar] [CrossRef]
- Ji, Y.; Scott, J.; Hanson, P.; Graham, E.; Maxwell, D. Sources of resistance, inheritance, and location of genetic loci conferring resistance to members of the tomato-infecting begomoviruses. In Tomato Yellow Leaf Curl Virus Disease; Czosnek, H., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 343–362. [Google Scholar]
- Hanson, P.; Green, S.; Kuo, G. Ty-2, a gene on chromosome 11 conditioning geminivirus resistance in tomato. Rep. Tomato Genet. Coop. 2006, 56, 17–18. [Google Scholar]
- Zamir, D.; Esktein-Michelson, I.; Zakay, Y.; Navot, N.; Zeidan, M. Mapping and introgression of a tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Genet. 1994, 88, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Eybishtz, A.; Peretz, Y.; Sade, D.; Gorovits, R.; Czosnek, H. Tomato yellow leaf curl virus infection of a resistant tomato line with a silenced sucrose transporter gene LeHT1 results in inhibition of growth, enhanced virus spread and necrosis. Planta 2009, 231, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Sade, D.; Eybishtz, A.; Gorovits, R.; Sobol, I.; Czosnek, H. A developmentally regulated lipocalin-like gene is overexpressed in tomato yellow leaf curl virus resistant tomato plants upon virus inoculation, and its silencing abolishes resistance. Plant Mol. Biol. 2012, 80, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Caro, M.; Hutton, S.F.; Scott, J.W.; Guo, Y.; Wang, X.; Rashid, M.H.; Szinay, D.; Jong, H.D.; Visser, R.G.F.; et al. Fine mapping of the tomato yellow leaf curl virus resistance gene Ty-2 on chromosome 11 of tomato. Mol. Breed. 2014, 34, 749–760. [Google Scholar] [PubMed]
- Prasanna, H.C.; Kashyap, S.; Krishna, R.; Sinha, D.P.; Reddy, S.; Malathi, V.G. Marker assisted selection of Ty-2 and Ty-3 carrying tomato lines and their implications in breeding tomato leaf curl disease resistant hybrids. Euphytica 2015, 204, 407–418. [Google Scholar] [CrossRef]
- Mubin, M.; Mansoor, S.; Hussain, M.; Zafar, Y. Silencing of the AV2 gene by antisense RNA protects transgenic plants against a bipartite begomovirus. Virol. J. 2007, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.K.; Singh, R.; Pandey, S.K.; Singh, B.P. Agrobacterium-mediated tomato transformation and regeneration of transgenic lines expressing tomato leaf curl virus coat protein gene for resistance against TLCV infection. Curr. Sci. 2005, 88, 1674–1679. [Google Scholar]
- Praveen, S.; Dasgupta, A.; Varma, A. Phylogenetic analysis and homologies of the replicase of tomato leaf curl geminiviruses: Implications for obtaining pathogen derived resistance. Virus Genes 2004, 28, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Praveen, S.; Mishra, A.K.; Dasgupta, A. Antisense suppression of replicase gene expression recovers tomato plants from leaf curl infection. Plant Sci. 2005, 168, 1011–1014. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Mishra, A.K.; Praveen, S. Hairpin RNA-mediated strategies for silencing of tomato leaf curl virus AC1 and AC4 Genes for effective resistance in plants. Oligonucleotide 2007, 17, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Taneja, J.; Dasgupta, I.; Mukherjee, S.K. Development of plants resistant to tomato geminiviruses using artificial trans-acting small interfering RNA. Mol. Plant Pathol. 2015, 16, 724–734. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriones, E.; Praveen, S.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. https://doi.org/10.3390/v9100264
Moriones E, Praveen S, Chakraborty S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses. 2017; 9(10):264. https://doi.org/10.3390/v9100264
Chicago/Turabian StyleMoriones, Enrique, Shelly Praveen, and Supriya Chakraborty. 2017. "Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops" Viruses 9, no. 10: 264. https://doi.org/10.3390/v9100264
APA StyleMoriones, E., Praveen, S., & Chakraborty, S. (2017). Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses, 9(10), 264. https://doi.org/10.3390/v9100264