
Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Origin
2.3. Sample Preparation
2.4. Total Phenols Determination
2.5. Radical Scavenging Activity
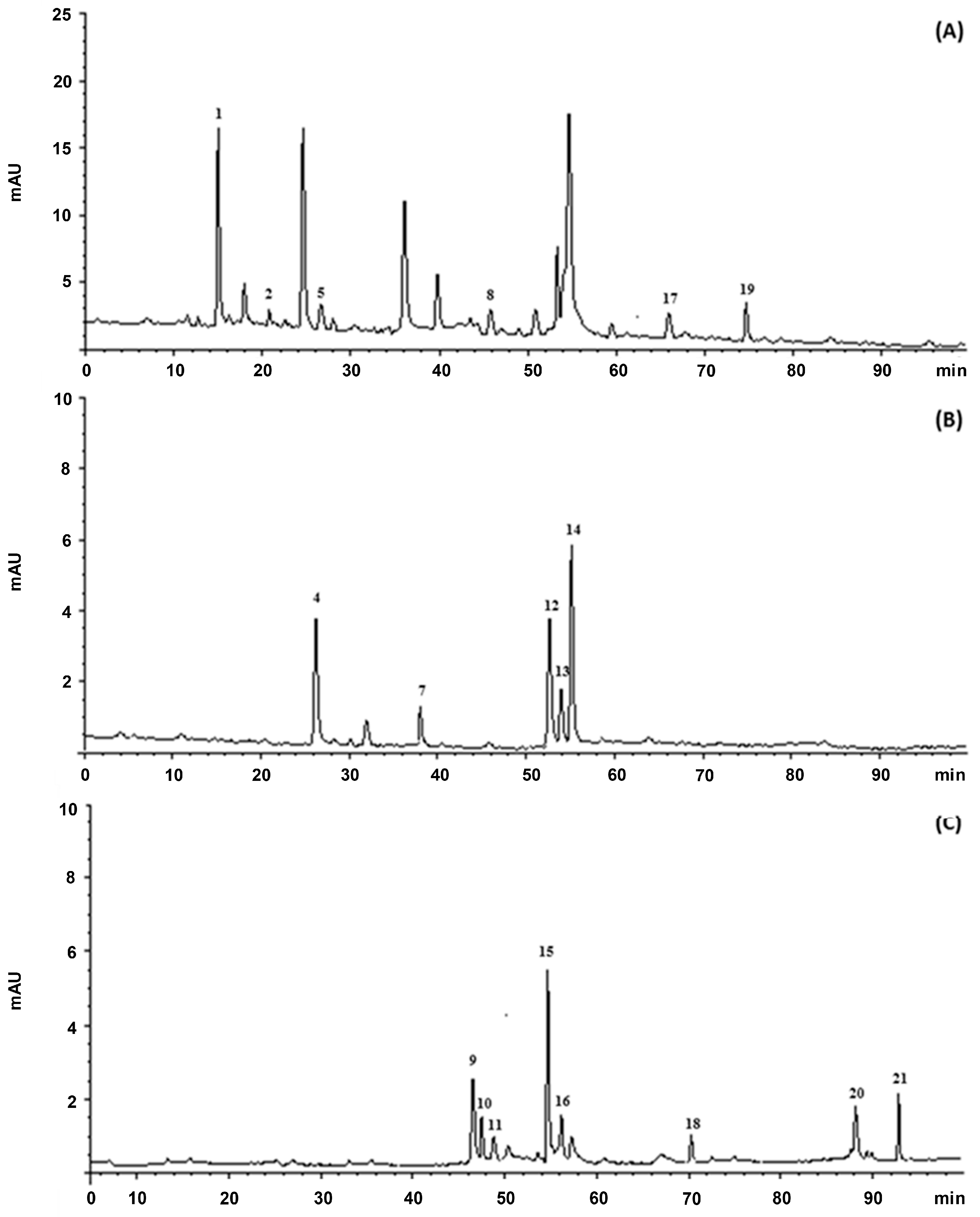
2.6. Determination of Polyphenolic Profile
2.7. Cells Culture and Virus
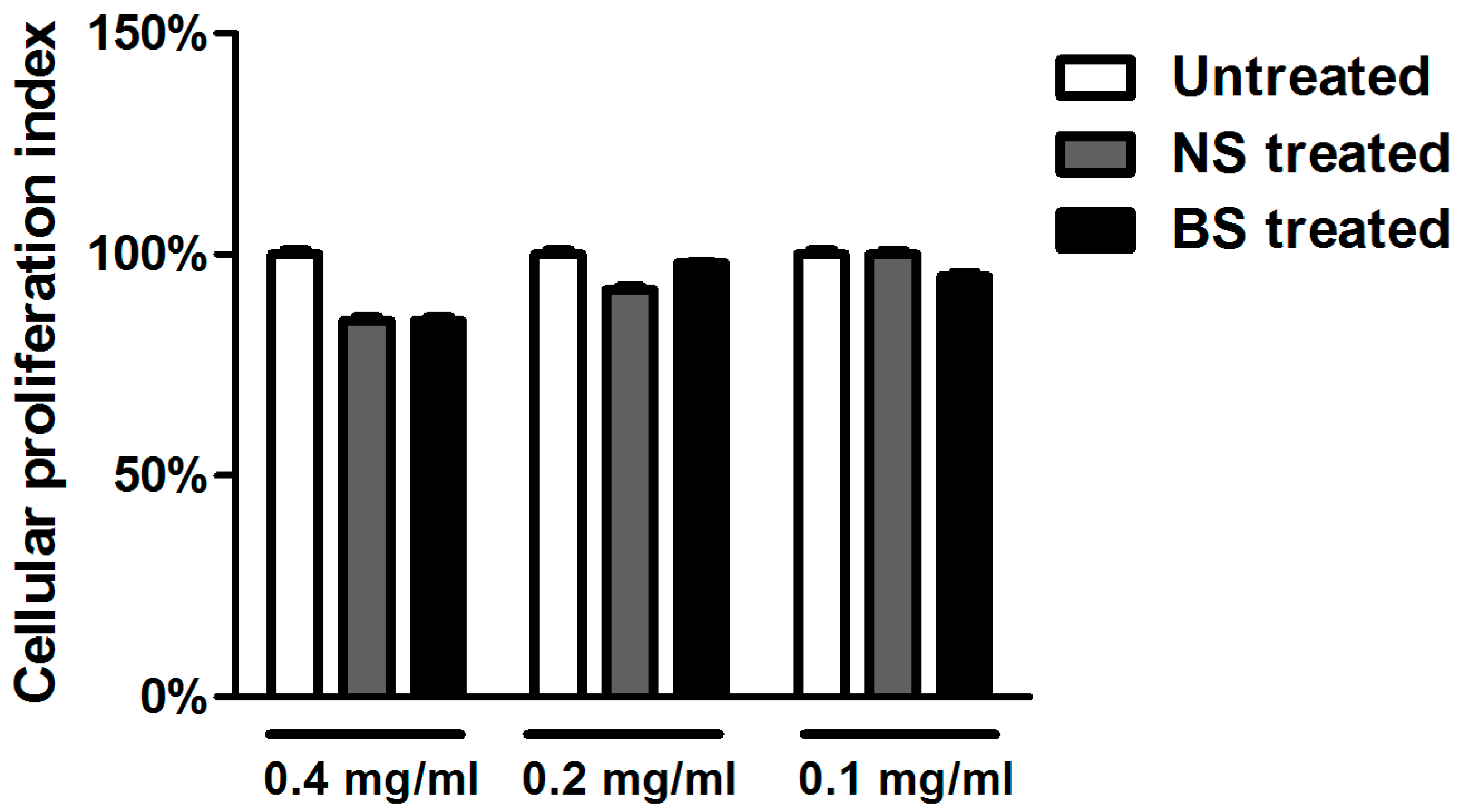
2.8. Cell Proliferation Assay
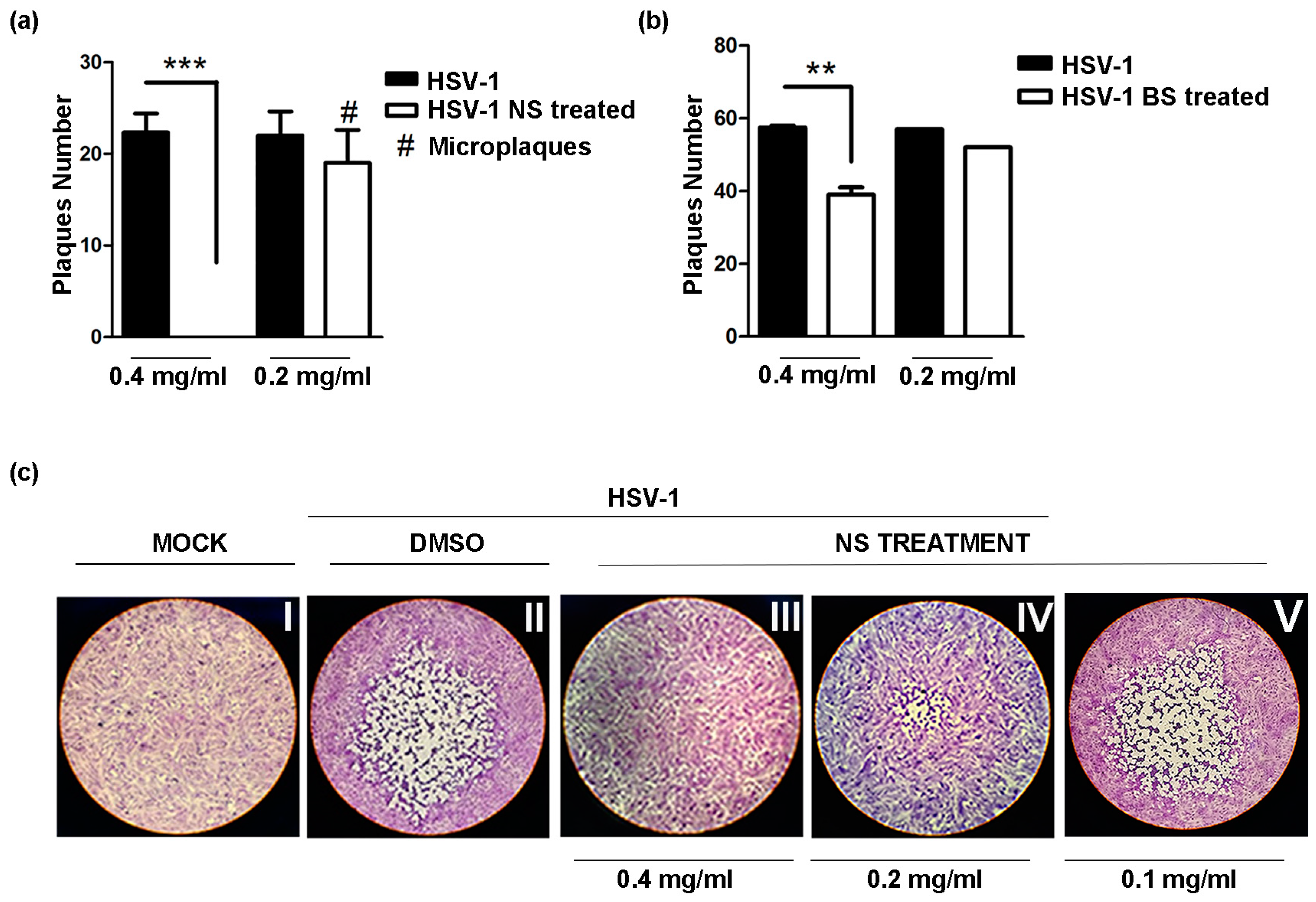
2.9. Plaque Reduction Assay
2.10. Protein Extraction and Immunoblot Analysis
2.11. DNA Extraction and Quantitative Real-Time PCR
2.12. The Binding Assay
2.13. Statistical Analysis
3. Results
3.1. Total Phenol Content and Radical Scavenging Activity of NS and BS Extracts
3.2. Cytotoxicity of Almond Extracts on Cells Cultures
3.3. Antiviral Activity of the Almond Extracts
3.4. Inhibition of HSV Replication by NS and BS Extracts
3.5. HSV Attachment to Target Cells Is Prevented by Almond Extracts
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Milbury, P.E.; Chen, C.Y.; Dolnikowski, G.G.; Blumberg, J.B. Determination of flavonoids and phenolics and their distribution in almonds. J. Agric. Food Chem. 2006, 54, 5027–5033. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Tomaino, A.; Arcoraci, T.; Martorana, M.; Lo Turco, V.; Cacciola, F.; Rich, G.T.; Bisignano, C.; Saija, A.; Dugo, P.; et al. Characterization of polyphenols, lipids and dietary fiber from almond skins (Amygdalus communis L.). J. Food Compost. Anal. 2010, 23, 166–174. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, X.; Huang, G.; Zhang, W.; Rao, P.; Ni, L. Prebiotic effects of almonds and almond skins on intestinal microbiota in healthy adult humans. Anaerobe 2014, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Garrido, I.; Lebrón-Aguilar, R.; Bartolome, B.; Gómez-Cordovés, C. Almond (Prunus dulcis (Mill.) D.A. Webb) skins as a potential source of bioactive polyphenols. J. Agric. Food Chem. 2007, 55, 8498–8507. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Vardakou, M.; Faulks, R.; Bisignano, C.; Martorana, M.; Smeriglio, A.; Trombetta, D. Food Matrix Effects of Polyphenol Bioaccessibility from Almond Skin during Simulated Human Digestion. Nutrients 2016, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Garrido, I.; Monagas, M.; Gómez-Cordovés, C.; Bartolomé, B. Polyphenols and Antioxidant Properties of Almond Skins: Influence of Industrial Processing. J. Food Sci. 2008, 73, C106–C115. [Google Scholar] [CrossRef] [PubMed]
- Llorach, R.; Garrido, I.; Monagas, M.; Urpi-Sarda, M.; Tulipani, S.; Bartolome, B.; Andres-Lacueva, C. Metabolomics study of human urinary metabolome modifications after intake of almond (Prunus dulcis (Mill.) D.A. Webb) skin polyphenols. J. Proteome Res. 2010, 9, 5859–5867. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, B.; Monagas, M.; Garrido, I.; Gómez-Cordovés, C.; Martín-Alvarez, P.J.; Lebrón-Aguilar, R.; Urpí-Sardà, M.; Llorach, R.; Andrés-Lacueva, C. Almond (Prunus dulcis (Mill.) D.A. Webb) polyphenols: From chemical characterization to targeted analysis of phenolic metabolites in humans. Arch. Biochem. Biophys. 2010, 501, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Blumberg, J.B. In vitro activity of almond skin polyphenols for scavenging free radicals and inducing quinone reductase. J. Agric. Food Chem. 2008, 56, 4427–4434. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Genovese, T.; Bisignano, C.; Mazzon, E.; Wickham, M.S.; Di Paola, R.; Bisignano, G.; Cuzzocrea, S. Neuroprotective effects of almond skins in experimental spinal cord injury. Clin. Nutr. 2011, 30, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bisignano, C.; Genovese, T.; Mazzon, E.; Wickham, M.S.; Paterniti, I.; Cuzzocrea, S. Natural almond skin reduced oxidative stress and inflammation in an experimental model of inflammatory bowel disease. Int. Immunopharmacol. 2011, 11, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bisignano, C.; D’Arrigo, M.; Ginestra, G.; Arena, A.; Tomaino, A.; Wickham, M.S. Antimicrobial potential of polyphenols extracted from almond skins. Lett. Appl. Microbiol. 2010, 51, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Bisignano, C.; Filocamo, A.; La Camera, E.; Zummo, S.; Fera, M.T.; Mandalari, G. Antibacterial activities of almond skins on cagA-positive and-negative clinical isolates of Helicobacter pylori. BMC Microbiol. 2013, 13, 103. [Google Scholar] [CrossRef] [PubMed]
- Arena, A.; Bisignano, C.; Stassi, G.; Mandalari, G.; Wickham, M.S.; Bisignano, G. Immunomodulatory and antiviral activity of almond skins. Immunol. Lett. 2010, 132, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Arena, A.; Bisignano, C.; Stassi, G.; Filocamo, A.; Mandalari, G. Almond Skin Inhibits HSV-2 Replication in Peripheral Blood Mononuclear Cells by Modulating the Cytokine Network. Molecules 2015, 20, 8816–8822. [Google Scholar] [CrossRef] [PubMed]
- Medici, M.A.; Sciortino, M.T.; Perri, D.; Amici, C.; Avitabile, E.; Ciotti, M.; Balestrieri, E.; De Smaele, E.; Franzoso, G.; Mastino, A. Protection by herpes simplex virus glycoprotein D against Fas-mediated apoptosis: Role of nuclear factor kappa B. J. Biol. Chem. 2003, 278, 36059–36067. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, M.T.; Medici, M.A.; Marino-Merlo, F.; Zaccaria, D.; Giuffrè-Cuculletto, M.; Venuti, A.; Grelli, S.; Mastino, A. Involvement of HVEM receptor in activation of nuclear factor kappa B by herpes simplex virus 1 glycoprotein D. Cell Microbiol. 2008, 10, 2297–2311. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.; Campadelli-Fiume, G.; Mocarski, E.; Moore, P.S.; Roizman, B.; Whitley, R.; Yamanishi, K. Human Herpesviruses Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007; pp. 1–1408. ISBN 9780521827140. [Google Scholar]
- Celum, C.; Wald, A.; Lingappa, J.R.; Magaret, A.S.; Wang, R.S.; Mugo, N.; Mujugira, A.; Baeten, J.M.; Mullins, J.I.; Hughes, J.P.; et al. Acyclovir and transmission of HIV-1 from persons infected with HIV-1 and HSV-2. N. Engl. J. Med. 2010, 5, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Kimberlin, D.W.; Whitley, R.J. Antiviral therapy of HSV-1 and -2. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Eds.; Cambridge University Press: Cambridge, UK, 2007; Chapter 64. [Google Scholar]
- Barreca, D.; Laganà, G.; Leuzzi, U.; Smeriglio, A.; Trombetta, D.; Bellocco, E. Evaluation of the nutraceutical, antioxidant and cytoprotective properties of ripe pistachio (Pistacia vera L., variety Bronte) hulls. Food Chem. 2016, 196, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Bellocco, E.; Barreca, D.; Lagana, G.; Calderaro, A.; El Lekhlifi, Z.; Chebaibi, S.; Smeriglio, A.; Trombetta, D. Cyanidin-3-O-galactoside in ripe pistachio (Pistachia vera L. variety Bronte) hulls: Identification and evaluation of its antioxidant and cytoprotective activities. J. Funct. Foods 2016, 27, 376–385. [Google Scholar] [CrossRef]
- Smeriglio, A.; Mandalari, G.; Bisignano, C.; Filocamo, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Polyphenolic content and biological properties of Avola almond (Prunus dulcis Mill. DA Webb) skin and its industrial by products. Ind. Crops Prod. 2016, 83, 283–293. [Google Scholar] [CrossRef]
- Siracusano, G.; Venuti, A.; Lombardo, D.; Mastino, A.; Esclatine, A.; Sciortino, M.T. Early activation of MyD88-mediated autophagy sustains HSV-1 replication in human monocytic THP-1 cells. Sci. Rep. 2016, 6, 31302. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, M.T.; Parisi, T.; Siracusano, G.; Mastino, A.; Taddeo, B.; Roizman, B. The virion host shutoff RNase plays a key role in blocking the activation of protein kinase R in cells infected with herpes simplex virus 1. J. Virol. 2013, 87, 3271–3276. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.W.; Dolnikowski, G.; Blumberg, J.B.; Chen, C.O. Polyphenol content and antioxidant activity of California almonds depend on cultivar and harvest year. Food Chem. 2010, 122, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Van de Perre, P.; Segondy, M.; Foulongne, V.; Ouedraogo, A.; Konate, I.; Huraux, J.M.; Mayaud, P.; Nagot, N. Herpes simplex virus and HIV-1: Deciphering viral synergy. Lancet Infect. Dis. 2008, 8, 490–497. [Google Scholar] [CrossRef]
- Paterson, I.; Anderson, E.A. Chemistry. The renaissance of natural products as drug candidates. Science 2005, 21, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.F.; Bai, L.P.; Huang, W.B.; Li, X.Z.; Zhao, S.S.; Zhong, N.S.; Jiang, Z.H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure-activity relationship analysis. Fitoterapia 2014, 93, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Thapa, M.; Kim, Y.; Desper, J.; Chang, K.O.; Hua, D.H. Synthesis and antiviral activity of substituted quercetins. Bioorg. Med. Chem. Lett. 2012, 1, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.E.; Kuster, R.M.; Yamamoto, K.A.; Salles, T.S.; Campos, R.; de Meneses, M.D.; Soares, M.R.; Ferreira, D. Quercetin and quercetin 3-O-glycosides from Bauhinia longifolia (Bong.) Steud. show anti-Mayaro virus activity. Parasites Vectors 2014, 28, 130. [Google Scholar] [CrossRef] [PubMed]
- Villaño, D.; Fernández-Pachón, M.S.; Moyá, M.L.; Troncoso, A.M.; García-Parrilla, M.C. Radical scavenging ability of polyphenolic compounds towards DPPH free radical. Talanta 2007, 71, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Khanduja, K.L.; Bhardwaj, A. Stable free radical scavenging and antiperoxidative properties of resveratrol compared in vitro with some other bioflavonoids. Indian J. Biochem. Biophys. 2003, 40, 416–422. [Google Scholar] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Bonarska-Kujawa, D.; Pruchnik, H.; Cyboran, S.; Żyłka, R.; Oszmiański, J.; Kleszczyńska, H. Biophysical Mechanism of the Protective Effect of Blue Honeysuckle (Lonicera caerulea L. var. kamtschatica Sevast.) Polyphenols Extracts Against Lipid Peroxidation of Erythrocyte and Lipid Membranes. J. Membr. Biol. 2014, 247, 611–625. [Google Scholar] [PubMed]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.P.; McCormick, T.G.; Nance, C.L.; Shearer, W.T. Epigallocatechin gallate, the main polyphenol in green tea, binds to the T-cell receptor, CD4: Potential for HIV-1 therapy. J. Allergy Clin. Immunol. 2006, 118, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, C.E.; Wen, G.Y.; Xu, W.; Jia, J.H.; Rohan, L.; Corbo, C.; Di Maggio, V.; Jenkins, E.C., Jr.; Hillier, S. Epigallocatechin gallate inactivates clinical isolates of herpes simplex virus. Antimicrob. Agents Chemother. 2008, 52, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.P.; Rybachuk, G.V.; Kousoulas, K.G. Expression of the enhanced green fluorescent protein by herpes simplex virus type 1 (HSV-1) as an in vitro or in vivo marker for virus entry and replication. J. Virol. Methods 1998, 75, 151–160. [Google Scholar] [CrossRef]
- Garner, J.A. Herpes simplex virion entry into and intracellular transport within mammalian cells. Adv. Drug Deliv. Rev. 2003, 55, 1497–1513. [Google Scholar] [CrossRef] [PubMed]
- Roizman, B.; Knipe, D.M.; Whitley, R. Herpes Simplex Viruses. In Fields Virology, 6th ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2013; Volume 2, pp. 1823–1897. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | (mg of Skin *) | mgGAE/100 g FW |
|---|---|---|
| NS | 0.032 ± 0.001 | 4228.52 ± 265.19 |
| BS | 0.117 ± 0.003 | 3841.16 ± 18.56 |
| Peak No. | Compound | * RT (min) | λmax (nm) |
|---|---|---|---|
| 1 | Protocatechuic acid | 13.081 | 259; 294 |
| 2 | p-Hydroxybenzoic acid | 20.442 | 255 |
| 3 | Catechin | 23.616 | 278 |
| 4 | Chlorogenic acid | 26.510 | 296; 326 |
| 5 | Vanillic acid | 26.892 | 261; 292 |
| 6 | Epicatechin | 32.561 | 278 |
| 7 | trans-p-Coumaric acid | 38.015 | 310 |
| 8 | Eryodictiol-7-O-glucoside | 45.933 | 284 |
| 9 | Quercetin-3-O-rutinoside | 47.434 | 256; 354 |
| 10 | Quercetin-3-O-galactoside | 47.757 | 256; 354 |
| 11 | Quercetin-3-O-glucoside | 48.892 | 256; 354 |
| 12 | Kaempferol-3-O-rutinoside | 53.395 | 265; 347 |
| 13 | Kaempferol-3-O-glucoside | 54.279 | 265; 347 |
| 14 | Naringenin-7-O-glucoside | 54.555 | 284; 338 |
| 15 | Isorhamnetin-3-O-rutinoside | 54.937 | 256; 354 |
| 16 | Isorhamnetin-3-O-glucoside | 56.650 | 254; 354 |
| 17 | Eriodictyol | 66.052 | 288 |
| 18 | Quercetin | 70.367 | 256; 372 |
| 19 | Naringenin | 73.018 | 288 |
| 20 | Kaempferol | 88.516 | 264; 366 |
| 21 | Isorhamnetin | 93.060 | 254; 370 |
| Compound | NS | BS |
|---|---|---|
| Hydroxybenzoic acids | ||
| Protocatechuic acid | 3862.97 ± 124.52 | 878.24 ± 22.65 |
| p-Hydroxybenzoic acid | 7094.27 ± 224.10 | 696.21 ± 32.41 |
| Vanillic acid | 5652.08 ± 236.54 | 709.027 ± 11.35 |
| Hydroxycinnamic acids | ||
| Chlorogenic acid | 1144.54 ± 84.25 | 375.41 ± 8.54 |
| trans-p-Coumaric acid | 791.62 ± 22.45 | 142.19 ± 5.24 |
| Flavanones | ||
| Eriodictyol | 2777.23 ± 78.65 | 364.8 ± 12.54 |
| Eryodictiol-7-O-glucoside | 57.9 ± 1.26 | 8.33 ± 0.221 |
| Naringenin | 5802.51 ± 185.44 | 474.54 ± 20.85 |
| Naringenin-7-O-glucoside | 34,020.11 ± 654.22 | 2148.22 ± 62.35 |
| Flavonols | ||
| Kaempferol-3-O-rutinoside | 8062.45 ± 261.33 | 1037.64 ± 52.14 |
| Kaempferol-3-O-glucoside | 29,530.44 ± 854.22 | 522.84 ± 21.47 |
| Isorhamnetin-3-O-glucoside | 16,597.37 ± 546.31 | 908.86 ± 18.95 |
| Quercetin | 3474.03 ± 105.62 | 2035.8 ± 68.54 |
| Kaempferol | 5573.6 ± 88.65 | 998.84 ± 22.64 |
| Isorhamnetin | 3513.47 ± 112.35 | 230.4 ± 9.87 |
| Quercetin-3-O-rutinoside | 1067 ± 55.62 | 299.55 ± 8.03 |
| Quercetin-3-O-galactoside | 1400.8 ± 37.45 | 123.56 ± 4.42 |
| Quercetin-3-O-glucoside | 1219.53 ± 24.89 | 108.67 ± 2.89 |
| Isorhamnetin-3-O-rutinoside | 17,620.45 ± 495.65 | 1526.53 ± 23.98 |
| Flavanols | ||
| Epicatechin | 26,948.72 ± 554.25 | 1900.8 ± 62.74 |
| Catechin | 47,998.32 ± 956.54 | 4210.91 ± 88.54 |
| Total amount | 224,209.41 | 19,701.36 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisignano, C.; Mandalari, G.; Smeriglio, A.; Trombetta, D.; Pizzo, M.M.; Pennisi, R.; Sciortino, M.T. Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell. Viruses 2017, 9, 178. https://doi.org/10.3390/v9070178
Bisignano C, Mandalari G, Smeriglio A, Trombetta D, Pizzo MM, Pennisi R, Sciortino MT. Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell. Viruses. 2017; 9(7):178. https://doi.org/10.3390/v9070178
Chicago/Turabian StyleBisignano, Carlo, Giuseppina Mandalari, Antonella Smeriglio, Domenico Trombetta, Maria Musarra Pizzo, Rosamaria Pennisi, and Maria Teresa Sciortino. 2017. "Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell" Viruses 9, no. 7: 178. https://doi.org/10.3390/v9070178
APA StyleBisignano, C., Mandalari, G., Smeriglio, A., Trombetta, D., Pizzo, M. M., Pennisi, R., & Sciortino, M. T. (2017). Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell. Viruses, 9(7), 178. https://doi.org/10.3390/v9070178