
Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids



Abstract
:1. Introduction
2. Materials and Methods
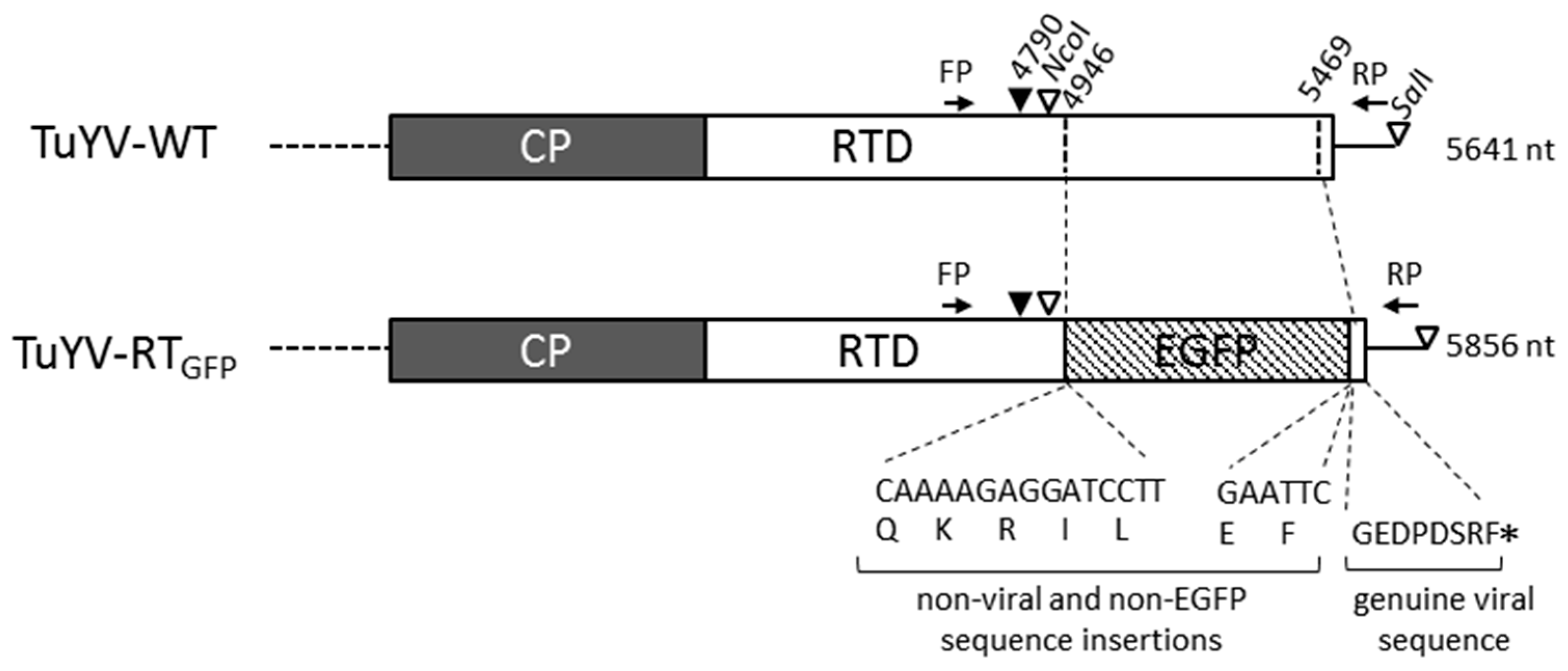
2.1. TuYV Constructs
2.2. Protoplasts and Plant Infection
2.3. Detection of Viral RNA and Capsid Proteins
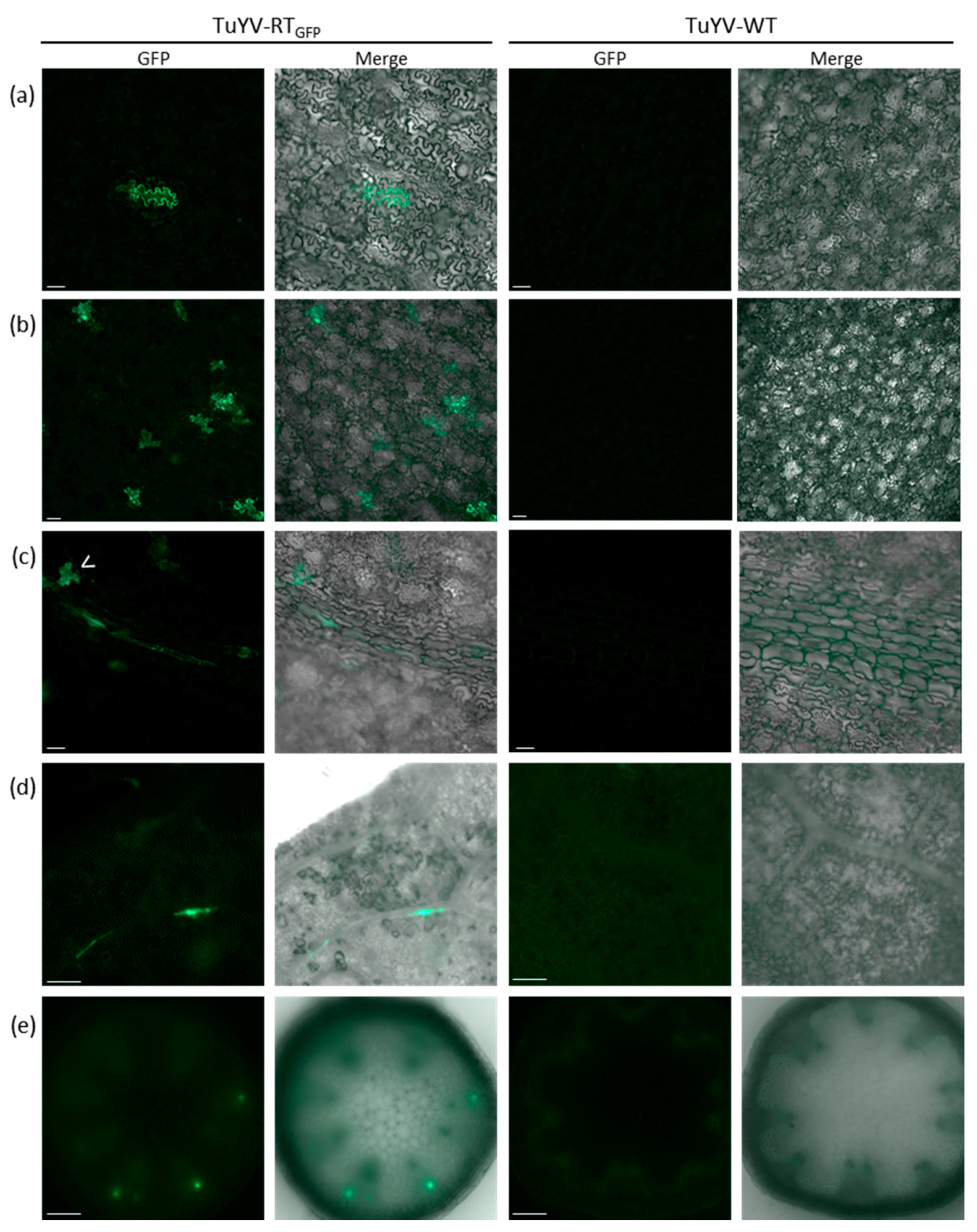
2.4. Epifluorescence Microscopy
3. Results
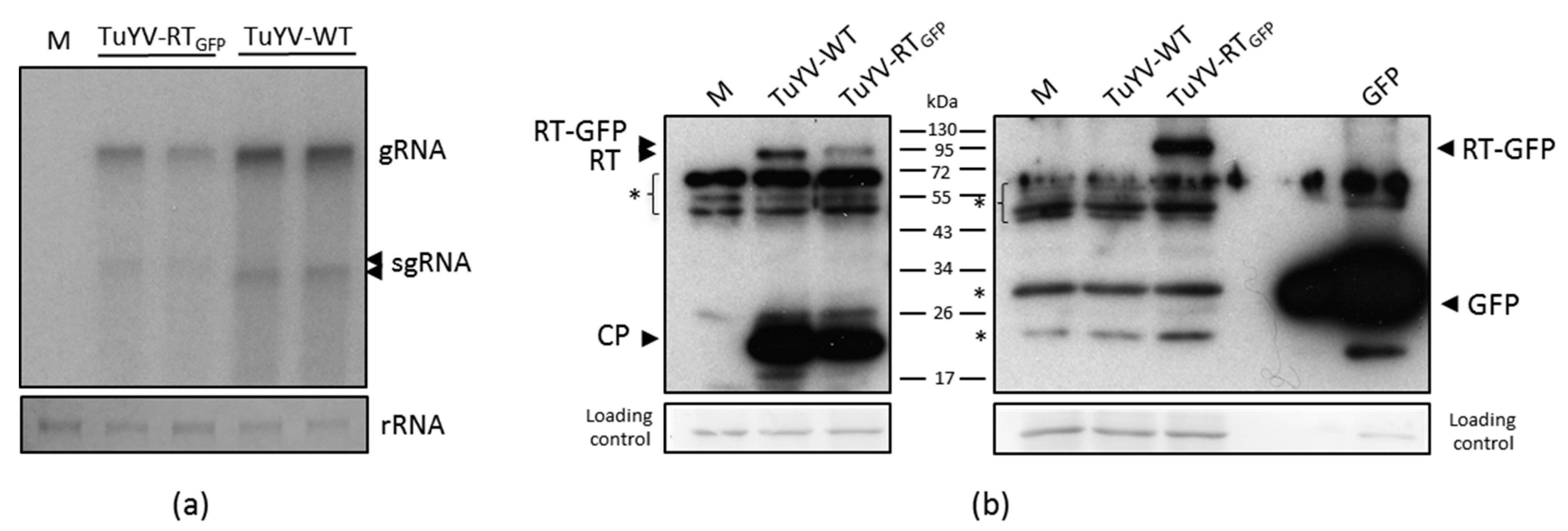
3.1. Infectivity of TuYV-RTGFP in Protoplasts and Inoculated Leaves
3.2. Viral Particle Formation
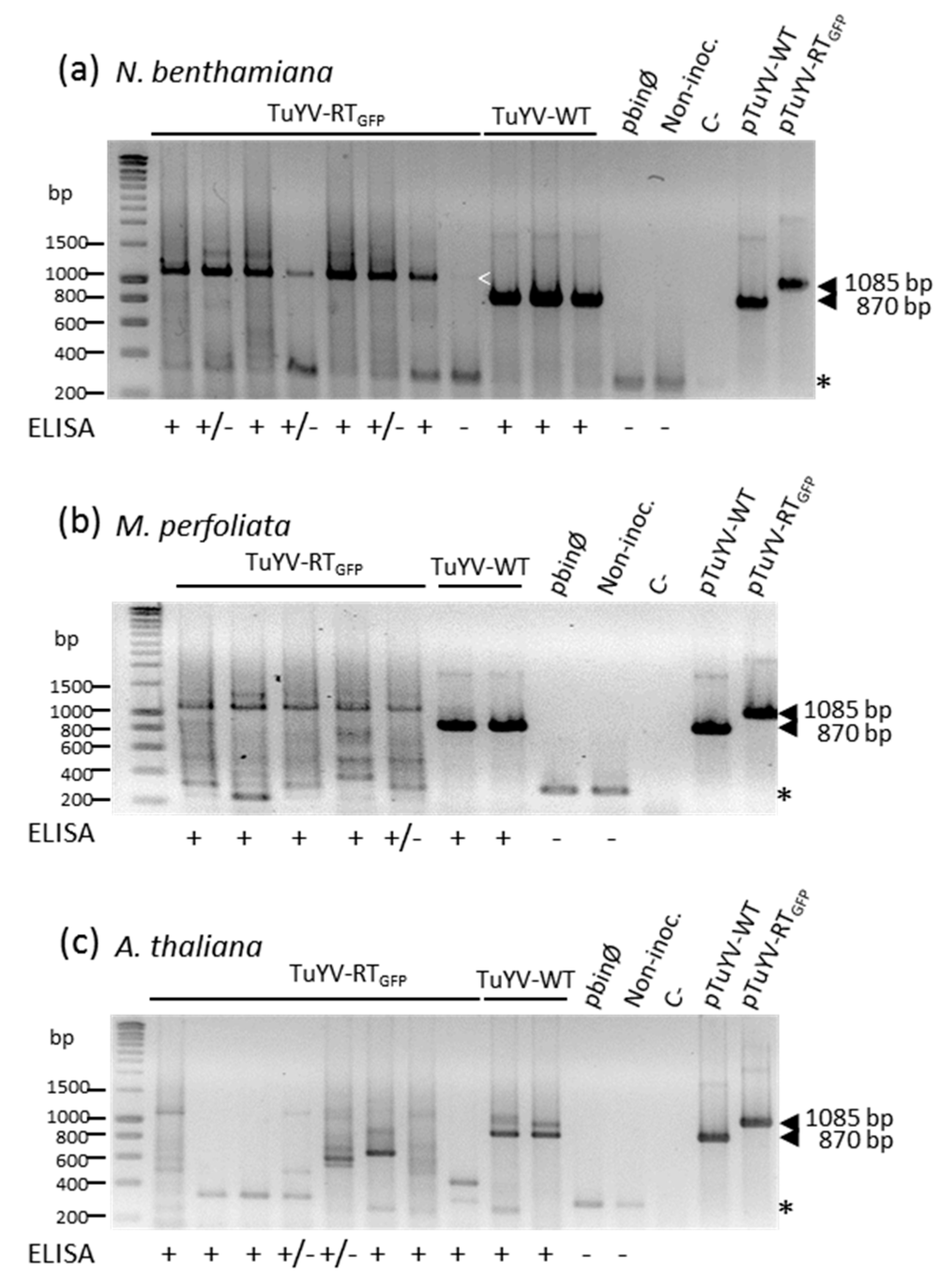
3.3. Systemic Movement of TuYV-RTGFP
3.4. Aphid Transmissibility of TuYV-RTGFP
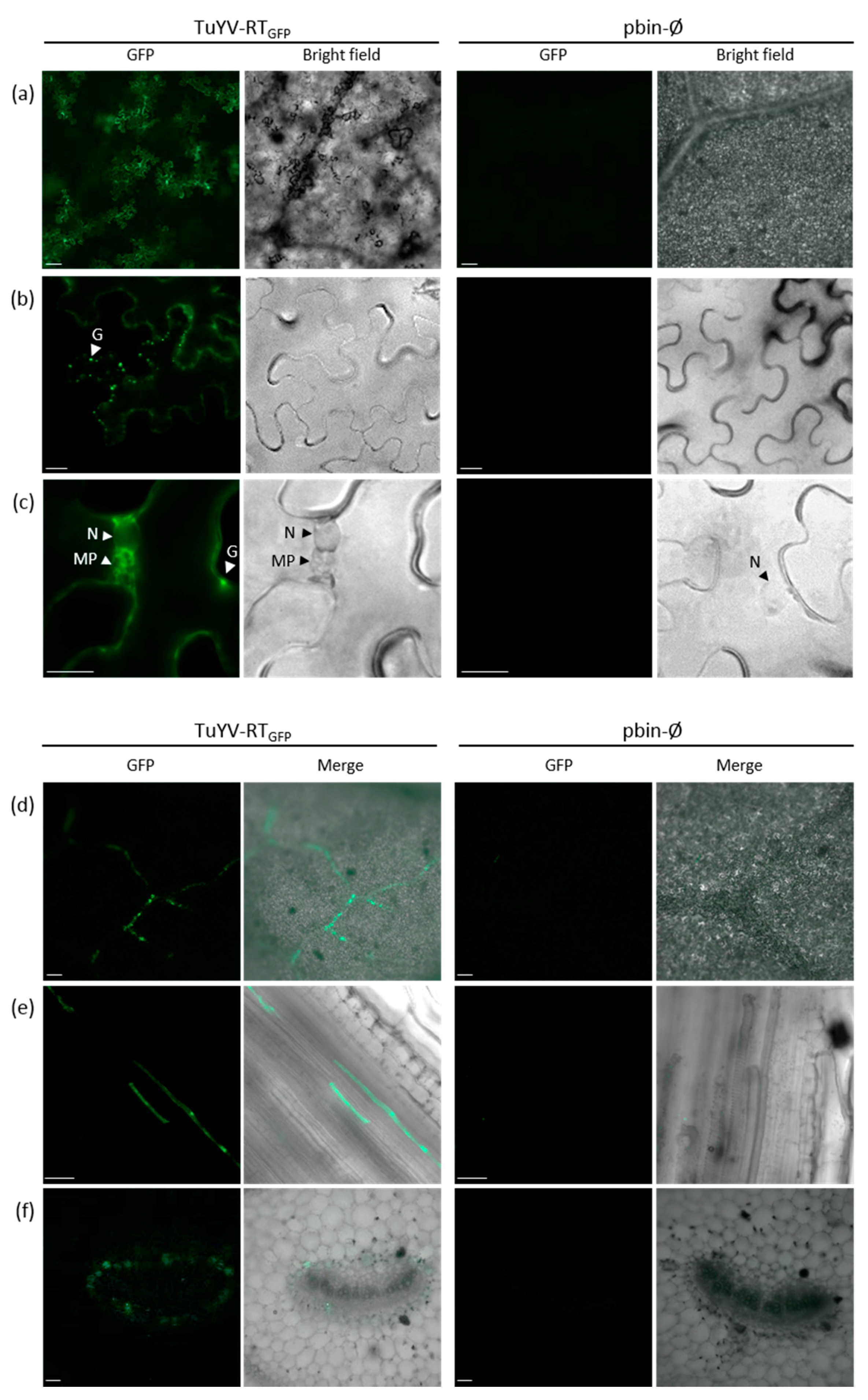
3.5. In Vivo Tracking of TuYV-RTGFP Following Agroinoculation or Aphid Transmission
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beuve, M.; Stevens, M.; Liu, H.Y.; Wintermantel, W.M.; Hauser, S.; Lemaire, O. Biological and molecular characterization of an american sugar beet-infecting Beet western yellows virus isolate. Plant Dis. 2008, 92, 51–60. [Google Scholar] [CrossRef]
- Smirnova, E.; Firth, A.E.; Miller, W.A.; Scheidecker, D.; Brault, V.; Reinbold, C.; Rakotondrafara, A.M.; Chung, B.Y.; Ziegler-Graff, V. Discovery of a small non-aug-initiated ORF in poleroviruses and luteoviruses that is required for long-distance movement. PLoS Pathog. 2015, 11, e1004868. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, C.J.; De Zoeten, G.A. Beet yellows western virus in phloem tissue of Thlaspi arvense. Phytopathology 1979, 69, 1194–1198. [Google Scholar] [CrossRef]
- Esau, K.; Hoefert, L.L. Ultrastructure of sugarbeet leaves infected with beet western yellows virus. J. Ultrastruct. Res. 1972, 40, 556–571. [Google Scholar] [CrossRef]
- Mutterer, J.D.; Stussi-Garaud, C.; Michler, P.; Richards, K.E.; Jonard, G.; Ziegler-Graff, V. Role of the beet western yellows virus readthrough protein in virus movement in Nicotiana clevelandii. J. Gen. Virol. 1999, 80, 2771–2778. [Google Scholar] [CrossRef] [PubMed]
- Shepardson, S.; Esau, K.; McCrum, R. Ultrastructure of potato leaf phloem infected with potato leafroll virus. Virology 1980, 105, 379–392. [Google Scholar] [CrossRef]
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. C. R. Biol. 2010, 333, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; Herrbach, E.; Reinbold, C. Electron microscopy studies on luteovirid transmission by aphids. Micron 2007, 38, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Baumberger, N.; Tsai, C.H.; Lie, M.; Havecker, E.; Baulcombe, D.C. The polerovirus silencing suppressor P0 targets argonaute proteins for degradation. Curr. Biol. 2007, 17, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Bortolamiol, D.; Pazhouhandeh, M.; Marrocco, K.; Genschik, P.; Ziegler-Graff, V. The polerovirus F box protein P0 targets ARGONAUTE1 to suppress RNA silencing. Curr. Biol. 2007, 17, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Lozsa, R.; Hutvagner, G.; Burgyan, J. Polerovirus protein P0 prevents the assembly of small RNA-containing RISC complexes and leads to degradation of ARGONAUTE1. Plant J. 2010, 62, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Derrien, B.; Baumberger, N.; Schepetilnikov, M.; Viotti, C.; De Cillia, J.; Ziegler-Graff, V.; Isono, E.; Schumacher, K.; Genschik, P. Degradation of the antiviral component argonaute1 by the autophagy pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 15942–15946. [Google Scholar] [CrossRef] [PubMed]
- Pazhouhandeh, M.; Dieterle, M.; Marrocco, K.; Lechner, E.; Berry, B.; Brault, V.; Hemmer, O.; Kretsch, T.; Richards, K.E.; Genschik, P.; et al. F-box-like domain in the polerovirus protein P0 is required for silencing suppressor function. Proc. Natl. Acad. Sci. USA 2006, 103, 1994–1999. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, D.; Kawchuk, L.; Monecke, M.; Nowok, S.; Fischer, R.; Rohde, W. Immunological analysis of potato leafroll luteovirus (PLRV) P1 expression identifies a 25 kDa RNA-binding protein derived via P1 processing. Nucleic Acids Res. 1999, 27, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Van der Wilk, F.; Verbeek, M.; Dullemans, A.M.; van den Heuvel, J.F. The genome-linked protein of potato leafroll virus is located downstream of the putative protease domain of the ORF1 product. Virology 1997, 234, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Bahner, I.; Lamb, J.; Mayo, M.A.; Hay, R.T. Expression of the genome of potato leafroll virus: Readthrough of the coat protein termination codon in vivo. J. Gen. Virol. 1990, 71, 2251–2256. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; van den Heuvel, J.F.; Verbeek, M.; Ziegler-Graff, V.; Reutenauer, A.; Herrbach, E.; Garaud, J.C.; Guilley, H.; Richards, K.; Jonard, G. Aphid transmission of beet western yellows luteovirus requires the minor capsid read-through protein P74. EMBO J. 1995, 14, 650–659. [Google Scholar] [PubMed]
- Wang, J.Y.; Chay, C.; Gildow, F.E.; Gray, S.M. Readthrough protein associated with virions of barley yellow dwarf luteovirus and its potential role in regulating the efficiency of aphid transmission. Virology 1995, 206, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Boissinot, S.; Erdinger, M.; Monsion, B.; Ziegler-Graff, V.; Brault, V. Both structural and non-structural forms of the readthrough protein of cucurbit aphid-borne yellows virus are essential for efficient systemic infection of plants. PLoS ONE 2014, 9, e93448. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; Bergdoll, M.; Mutterer, J.; Prasad, V.; Pfeffer, S.; Erdinger, M.; Richards, K.E.; Ziegler-Graff, V. Effects of point mutations in the major capsid protein of beet western yellows virus on capsid formation, virus accumulation, and aphid transmission. J. Virol. 2003, 77, 3247–3256. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; Mutterer, J.; Scheidecker, D.; Simonis, M.T.; Herrbach, E.; Richards, K.; Ziegler-Graff, V. Effects of point mutations in the readthrough domain of the beet western yellows virus minor capsid protein on virus accumulation in planta and on transmission by aphids. J. Virol. 2000, 74, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Bruyère, A.; Brault, V.; Ziegler-Graff, V.; Simonis, M.T.; Van den Heuvel, J.F.; Richards, K.; Guilley, H.; Jonard, G.; Herrbach, E. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging and on virus accumulation, symptoms, and aphid transmission. Virology 1997, 230, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Peter, K.A.; Gildow, F.; Palukaitis, P.; Gray, S.M. The C terminus of the polerovirus P5 readthrough domain limits virus infection to the phloem. J. Virol. 2009, 83, 5419–5429. [Google Scholar] [CrossRef] [PubMed]
- Peter, K.A.; Liang, D.; Palukaitis, P.; Gray, S.M. Small deletions in the potato leafroll virus readthrough protein affect particle morphology, aphid transmission, virus movement and accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Medina, C.; Boissinot, S.; Chapuis, S.; Gereige, D.; Rastegar, M.; Erdinger, M.; Revers, F.; Ziegler-Graff, V.; Brault, V. A protein kinase binds the C-terminal domain of the readthrough protein of Turnip yellows virus and regulates virus accumulation. Virology 2015, 486, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Hofius, D.; Herbers, K.; Melzer, M.; Omid, A.; Tacke, E.; Wolf, S.; Sonnewald, U. Evidence for expression level-dependent modulation of carbohydrate status and viral resistance by the potato leafroll virus movement protein in transgenic tobacco plants. Plant J. 2001, 28, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Palukaitis, P.; Gray, S.M. Host-dependent requirement for the potato leafroll virus 17-kDa protein in virus movement. Mol. Plant Microbe Interact. 2002, 15, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.; Hofius, D.; Sonnewald, U. Intracellular trafficking of potato leafroll virus movement protein in transgenic arabidopsis. Traffic 2007, 8, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Graff, V.; Brault, V.; Mutterer, J.; Simonis, M.T.; Herrbach, E.; Guilley, H.; Richards, K.; Jonard, G. The coat protein of beet western yellows luteovirus is essential for systemic infection but the viral gene products P29 and P19 are dispensable for systemic infection and aphid transmission. Mol. Plant Microbe Interact. 1996, 9, 501–510. [Google Scholar] [CrossRef]
- Nurkiyanova, K.M.; Ryabov, E.V.; Commandeur, U.; Duncan, G.H.; Canto, T.; Gray, S.M.; Mayo, M.A.; Taliansky, M.E. Tagging potato leafroll virus with the jellyfish green fluorescent protein gene. J. Gen. Virol. 2000, 81, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; Vigano, F. Production of a full-length infectious GFP-tagged cDNA clone of beet mild yellowing virus for the study of plant-polerovirus interactions. Virus Genes 2007, 34, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Gurtu, V.; Kain, S.R. An enhanced green fluorescent protein allows sensitive detection of gene transfer in mammalian cells. Biochem. Biophys. Res. Commun. 1996, 227, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Leiser, R.M.; Ziegler-Graff, V.; Reutenauer, A.; Herrbach, E.; Lemaire, O.; Guilley, H.; Richards, K.; Jonard, G. Agroinfection as an alternative to insects for infecting plants with beet western yellows luteovirus. Proc. Natl. Acad. Sci. USA 1992, 89, 9136–9140. [Google Scholar] [CrossRef] [PubMed]
- Holsters, M.; Silva, B.; Van Vliet, F.; Genetello, C.; De Block, M.; Dhaese, P.; Depicker, A.; Inze, D.; Engler, G.; Villarroel, R.; et al. The functional organization of the nopaline A. Tumefaciens plasmid pTIC58. Plasmid 1980, 3, 212–230. [Google Scholar] [CrossRef]
- English, J.J.; Davenport, G.F.; Elmayan, T.E.; Vaucheret, H.; Baulcombe, D.C. Requirement of sense transcription for homology-dependent virus resistance and trans-inactivation. Plant J. 1997, 12, 597–603. [Google Scholar] [CrossRef]
- Van den Heuvel, J.F.J.M.; Boerma, T.M.; Peters, D. Transmission of potato leafroll virus from plants and artificial diets by Myzus persicae. Phytopathology 1991, 81, 150–154. [Google Scholar] [CrossRef]
- Reutenauer, A.; Ziegler-Graff, V.; Lot, H.; Scheidecker, D.; Guilley, H.; Richards, K.; Jonard, G. Identification of beet western yellows luteovirus genes implicated in viral replication and particle morphogenesis. Virology 1993, 195, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Veidt, I.; Bouzoubaa, S.E.; Leiser, R.M.; Ziegler-Graff, V.; Guilley, H.; Richards, K.; Jonard, G. Synthesis of full-length transcripts of beet western yellows virus RNA: Messenger properties and biological activity in protoplasts. Virology 1992, 186, 192–200. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Revollon, S.; Strub, J.M.; Fitchette, A.C.; Wiss, L.; Gomord, V.; Van Dorsselaer, A.; Brault, V. A reinvestigation provides no evidence for sugar residues on structural proteins of poleroviruses and argues against a role for glycosylation of virus structural proteins in aphid transmission. Virology 2010, 402, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Hipper, C.; Monsion, B.; Bortolamiol-Becet, D.; Ziegler-Graff, V.; Brault, V. Formation of virions is strictly required for Turnip yellows virus long-distance movement in plants. J. Gen. Virol. 2014, 95, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Imlau, A.; Truernit, E.; Sauer, N. Cell-to-cell and long-distance trafficking of the green fluorescent protein in the phloem and symplastic unloading of the protein into sink tissues. Plant Cell 1999, 11, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Stadler, R.; Wright, K.M.; Lauterbach, C.; Amon, G.; Gahrtz, M.; Feuerstein, A.; Oparka, K.J.; Sauer, N. Expression of GFP-fusions in Arabidopsis companion cells reveals non-specific protein trafficking into sieve elements and identifies a novel post-phloem domain in roots. Plant J. 2005, 41, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Aguero, J.; Vives, M.C.; Velazquez, K.; Ruiz-Ruiz, S.; Juarez, J.; Navarro, L.; Moreno, P.; Guerri, J. Citrus leaf blotch virus invades meristematic regions in Nicotiana benthamiana and citrus. Mol. Plant Pathol. 2013, 14, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Le Gall, O.; Maisonneuve, B.; German-Retana, S.; Redondo, E. The use of green fluorescent protein-tagged recombinant viruses to test lettuce mosaic virus resistance in lettuce. Phytopathology 2002, 92, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.S.; Roberts, A.G.; Prior, D.A.; Chapman, S.; Oparka, K.J. Cell-to-cell and phloem-mediated transport of potato virus X. The role of virions. Plant Cell 1998, 10, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-of-susceptibility mutants of Arabidopsis thaliana reveal an essential role for eIF(iso)4E during potyvirus infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef]
- Matevz, R.; Florence, F.; Michel, T.; Ion, G.A.; Agnes, D.; Laurent, G.; Maja, K.; David, D.; Kristina, G.; Emmanuel, J.; et al. Fluorescently tagged potato virus Y: A versatile tool for functional analysis of plant-virus interactions. Mol. Plant Microbe Interact. 2015, 28, 739–750. [Google Scholar] [PubMed]
- Naderpour, M.; Johansen, I.E. Visualization of resistance responses in Phaseolus vulgaris using reporter tagged clones of Bean common mosaic virus. Virus Res. 2011, 159, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; McMechan, A.J.; Bartels, M.; Hein, G.L.; Graybosch, R.A. In vitro transcripts of wild-type and fluorescent protein-tagged Triticum mosaic virus (family Potyviridae) are biologically active in wheat. Phytopathology 2015, 105, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Krenz, B.; Bronikowski, A.; Lu, X.; Ziebell, H.; Thompson, J.R.; Perry, K.L. Visual monitoring of Cucumber mosaic virus infection in Nicotiana benthamiana following transmission by the aphid vector Myzus persicae. J. Gen. Virol. 2015, 96, 2904–2912. [Google Scholar] [CrossRef] [PubMed]
- Bordat, A.; Houvenaghel, M.C.; German-Retana, S. Gibson assembly: An easy way to clone potyviral full-length infectious cDNA clones expressing an ectopic VPg. Virol. J. 2015, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Cotton, S.; Grangeon, R.; Thivierge, K.; Mathieu, I.; Ide, C.; Wei, T.; Wang, A.; Laliberte, J.F. Turnip mosaic virus RNA replication complex vesicles are mobile, align with microfilaments, and are each derived from a single viral genome. J. Virol. 2009, 83, 10460–10471. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Wu, Z.; Wang, A. The multifunctional protein CI of potyviruses plays interlinked and distinct roles in viral genome replication and intercellular movement. Virol. J. 2015, 12, 141. [Google Scholar] [CrossRef] [PubMed]
- Tilsner, J.; Linnik, O.; Wright, K.M.; Bell, K.; Roberts, A.G.; Lacomme, C.; Santa Cruz, S.; Oparka, K.J. The TGB1 movement protein of potato virus X reorganizes actin and endomembranes into the X-body, a viral replication factory. Plant Physiol. 2012, 158, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Ferriol, I.; Turina, M.; Zamora-Macorra, E.J.; Falk, B.W. RNA1-independent replication and GFP expression from Tomato marchitez virus isolate M cloned cDNA. Phytopathology 2016, 106, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Venkataraman, S.; Li, C.; Wang, W.; Dayan-Glick, C.; Mawassi, M. Construction and biological activities of the first infectious cDNA clones of the genus Foveavirus. Virology 2013, 435, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Poque, S.; Pagny, G.; Ouibrahim, L.; Chague, A.; Eyquard, J.P.; Caballero, M.; Candresse, T.; Caranta, C.; Mariette, S.; Decroocq, V. Allelic variation at the rpv1 locus controls partial resistance to Plum pox virus infection in Arabidopsis thaliana. BMC Plant Biol. 2015, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Verver, J.; Wellink, J.; Van Lent, J.; Gopinath, K.; Van Kammen, A. Studies on the movement of cowpea mosaic virus using the jellyfish green fluorescent protein. Virology 1998, 242, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xiao, C.; Han, K.; Peng, J.; Lin, L.; Lu, Y.; Xie, L.; Wu, X.; Xu, P.; Li, G.; et al. Development of an agroinoculation system for full-length and GFP-tagged cDNA clones of cucumber green mottle mosaic virus. Arch. Virol. 2015, 160, 2867–2872. [Google Scholar] [CrossRef] [PubMed]
- Cadena-Nava, R.D.; Comas-Garcia, M.; Garmann, R.F.; Rao, A.L.; Knobler, C.M.; Gelbart, W.M. Self-assembly of viral capsid protein and RNA molecules of different sizes: Requirement for a specific high protein/RNA mass ratio. J. Virol. 2012, 86, 3318–3326. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Morris, T.J. Encapsidation of turnip crinkle virus is defined by a specific packaging signal and RNA size. J. Virol. 1997, 71, 1428–1435. [Google Scholar] [PubMed]
- Novoa, R.R.; Calderita, G.; Arranz, R.; Fontana, J.; Granzow, H.; Risco, C. Virus factories: Associations of cell organelles for viral replication and morphogenesis. Biol. Cell 2005, 97, 147–172. [Google Scholar] [CrossRef] [PubMed]
- Haupt, S.; Stroganova, T.; Ryabov, E.; Kim, S.H.; Fraser, G.; Duncan, G.; Mayo, M.A.; Barker, H.; Taliansky, M. Nucleolar localization of potato leafroll virus capsid proteins. J. Gen. Virol. 2005, 86, 2891–2896. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F.; Hogen Esch, T. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nicotiana benthamiana | Montia perfoliata | Arabidopsis thaliana | ||||
|---|---|---|---|---|---|---|
| Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | |
| TuYV-RTGFP | 4/4 | 2.59 ± 0.42 | 3/3 | 2.11 ± 0.15 | 4/4 | 1.80 ± 0.87 |
| TuYV-WT | 3/3 | 2.87 ± 0.05 | 4/4 | 2.61 ± 0.20 | 4/4 | 1.83 ± 0.59 |
| pbin-Ø | 0/2 | 0.16 ± 0.00 | 0/2 | 0.14 ± 0.01 | 0/2 | 0.11 ± 0.01 |
| Non-inoc. 3 | 0/2 | 0.15 ± 0.01 | 0/2 | 0.14 ± 0.00 | 0/1 | 0.10 ± 0.00 |
| Nicotiana benthamiana | Montia perfoliata | Arabidopsis thaliana | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp.1 | Exp.2 | Exp.3 | Exp.1 | Exp.2 | Exp.1 | Exp.2 | ||||||||
| Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | Nb pl. inf./pl. inoc. 1 | OD ± SD 2 | |
| TuYV-RTGFP | 4/8 (50%) | 1.12 ± 0.59 | 13/17 (76%) | 0.52 ± 0.23 | 4/7 (57%) | 0.54 ± 0.12 | 4/6 (67%) | 1.42 ± 0.69 | 6/8 (75%) | 0.70 ± 0.34 | 6/16 (38%) | 0.31 ± 0.13 | 2/9 (22%) | 0.69 ± 0.16 |
| TuYV-WT | 8/8 (100%) | 1.86 ± 0.30 | 4/4 (100%) | 0.63 ± 0.22 | 6/7 (86%) | 1.61 ± 0.79 | 4/6 (67%) | 1.58 ± 0.62 | 7/8 (88%) | 1.90 ± 0.24 | 8/8 (100%) | 1.70 ± 0.16 | 4/6 (67%) | 1.40 ± 1.23 |
| pbin-Ø | 0/2 | 0.14 ± 0.30 | 0/1 | 0.11 | 0/1 | 0.15 | 0/2 | 0.13 ± 0.00 | 0/2 | 0.11 ± 0.01 | 0/2 | 0.10 ± 0.00 | 0/2 | 0.12 ± 0.00 |
| Non-inoc.3 | 0/3 | 0.14 ± 0.00 | 0/3 | 0.13 ± 0.01 | 0/3 | 0.16 ± 0.01 | 0/2 | 0.11 ± 0.00 | 0/3 | 0.10 ± 0.01 | 0/3 | 0.10 ± 0.01 | 0/3 | 0.11 ± 0.01 |
| N. benthamiana 1 | M. perfoliata 1 | A. thaliana 1 | ||
|---|---|---|---|---|
| Exp.1 | Exp.1 | Exp.2 | Exp.1 | |
| Nb pl.inf./pl. inoc. 2 | Nb pl.inf./pl. inoc. 2 | Nb pl.inf./pl. inoc. 2 | Nb pl.inf./pl. inoc. 2 | |
| TuYV-RTGFP | 10/10 | 9/10 | 5/10 | 4/10 |
| TuYV-WT | 9/10 | 10/10 | 10/10 | 10/10 |
| Non-inoc. 3 | 0/3 | 0/3 | 0/3 | 0/3 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boissinot, S.; Pichon, E.; Sorin, C.; Piccini, C.; Scheidecker, D.; Ziegler-Graff, V.; Brault, V. Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids. Viruses 2017, 9, 166. https://doi.org/10.3390/v9070166
Boissinot S, Pichon E, Sorin C, Piccini C, Scheidecker D, Ziegler-Graff V, Brault V. Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids. Viruses. 2017; 9(7):166. https://doi.org/10.3390/v9070166
Chicago/Turabian StyleBoissinot, Sylvaine, Elodie Pichon, Céline Sorin, Céline Piccini, Danièle Scheidecker, Véronique Ziegler-Graff, and Véronique Brault. 2017. "Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids" Viruses 9, no. 7: 166. https://doi.org/10.3390/v9070166
APA StyleBoissinot, S., Pichon, E., Sorin, C., Piccini, C., Scheidecker, D., Ziegler-Graff, V., & Brault, V. (2017). Systemic Propagation of a Fluorescent Infectious Clone of a Polerovirus Following Inoculation by Agrobacteria and Aphids. Viruses, 9(7), 166. https://doi.org/10.3390/v9070166