
Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
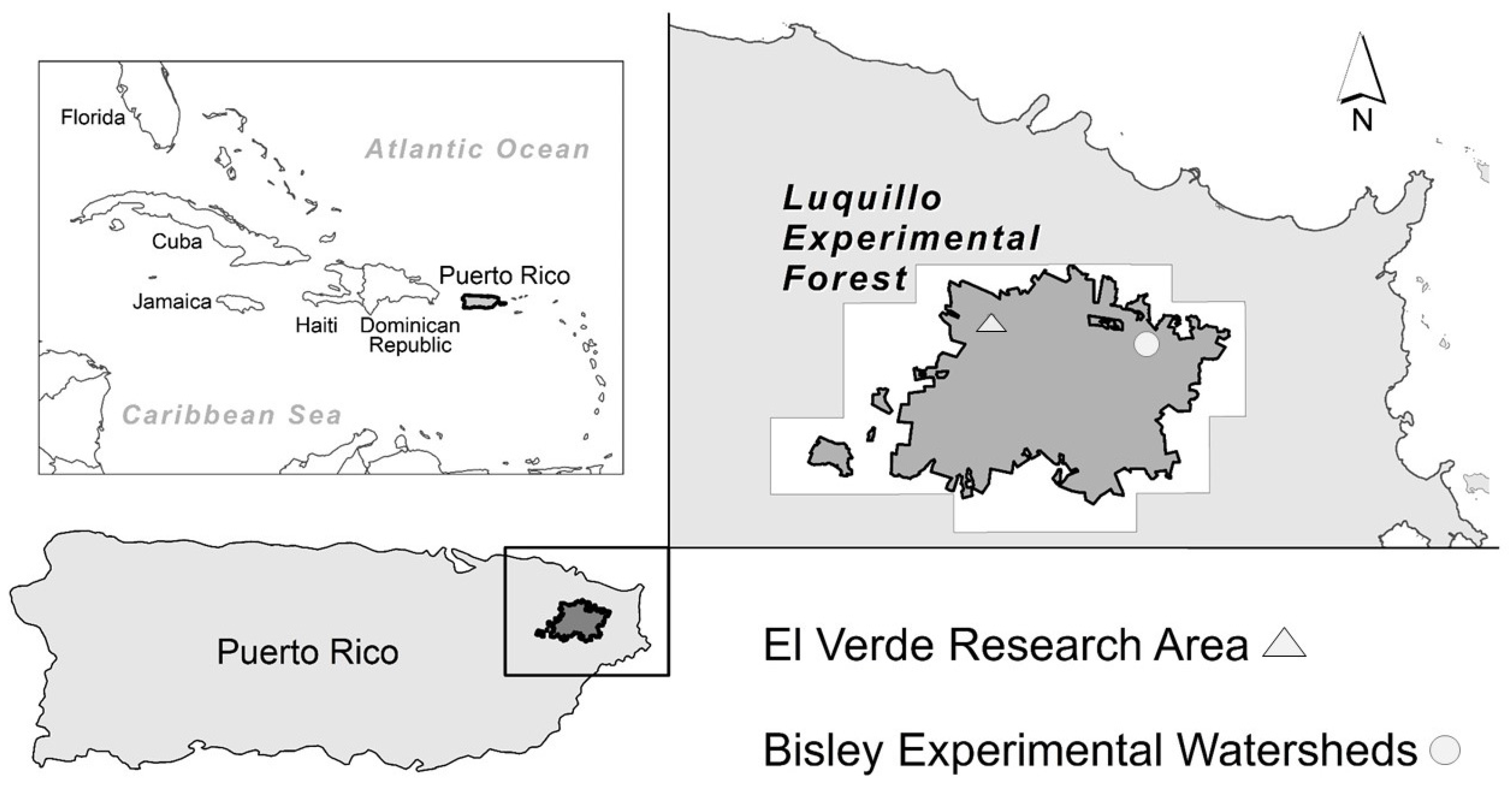
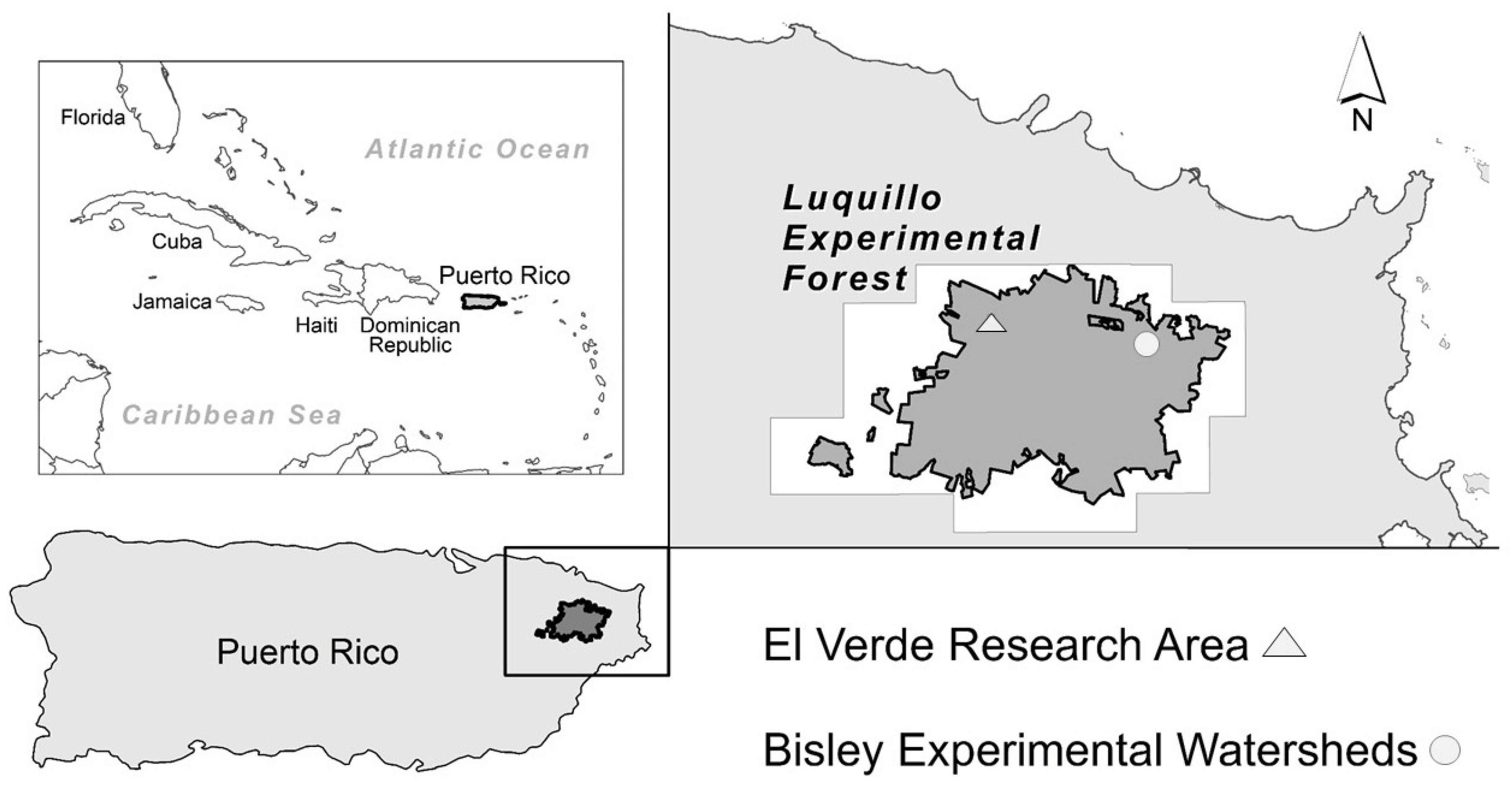
2.1. Study Site
2.1.1. Permanent Plots
2.1.2. Data Analyses
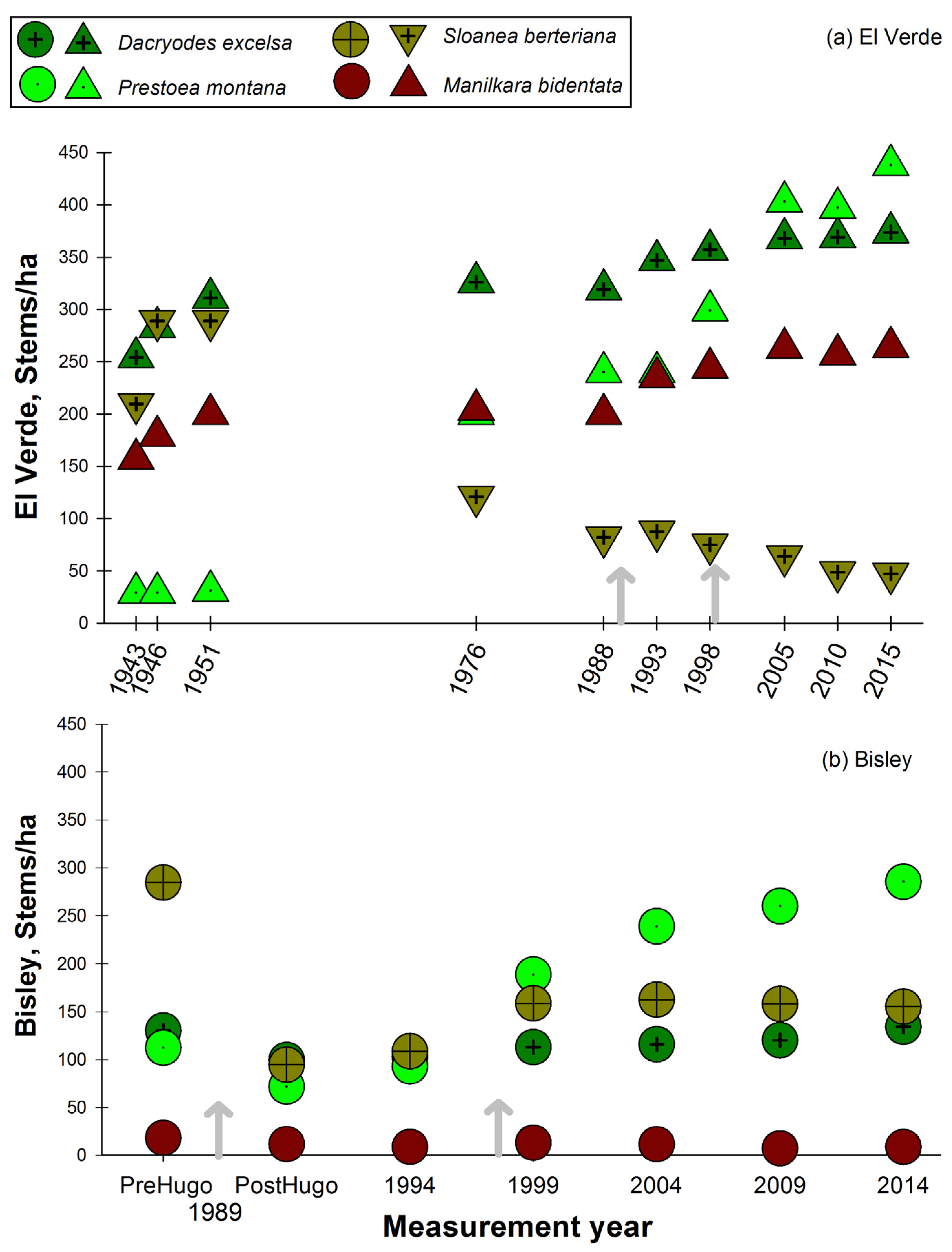
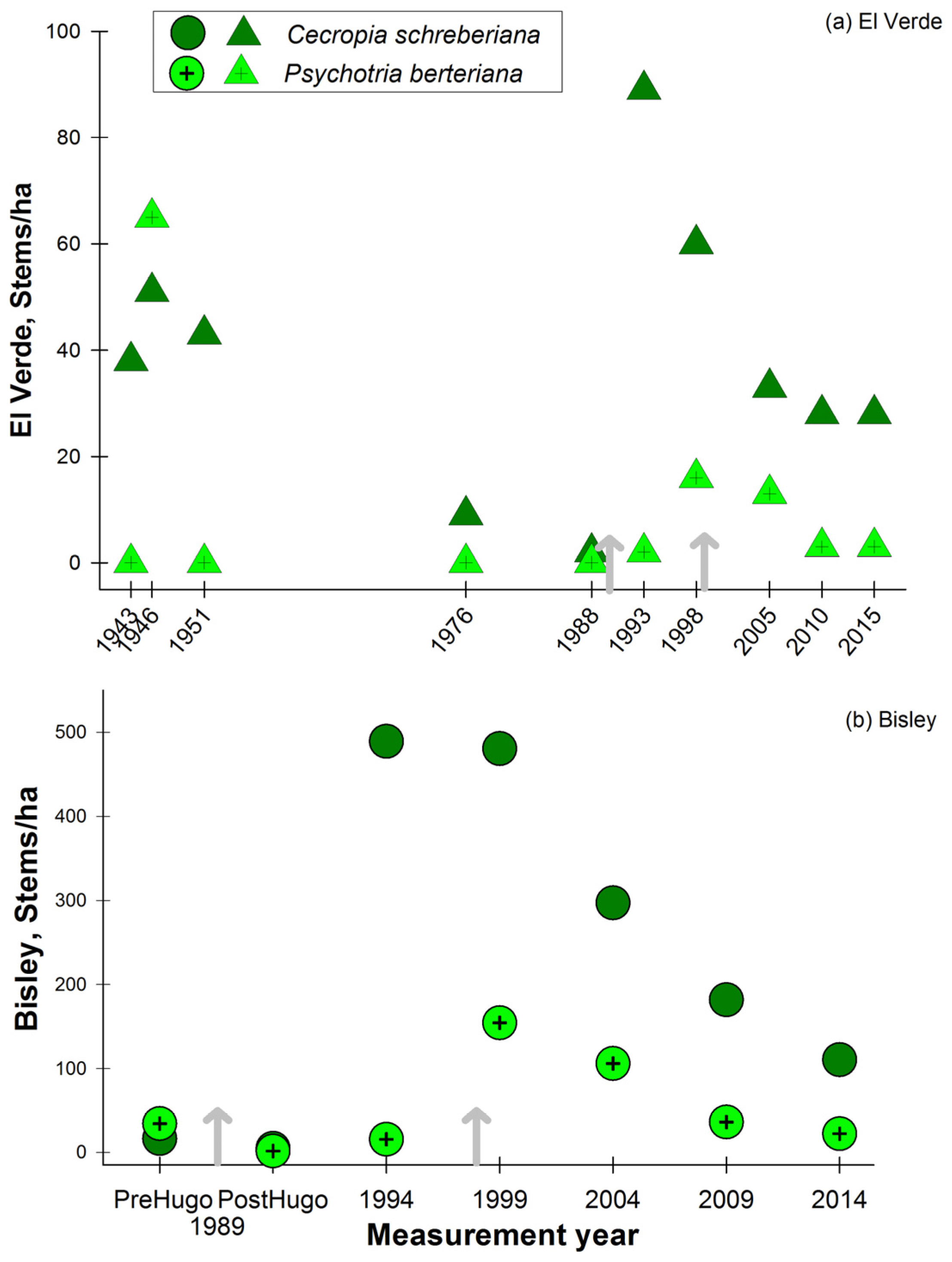
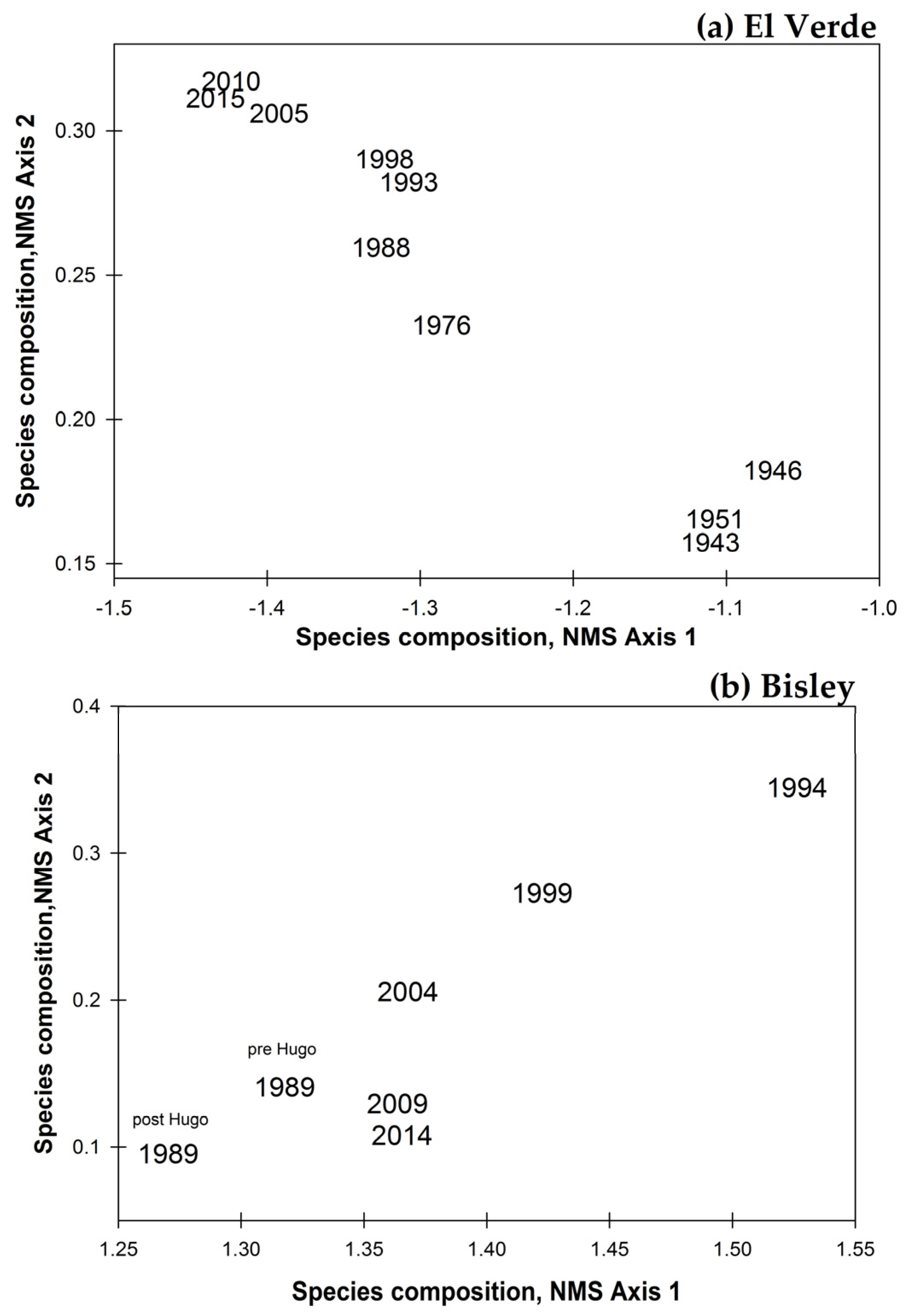
3. Results
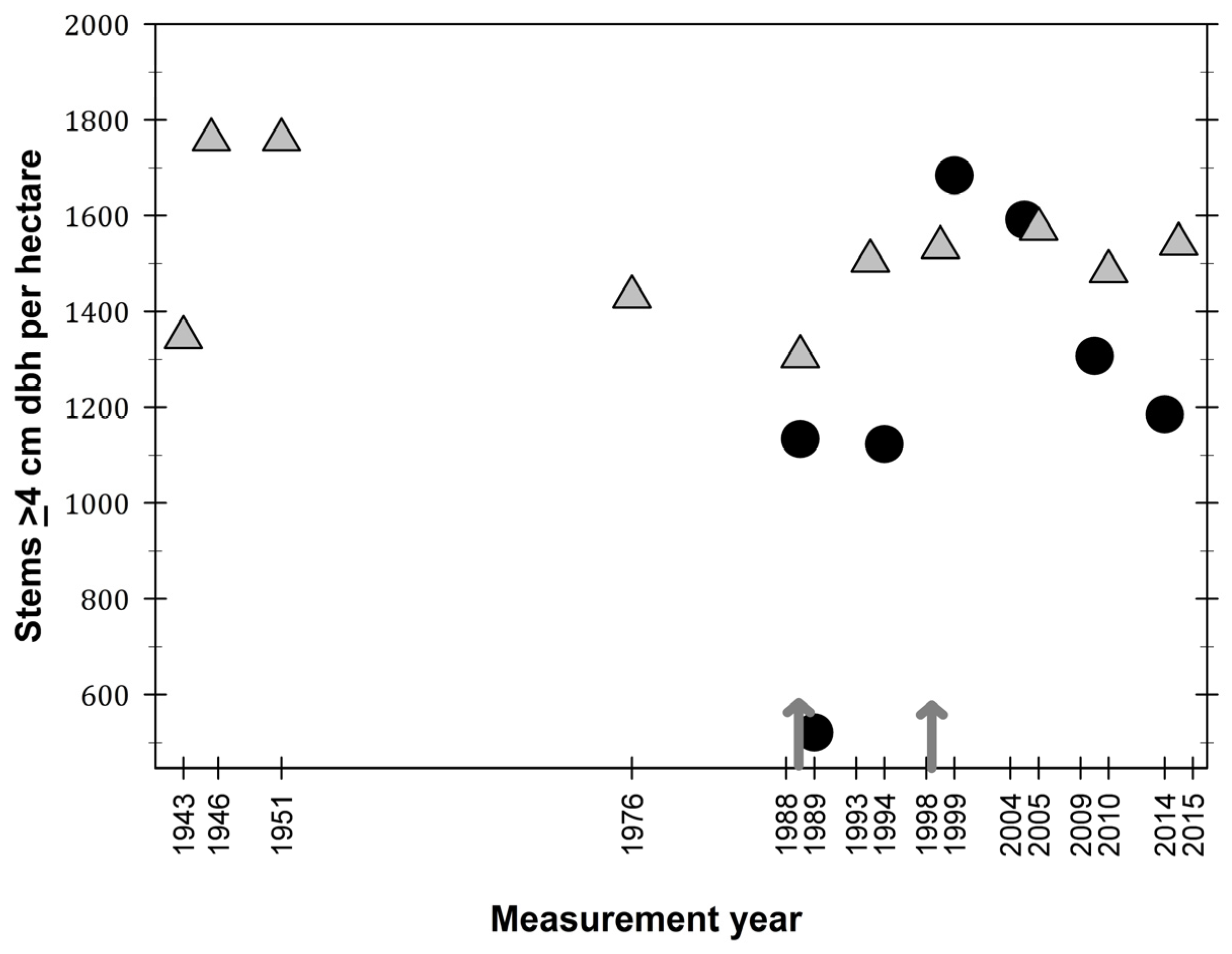
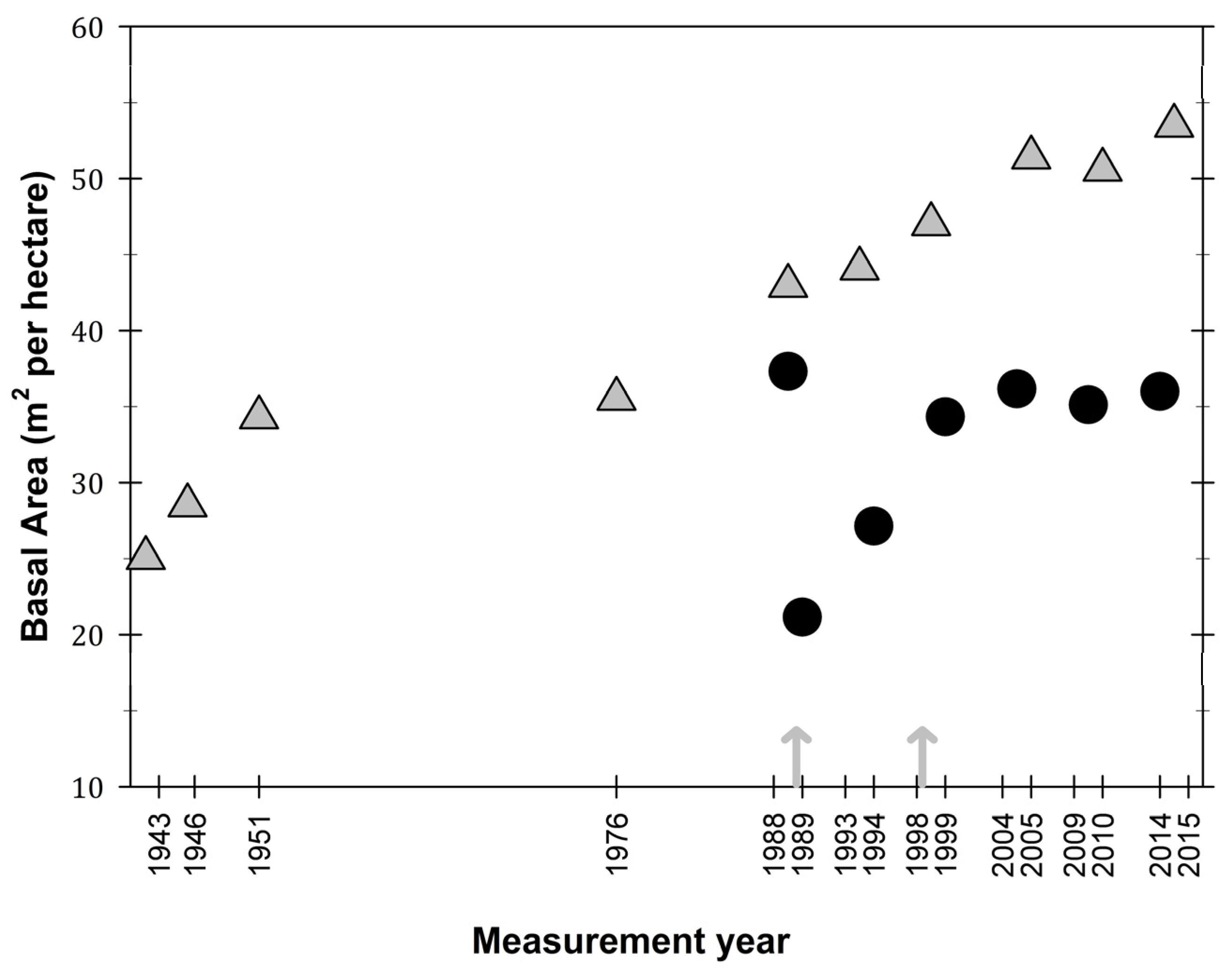
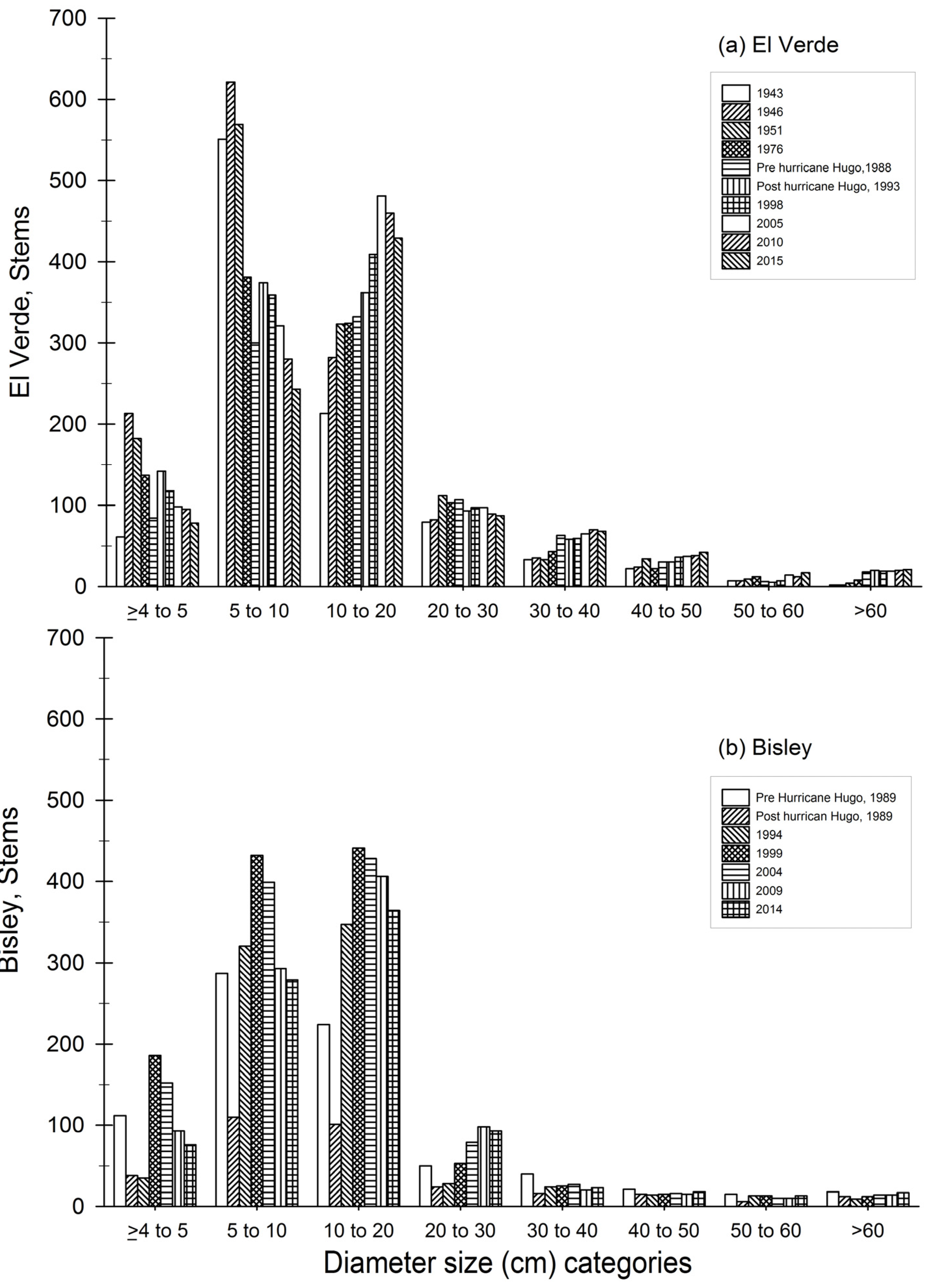
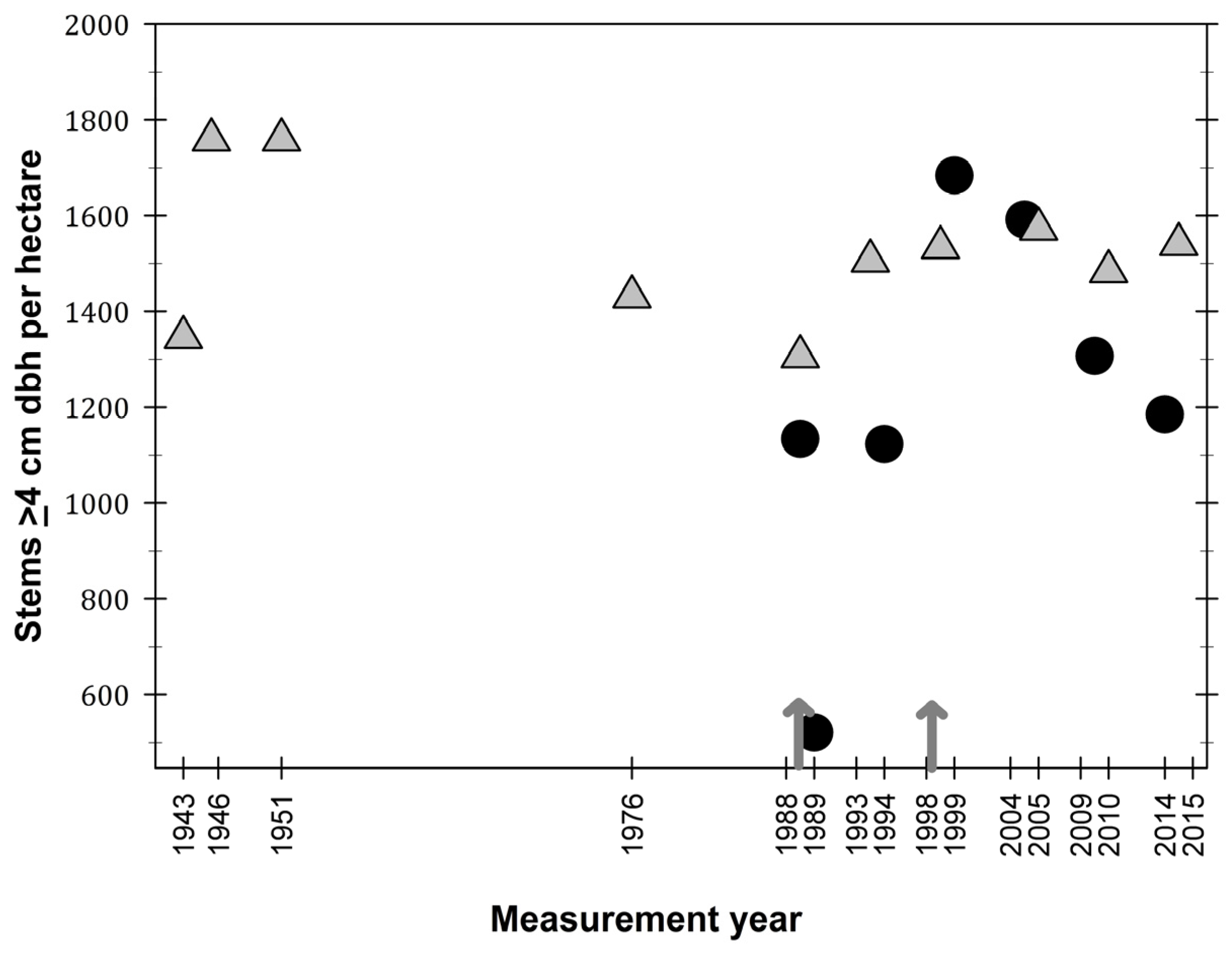
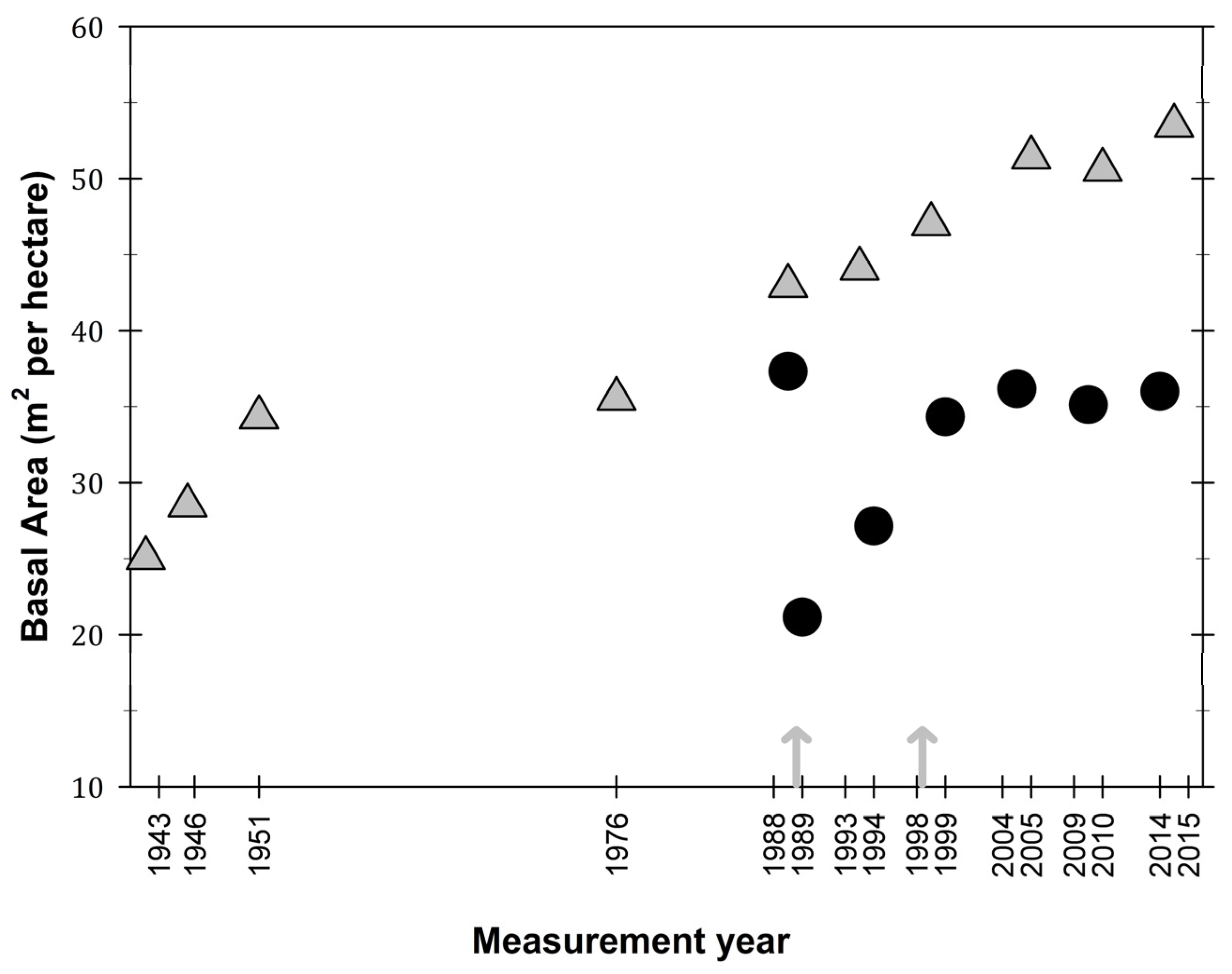
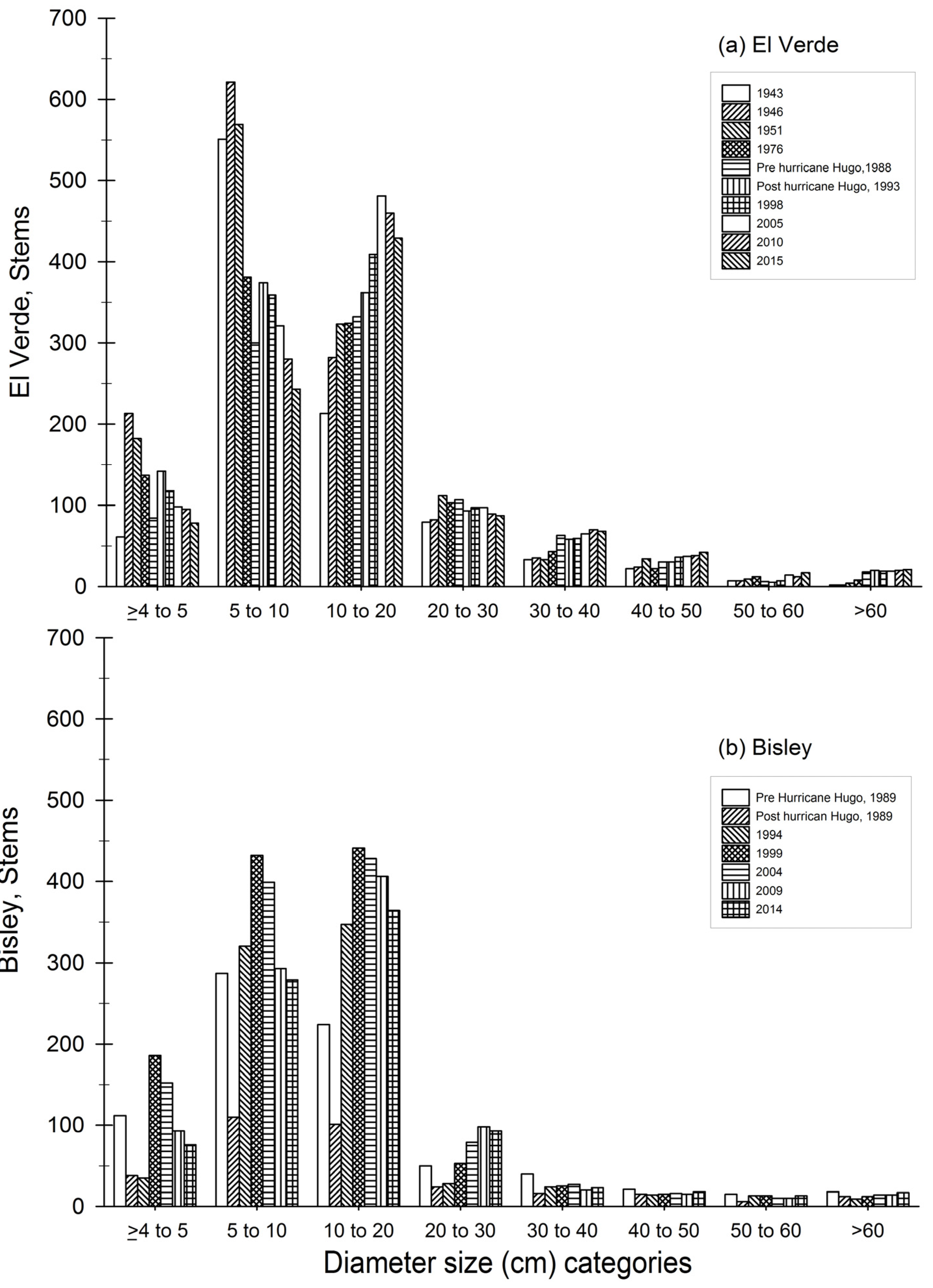
3.1. Observations on Structural Characteristics
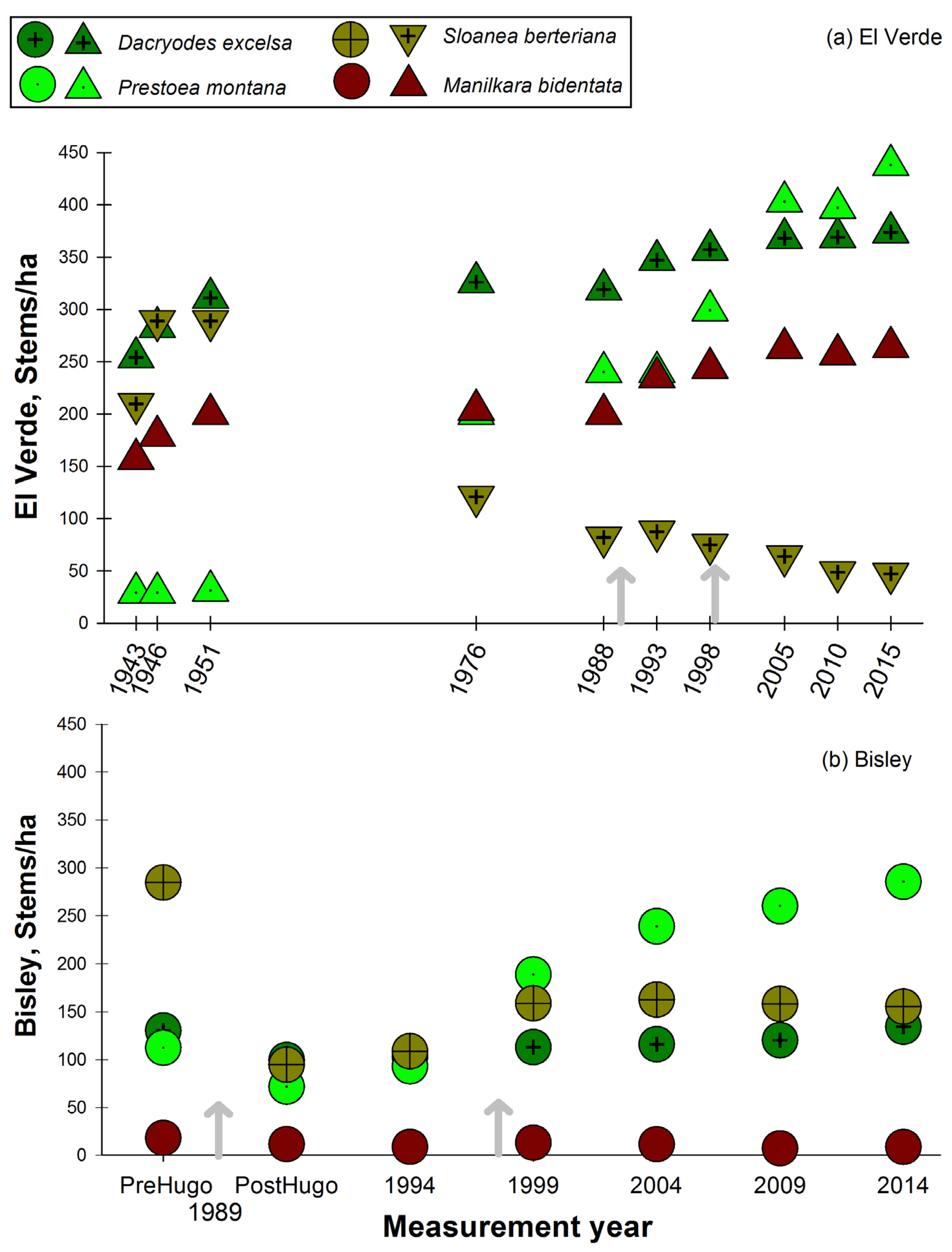
3.2. Observations on Species Abundances and Plot Species Composition
4. Discussion
Acknowledgments
Conflicts of Interest
References
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Walker, L.R. Summary of the effects of Caribbean hurricanes on vegetation. Biotropica 1991, 23, 442–447. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Willig, M.R.; Walker, L.R.; Silver, W.L. Introduction: Disturbance and Caribbean ecosystems. Biotropica 1996, 28, 414–423. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- López-Marrero, T.; Heartsill-Scalley, T. Get up, stand up: Environmental situation, threats, and opportunities in the insular Caribbean. Caribb. Stud. 2012, 40, 3–14. [Google Scholar] [CrossRef]
- Lugo, A.E.; Helmer, E.H.; Santiago Valentín, E. Caribbean landscapes and their biodiversity. Interciencia 2012, 37, 705–710. [Google Scholar]
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2012, 33, 368–398. [Google Scholar] [CrossRef]
- Chai, S.L.; Healey, J.R.; Tanner, E.V. Evaluation of forest recovery over time and space using permanent plots monitored over 30 years in a Jamaican montane rain forest. PLoS ONE 2012, 7, e48859. [Google Scholar] [CrossRef] [PubMed]
- Luke, D.; McLaren, K.; Wilson, B. Modeling hurricane exposure in a Caribbean lower montane tropical wet forest: The Effects of frequent, intermediate disturbances and topography on forest structural dynamics and composition. Ecosystems 2016, 19, 1178–1195. [Google Scholar] [CrossRef]
- Crow, T.R. A rainforest chronicle: A 30-year record of change in structure and composition at El Verde, Puerto Rico. Biotropica 1980, 12, 42–55. [Google Scholar] [CrossRef]
- Johnston, M.H. Successional Change and Species/Site Relationships in a Puerto Rican Tropical Forest. Ph.D. Thesis, State University of New York, New York, NY, USA, 1991; p. 253. [Google Scholar]
- Drew, A.P.; Boley, J.D.; Zhao, Y.; Johnston, M.H.; Wadsworth, F.H. Sixty-two years of change in subtropical wet forest structure and composition at El Verde, Puerto Rico. Interciencia 2009, 34, 34–40. [Google Scholar]
- Heartsill, S.; Tamara, S.; Frederick, N.; Lugo, A.E.; Moya, S.; Estrada Ruiz, C.R. Changes in structure, composition, and nutrients during 15 years of hurricane-induced succession in a subtropical wet forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Lugo, A.E.; Frangi, J.L. Long-term response of Caribbean palm forests to hurricanes. Caribb. Nat. 2016, 157–175. [Google Scholar]
- Walker, L.R.; Zarin, D.J.; Fetcher, N.; Myster, R.W.; Johnson, A.H. Ecosystem development and plant succession on landslides in the Caribbean. Biotropica 1996, 28, 566–576. [Google Scholar] [CrossRef]
- Coll, M.; Fonseca, A.C.; Cortés, J. The mangrove and others vegetation associations in de Gandoca lagoon, Limón, Costa Rica. Rev. Biol. Trop. 2001, 49, 321–329. [Google Scholar] [PubMed]
- Heartsill Scalley, T.; Scatena, F.N.; Estrada, C.; McDowell, W.H.; Lugo, A.E. Disturbance and long-term patterns of rainfall and throughfall nutrient fluxes in a subtropical wet forest in Puerto Rico. J. Hydrol. 2007, 333, 472–485. [Google Scholar] [CrossRef]
- Gonzalez, G.; Waide, R.B.; Willig, M.R. Advancements in the understanding of spatiotemporal gradients in tropical landscapes: A Luquillo focus and global perspective. Ecol. Bull. 2013, 54, 245–250. [Google Scholar]
- Thomas, S.C.; Martin, A.R.; Mycroft, E.E. Tropical trees in a wind-exposed island ecosystem: Height-diameter allometry and size at onset of maturity. J. Ecol. 2015, 103, 594–605. [Google Scholar] [CrossRef]
- Brokaw, N.; Zimmerman, J.K.; Willig, M.; Camilo, G.; Covich, A.; Crowl, T.; Fetcher, N.; Haines, B.; Lodge, J.; Lugo, A.E.; et al. A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response; Brokaw, N., Crowl, T.A., Lugo, A.E., McDowell, W.H., Scatena, F.N., Waide, R.B., Willig, M.R., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 201–271. [Google Scholar]
- Taylor, M.A.; Stephenson, T.S.; Chen, A.A.; Stephenson, K.A. Climate change and the Caribbean: Review and response. Caribb. Stud. 2012, 40, 169–200. [Google Scholar] [CrossRef]
- Beard, J.S. The Natural Vegetation of the Windward and Leeward Islands; Clarendon Press: Oxford, UK, 1949; p. 192. [Google Scholar]
- Alvarez Ruiz, M. Effects of Human Activities on Stand Structure and Composition, and Genetic Diversity of Dacryodes Excelsa Vahl (Tabonuco); University of Puerto Rico: San Juan, Puerto Rico, 2002; p. 266. [Google Scholar]
- Lugo, A.E.; Scatena, F.N. Ecosystem-level properties of the Luquillo Experimental Forest with emphasis on the Tabonuco Forest. In Tropical Forests: Management and Ecology; Springer: New York, NY, USA, 1995; pp. 59–108. [Google Scholar]
- Basnet, K.; Scatena, F.N.; Likens, G.E.; Lugo, A.E. Ecological consequences of root grafting in Tabonuco (Dacryodes excelsa) trees in the Luquillo Experimental Forest, Puerto Rico. Biotropica 1993, 25, 28–35. [Google Scholar] [CrossRef]
- Weaver, P.L. Sloanea Berteriana Choisy Motillo. Elaeocarpaceae. Elaeocarpus Family; USDA Forest Service, International Institute of Tropical Forestry: Rio Piedras, Puerto Rico, 1997; p. 7.
- Lugo, A.E.; Francis, J.K.; Frangi, J.L. Prestoea Montana (R. Graham) Nichols. Sierra Palm. Palmaceae. Palm Family; US Department of Agriculture, Forest Service, International Institute of Tropical Forestry: Rio Piedras, Puerto Rico, 1998; p. 9.
- Weaver, P.L. Ecological observations on Magnolia splendens urban in the Luquillo mountains of Puerto Rico. Caribb. J. Sci. 1987, 23, 340–351. [Google Scholar]
- Garcia-Montiel, D.; Scatena, F.N. The effect of human activity on the structure and composition of a tropical forest in Puerto Rico. Forest Ecol. Manag. 1994, 63, 57–78. [Google Scholar] [CrossRef]
- Scatena, F.N. An Introduction to the Physiography and History of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico; Gen. Tech. Rep. SO-72. U.S.D.A; Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989; p. 22.
- Scatena, F.N.; Moya, S.; Estrada, C.; Chinea, J.D. The first five year in the reorganization of aboveground biomass and nutrient use following Hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica 1996, 28, 424–440. [Google Scholar] [CrossRef]
- Ostertag, R.; Silver, W.L.; Lugo, A.E. Factors affecting mortality and resistance to damage following hurricanes in a rehabilitated subtropical moist forest. Biotropica 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Larsen, M.C. Analysis of 20th century rainfall and streamflow to characterize drought and water resources in Puerto Rico. Phys. Geogr. 2000, 21, 494–521. [Google Scholar]
- Beard, K.H.; Vogt, K.A.; Vogt, D.J.; Scatena, F.N.; Covich, A.P.; Sigurdardottir, R.; Siccama, T.G.; Crowl, T.A. Structural and functional responses of a subtropical forest to 10 year of hurricanes and droughts. Ecol. Monogr. 2005, 75, 345–361. [Google Scholar] [CrossRef]
- Lugo, A.E.; Zimmerman, J.K. Ecological Life Histories. In Tropical Tree Seed Manual; Vozzo, J.A., Ed.; USDA Forest Service: Washington, DC, USA, 2002; pp. 191–213. [Google Scholar]
- Shiels, A.B.; Zimmerman, J.K.; García-Montiel, D.C.; Jonckheere, I.; Holm, J.; Horton, D.; Brokaw, N. Plant responses to simulated hurricane impacts in a subtropical wet forest, Puerto Rico. J. Ecol. 2010, 98, 659–673. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Murphy, L.; Sabat, A.M.; Fetcher, N.; Haines, B.L. Natural disturbance and human land use as determinants of tropical forest dynamics: Results from a forest simulator. Ecol. Monogr. 2009, 79, 423–443. [Google Scholar] [CrossRef]
- Johnson, D.E. Applied Multivariate Methods for Data Analysts; Duxbury Press: Pacific Grove, CA, USA, 1998; p. 567. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MJM Software Design: Gleneden Beach, OR, USA, 2002; p. 300. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. In Multivariate Analysis of Ecological Data; Version 5.1; Mjm Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Lugo, A.E. Effects and outcomes of Caribbean hurricanes in a climate change scenario. Sci. Total Environ. 2000, 262, 243–251. [Google Scholar] [CrossRef]
- Boose, E.R.; Foster, D.R.; Fluet, M. Hurricane impacts to tropical and temperate forest landscapes. Ecol. Monogr. 1994, 64, 369–400. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Damage and responsiveness of Jamaican montane forest tree species after disturbance by a hurricane. Ecology 1995, 76, 2562–2580. [Google Scholar] [CrossRef]
- Lodge, D.J.; Cantrell, S. Fungal communities in wet tropical forests: Variation in time and space. Can. J. Bot. 1995, 73, 1391–1398. [Google Scholar] [CrossRef]
- Lodge, D.J. Microorganisms. In The Food Wed of a Tropical Forest; Reagan, D.P., Waide, R.B., Eds.; University of Chicago press: Chicago, IL, USA, 1996; pp. 53–108. [Google Scholar]
- Yap, S.L.; Davies, S.J.; Condit, R. Dynamic response of a Philippine dipterocarp forest to typhoon disturbance. J. Veg. Sci. 2016, 27, 133–143. [Google Scholar] [CrossRef]
- Murphy, H.T.; Metcalfe, D.J.; Bradford, M.G.; Ford, A.J. Community divergence in a tropical forest following a severe cyclone. Austral Ecol. 2014, 39, 696–709. [Google Scholar] [CrossRef]
- Foster, D.R.; Knight, D.H.; Franklin, J.F. Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1988, 1, 497–510. [Google Scholar] [CrossRef]
- Willig, M.R.; Bloch, C.P.; Covich, A.P.; Hall, C.A.S.; Lodge, D.J.; Lugo, A.E.; Silver, W.L.; Waide, R.B.; Walker, L.R.; Zimmerman, J.K. Long-term research in the Luquillo Mountains, synthesis and foundations for the future. In A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response; Brokaw, N., Crowl, T.A., Lugo, A.E., Mcdowell, W.H., Scatena, F.N., Waide, R.B., Willig, M.R., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 361–441. [Google Scholar]
- Imbert, D.; Labbe, P.; Rousteau, A. Hurricane damage and forest structure in Guadeloupe, French West Indies. J. Trop. Ecol. 1996, 12, 663–680. [Google Scholar] [CrossRef]
- Royo, A.A.; Scalley, T.H.; Moya, S.; Scatena, F.N. Non-arborescent vegetation trajectories following repeated hurricane disturbance: Ephemeral versus enduring responses. Ecosphere 2011, 2, 77. [Google Scholar] [CrossRef]



© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heartsill Scalley, T. Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains. Forests 2017, 8, 204. https://doi.org/10.3390/f8060204
Heartsill Scalley T. Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains. Forests. 2017; 8(6):204. https://doi.org/10.3390/f8060204
Chicago/Turabian StyleHeartsill Scalley, Tamara. 2017. "Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains" Forests 8, no. 6: 204. https://doi.org/10.3390/f8060204
APA StyleHeartsill Scalley, T. (2017). Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains. Forests, 8(6), 204. https://doi.org/10.3390/f8060204