
Anti-Ischemic Effect of Leptin in the Isolated Rat Heart Subjected to Global Ischemia-Reperfusion: Role of Cardiac-Specific miRNAs


 ,
,
Abstract
:1. Introduction
2. Results
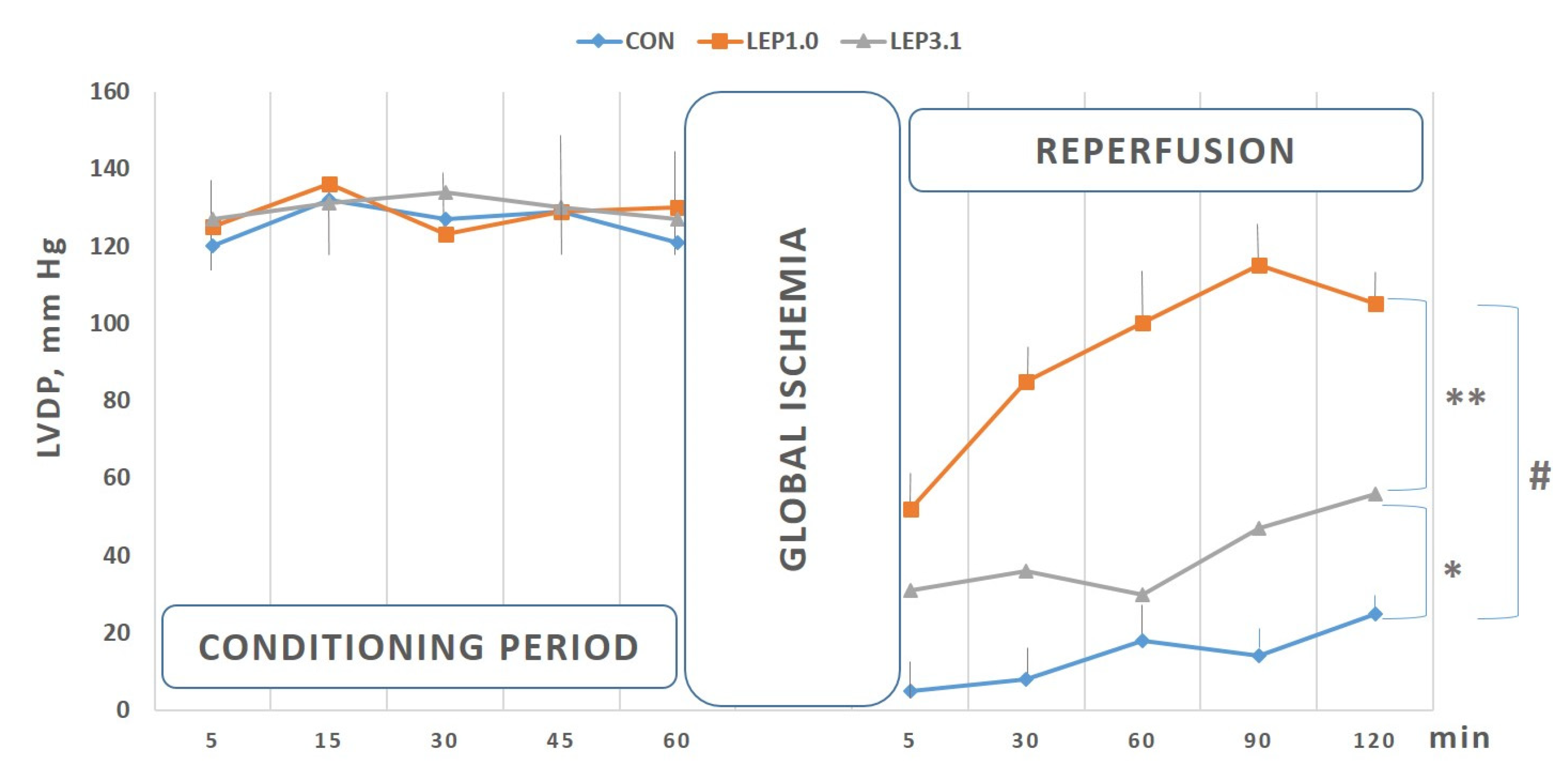
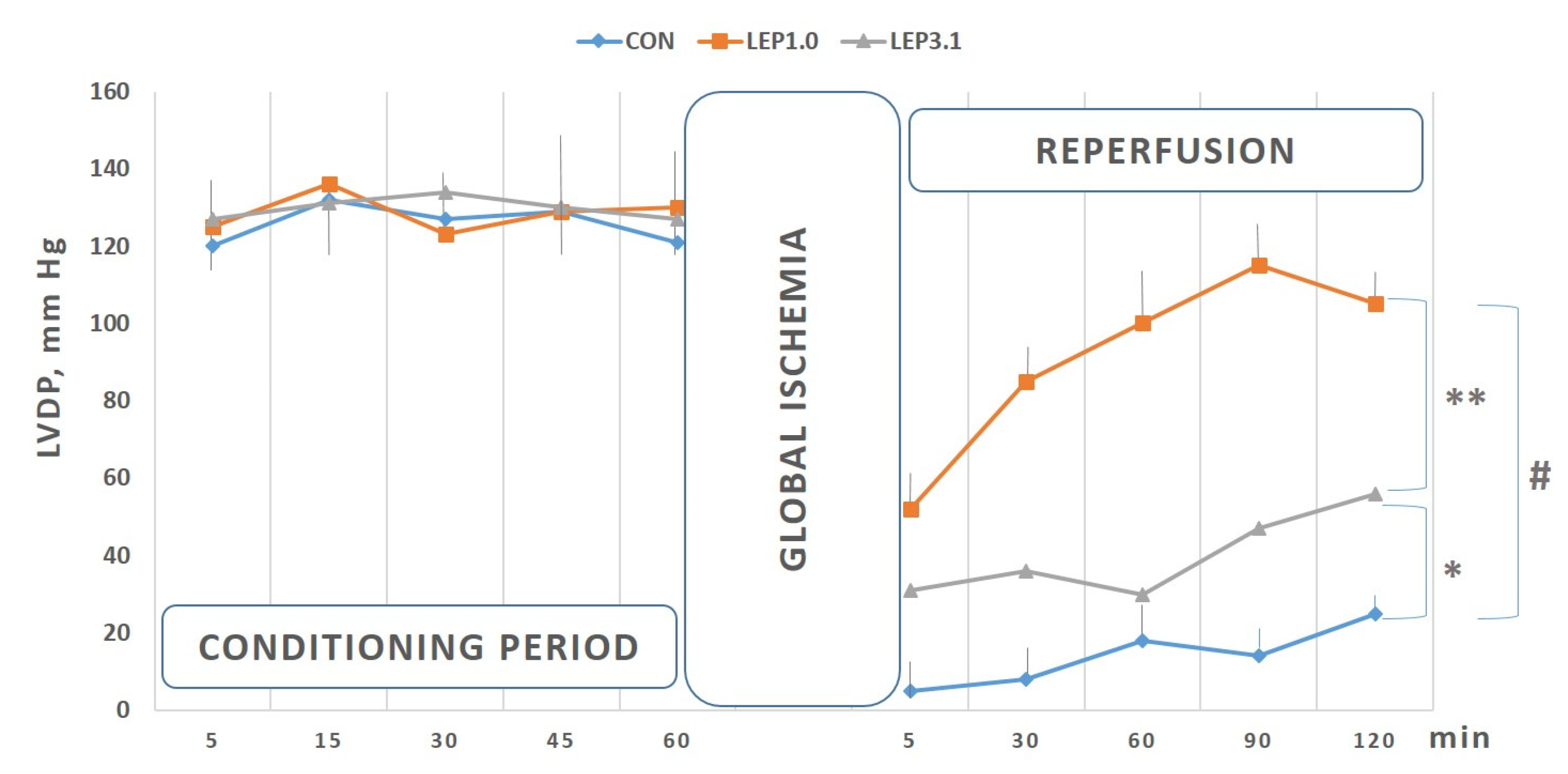
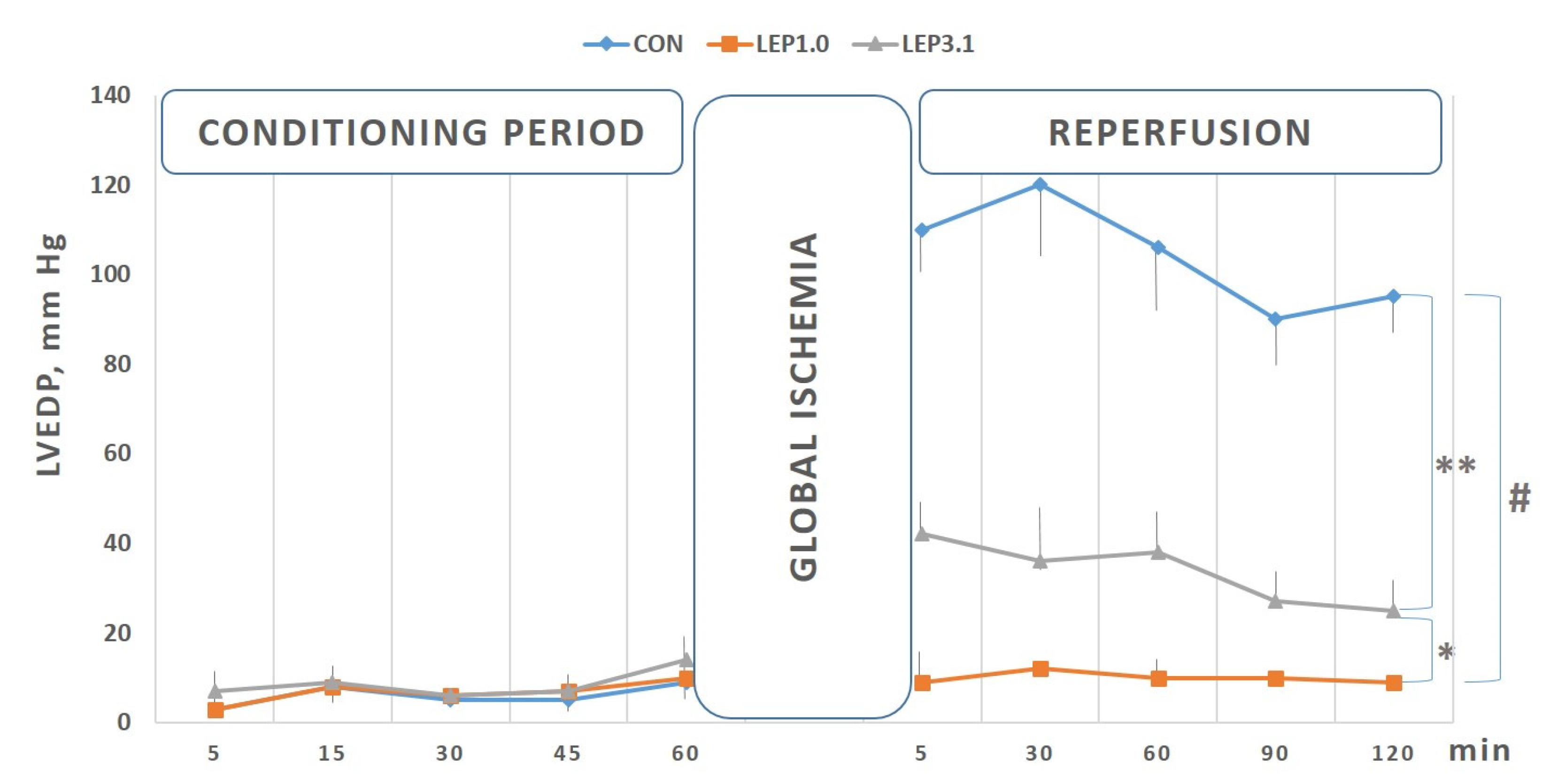
2.1. Left Ventricular Developed Pressure (LVDP) and Left Ventricular End-diastolic Pressure (LVEDP)
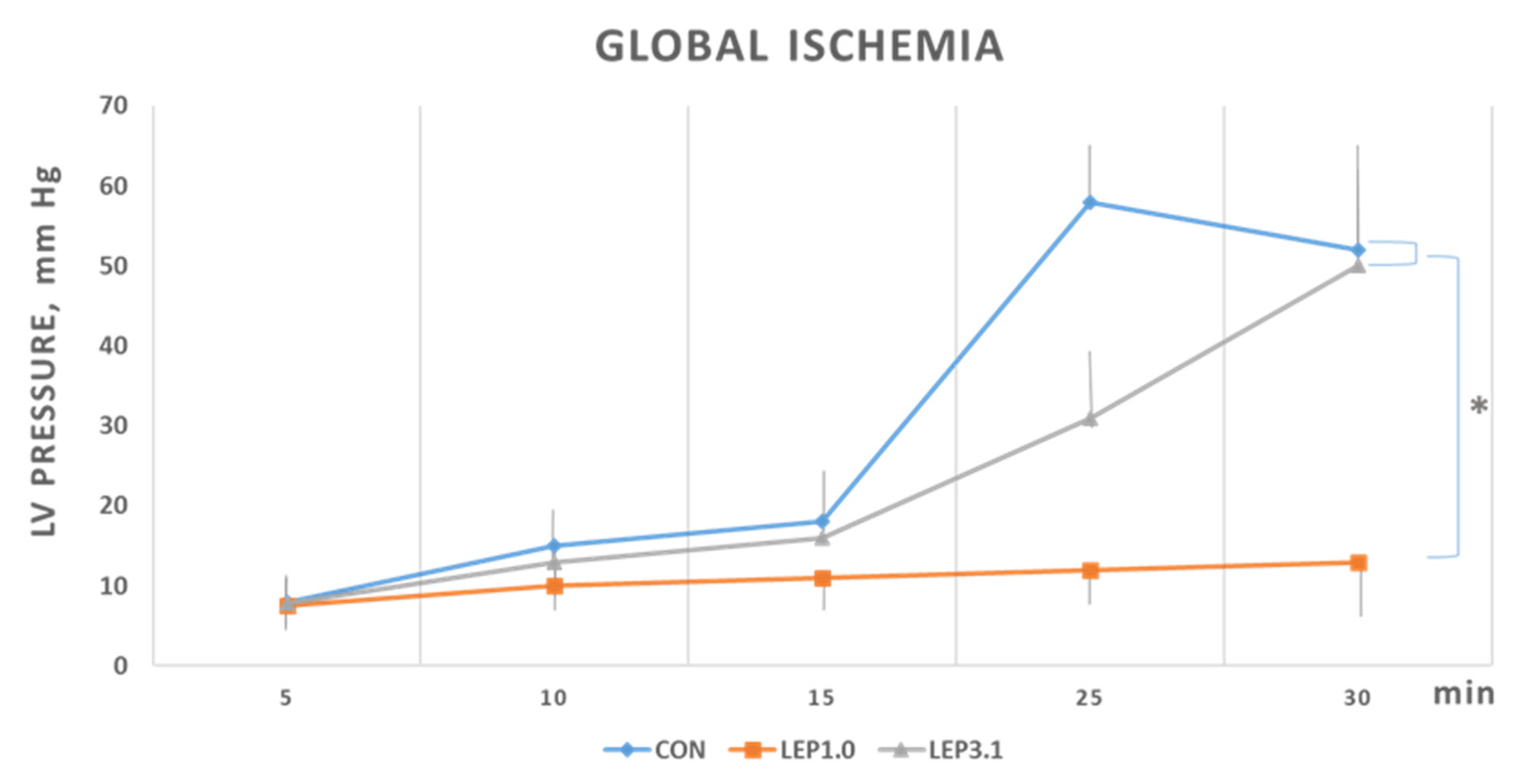
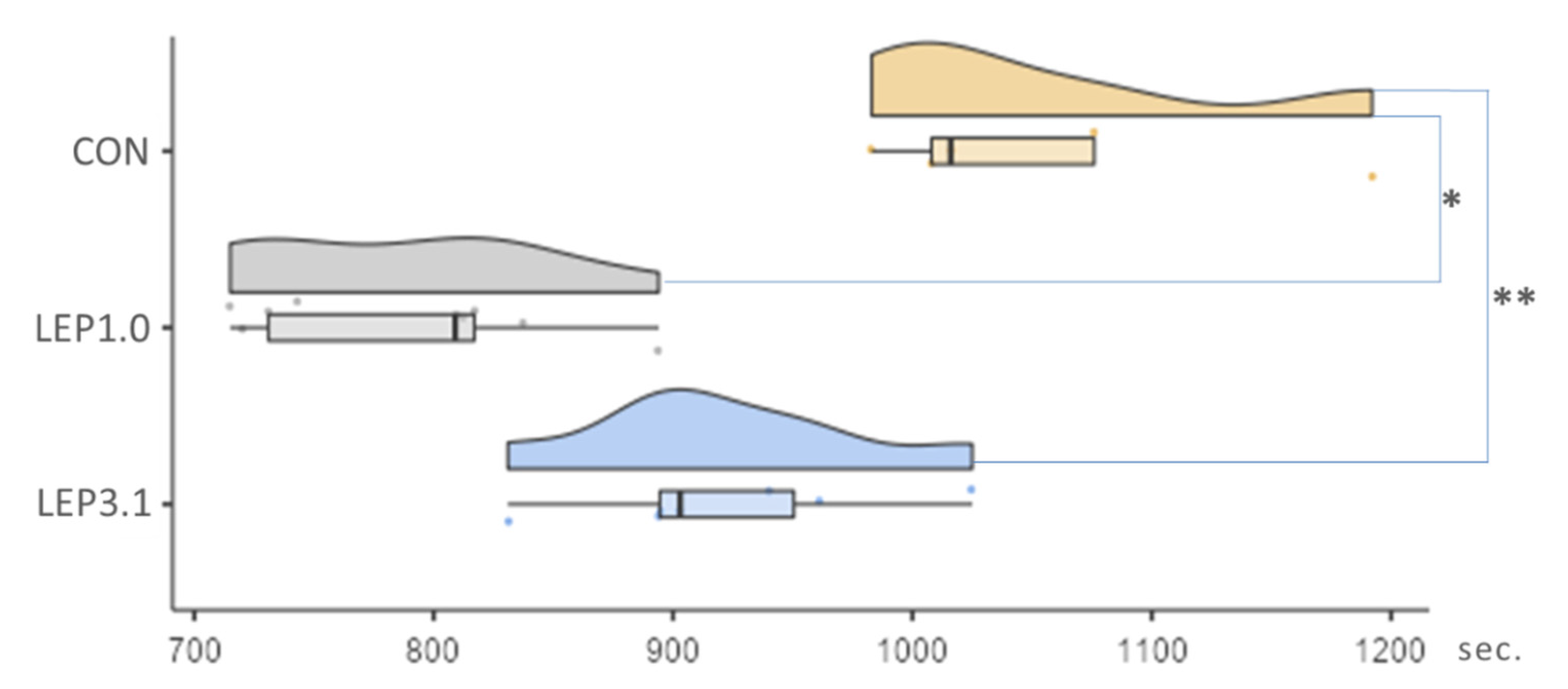
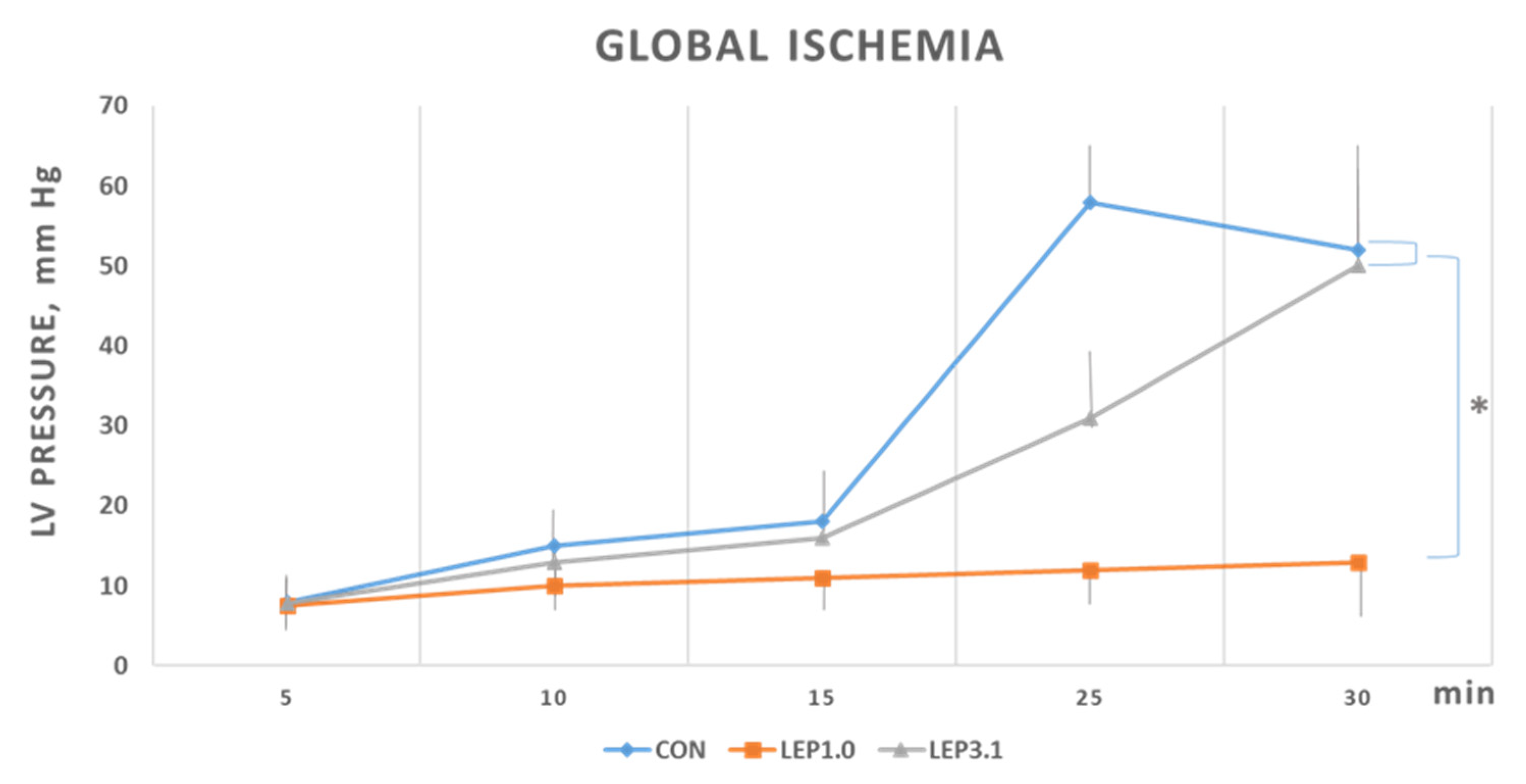
2.2. Ischemic Contracture
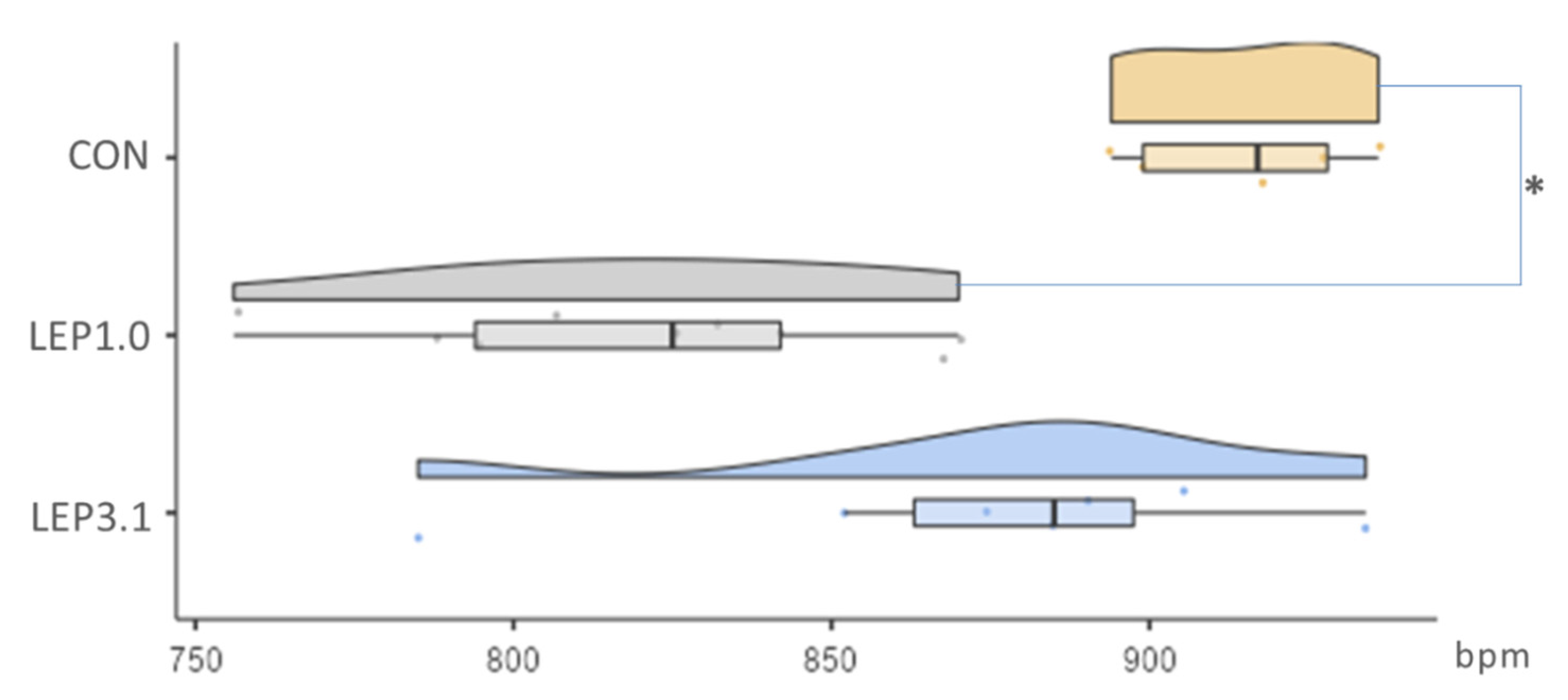
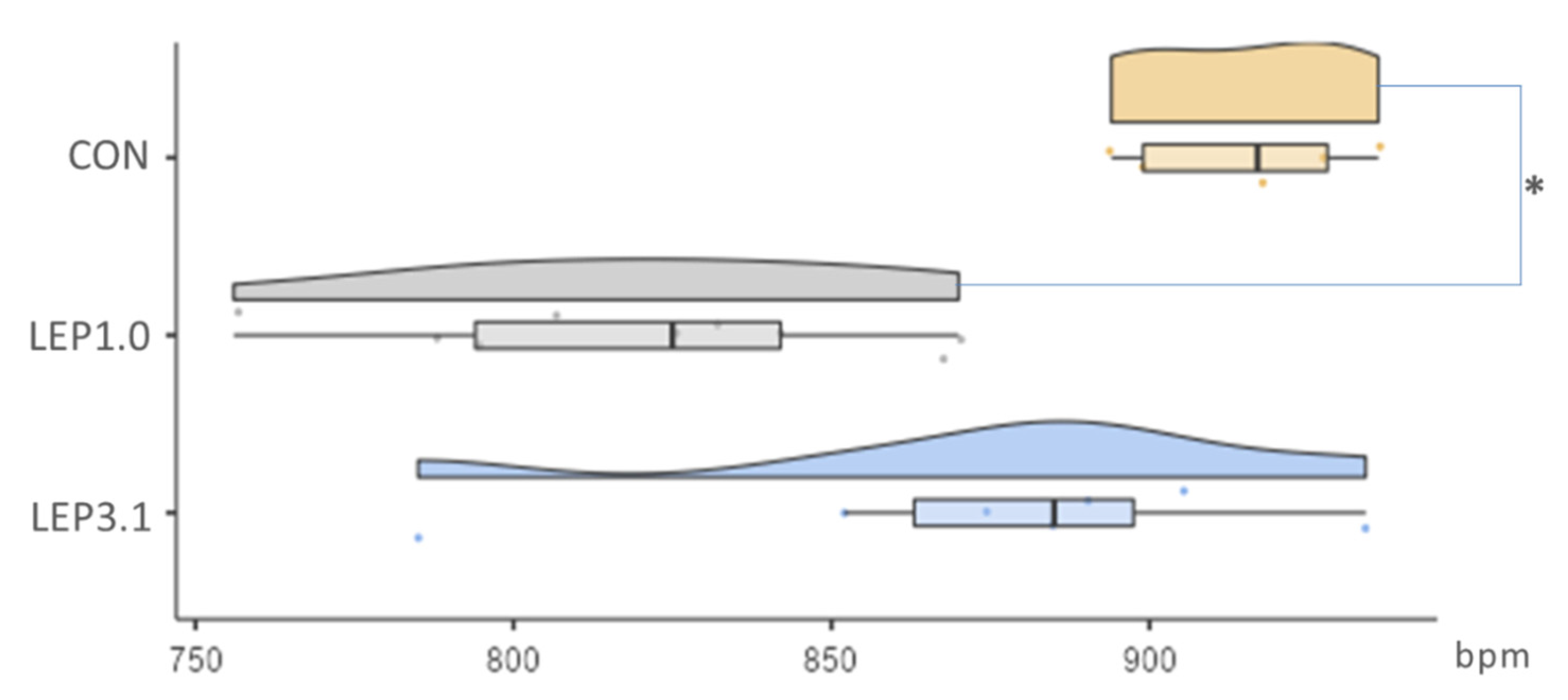
2.3. Coronary Flow Rate (CFR) and Heart Rate (HR)
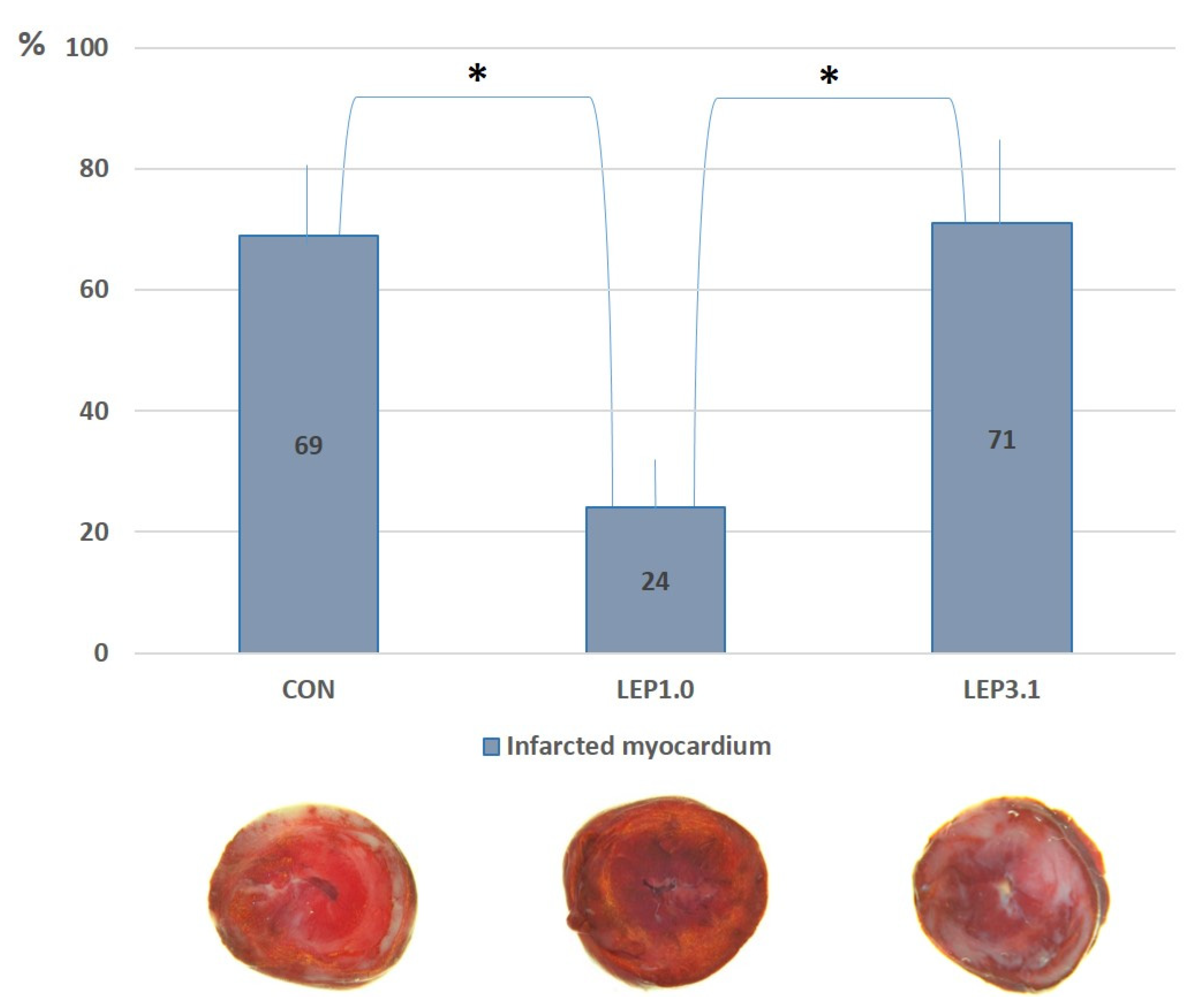
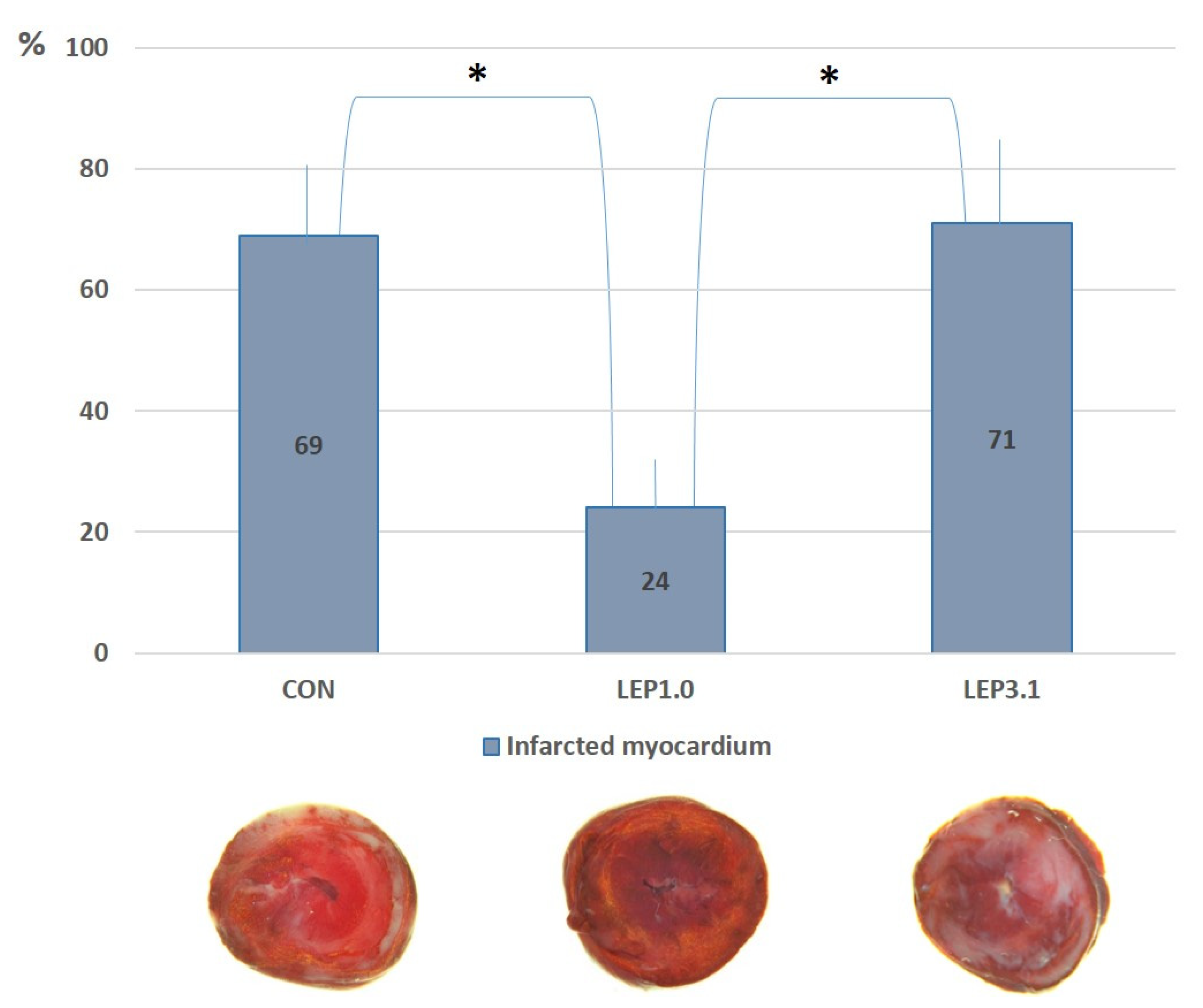
2.4. Myocardial Infarct Size
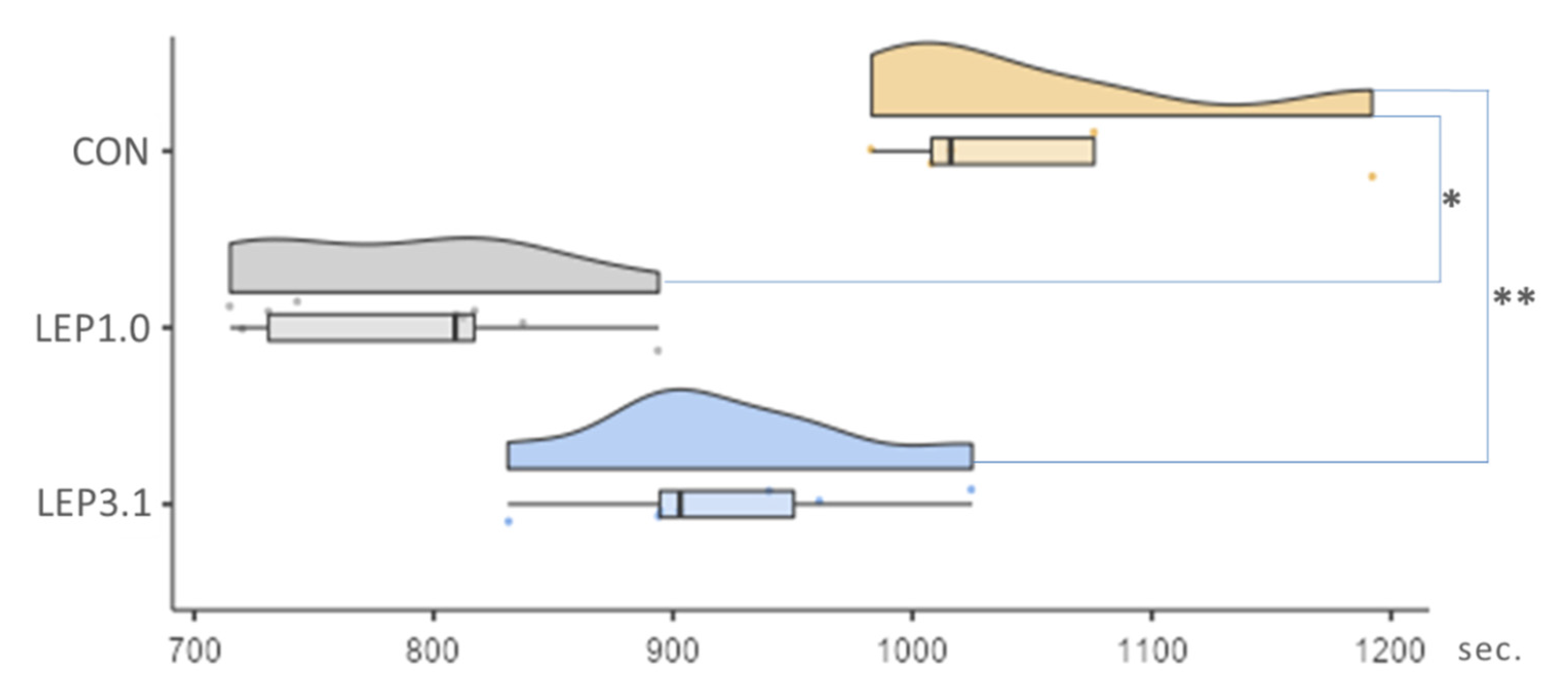
2.5. Rates of Ventricular Arrhythmia or Asystole during Reperfusion
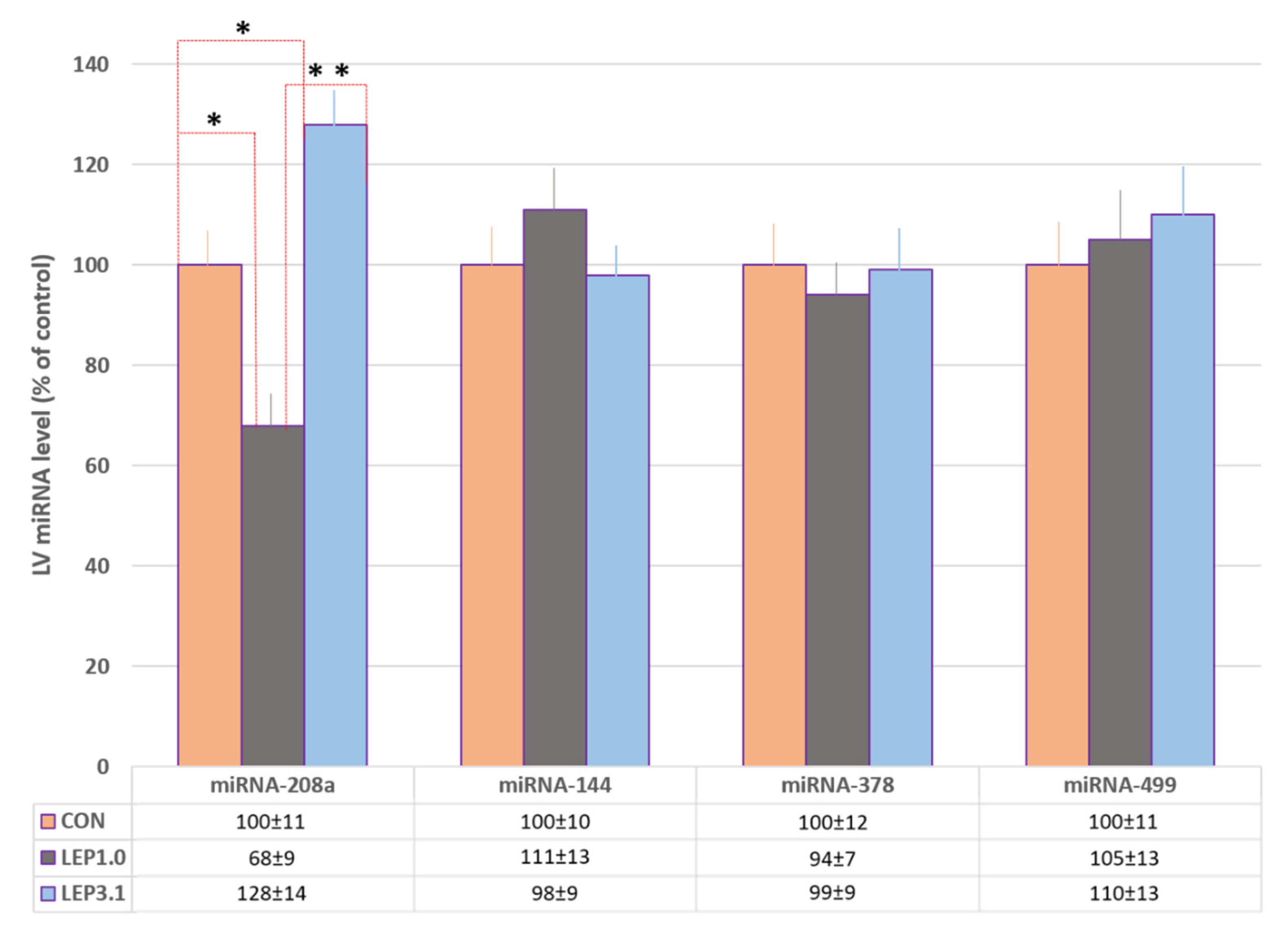
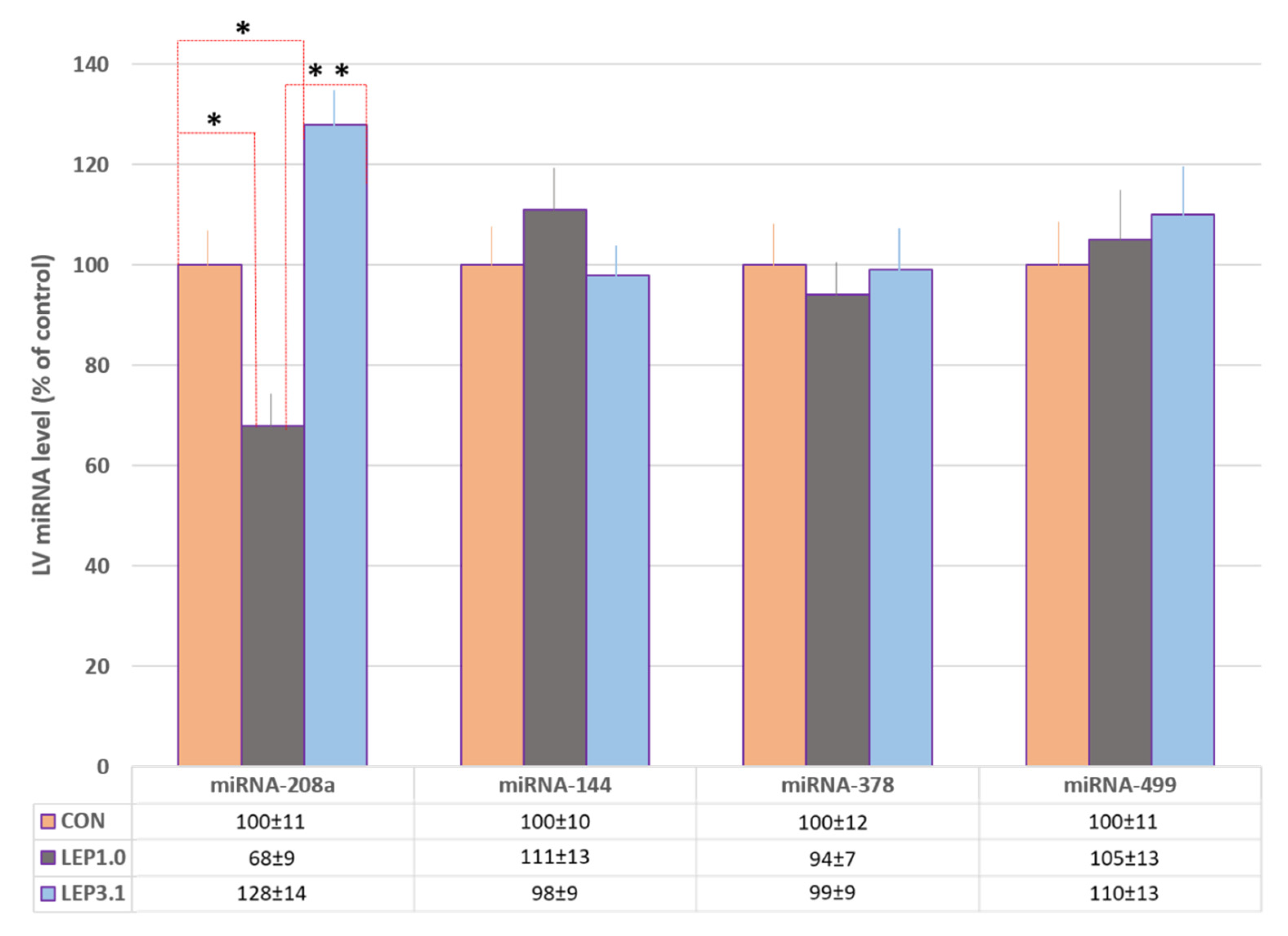
2.6. The Expression of miRNA-144, miRNA-208a, miRNA-378 and miRNA-499 in Rat Heart after Ischemic/Reperfusion Damage
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Perfusion of Isolated Hearts
4.4. Experimental Design and Protocol
4.5. Exclusion Criteria
4.6. Infarct Size Measurement
4.7. Analysis of miRNA Expression
4.8. Statistical Analysis
5. Conclusions
6. Study limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dornbush, S.; Aeddula, N.R. Physiology, Leptin. In StatPearls [Internet]; Treasure Island; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar] [PubMed]
- Benbaibeche, H.; Bounihi, A.; Koceir, E.A. Leptin level as a biomarker of uncontrolled eating in obesity and overweight. Ir. J. Med. Sci. 2021, 190, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Chen, Y.; Xin, S.; Yuan, N.; Liu, Y.; Sun, J.; Meng, X.; Qi, Y. Positive Association of Leptin and Artery Calcification of Lower Extremity in Patients with Type 2 Diabetes Mellitus: A Pilot Study. Front. Endocrinol. 2021, 12, 583575. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin-and leptin receptor-deficient rodent models: Relevance for human type 2 diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Polyakova, E.A.; Mikhaylov, E.N.; Sonin, D.L.; Cheburkin, Y.V.; Galagudza, M.M. Neurohumoral, cardiac and inflammatory markers in the evaluation of heart failure severity and progression. J. Geriatr. Cardiol. 2021, 18, E1–E20. [Google Scholar]
- Pereira, S.; Cline, D.L.; Glavas, M.M.; Covey, S.D.; Kieffer, T.J. Tissue-Specific Effects of Leptin on Glucose and Lipid Metabolism. Endocr Rev. 2021, 42, 1–28. [Google Scholar] [CrossRef]
- Cortigiani, L.; Haberka, M.; Ciampi, Q.; Bovenzi, F.; Villari, B.; Picano, E. The obesity paradox in the stress echo lab: Fat is better for hearts with ischemia or coronary microvascular dysfunction. Int. J. Obes. 2021, 45, 308–315. [Google Scholar] [CrossRef]
- Wang, J.; Yang, X. The function of miRNA in cardiac hypertrophy. Cell. Mol. Life Sci. 2012, 69, 3561–3570. [Google Scholar] [CrossRef] [Green Version]
- Grueter, C.E.; van Rooij, E.; Johnson, B.A.; DeLeon, S.M.; Sutherland, L.B.; Qi, X.; Gautron, L.; Elmquist, J.K.; Bassel-Duby, R.; Olson, E.N. A cardiac microRNA governs systemic energy homeostasis by regulation of MED13. Cell 2012, 149, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Lobo, A.M.; Donato, J., Jr. The role of leptin in health and disease. Temperature 2017, 4, 258–291. [Google Scholar] [CrossRef] [Green Version]
- Overton, J.M.; Williams, T.D.; Chambers, J.B.; Rashotte, M.E. Central leptin infusion attenuates the cardiovascular and metabolic effects of fasting in rats. Hypertension 2001, 37, 663–669. [Google Scholar] [CrossRef]
- Neumann, U.H.; Kwon, M.M.; Baker, R.K.; Kieffer, T.J. Leptin contributes to the beneficial effects of insulin treatment in streptozotocin-diabetic male mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1264–E1273. [Google Scholar] [CrossRef] [PubMed]
- Shirasaka, T.; Takasaki, M.; Kannan, H. Cardiovascular effects of leptin and orexins. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R639–R651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J. Leptin and hyperleptinemia-from friend to foe for cardiovascular function. J. Endocrinol. 2004, 181, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purdham, D.M.; Zou, M.X.; Rajapurohitam, V.; Karmazyn, M. Rat heart is a site of leptin production and action. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H2877–H2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avraham, Y.; Davidi, N.; Porat, M.; Chernoguz, D.; Magen, I.; Vorobeiv, L.; Berry, E.M.; Leker, R.R. Leptin reduces infarct size in association with enhanced expression of CB2, TRPV1, SIRT-1 and leptin receptor. Curr. Neurovasc. Res. 2010, 7, 136–143. [Google Scholar] [CrossRef]
- Smith, C.C.; Mocanu, M.M.; Davidson, S.M.; Wynne, A.M.; Simpkin, J.C.; Yellon, D.M. Leptin, the obesity-associated hormone, exhibits direct cardioprotective effects. Br. J. Pharmacol. 2006, 149, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.C.; Dixon, R.A.; Wynne, A.M.; Theodorou, L.; Ong, S.G.; Subrayan, S.; Davidson, S.M.; Hausenloy, D.J.; Yellon, D.M. Leptin-induced cardioprotection involves JAK/STAT signaling that may be linked to the mitochondrial permeability transition pore. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1265–H1270. [Google Scholar] [CrossRef] [Green Version]
- Leclercq-Meyer, V.; Considine, R.V.; Sener, A.; Malaisse, W.J. Do leptin receptors play a functional role in the endocrine pancreas? Biochem. Biophys. Res. Commun. 1996, 229, 794–798. [Google Scholar] [CrossRef]
- Ghantous, C.M.; Farhat, R.; Djouhri, L.; Alashmar, S.; Anlar, G.; Korashy, H.M.; Agouni, A.; Zeidan, A. Molecular Mechanisms of Adiponectin-Induced Attenuation of Mechanical Stretch-Mediated Vascular Remodeling. Oxid. Med. Cell. Longev. 2020, 2020, 6425782. [Google Scholar] [CrossRef]
- Minasian, S.M.; Galagudza, M.M.; Dmitriev, Y.V.; Kurapeev, D.I.; Vlasov, T.D. Myocardial protection against global ischemia with Krebs-Henseleit buffer-based cardioplegic solution. J. Cardiothorac Surg. 2013, 8, 60. [Google Scholar] [CrossRef]
- Hall, M.E.; Harmancey, R.; Stec, D.E. Lean heart: Role of leptin in cardiac hypertrophy and metabolism. World J. Cardiol. 2015, 7, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, S.; Signore, A.P.; Chen, J. Neuroprotective effects of leptin against ischemic injury induced by oxygen-glucose deprivation and transient cerebral ischemia. Stroke 2007, 38, 2329–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkasap, S.; Erkasap, N.; Koken, T.; Kahraman, A.; Uzuner, K.U.B.İ.L.A.Y.; Yazihan, N.; Ates, E. Effect of leptin on renal ischemia reperfusion damage in rats. J. Physiol. Biochem. 2004, 60, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, T.; Konturek, P.C.; Pajdo, R.; Kwiecien, S.; Ptak, A.; Sliwowski, Z.; Drozdowicz, D.; Pawlik, M.; Konturek, S.J.; Hahn, E.G. Brain-gut axis in gastroprotection by leptin and cholecystokinin against ischemia-reperfusion induced gastric lesions. J. Physiol. Pharmacol. 2001, 52, 583–602. [Google Scholar] [PubMed]
- Peliciari-Garcia, R.A.; Andrade-Silva, J.; Cipolla-Neto, J.; Carvalho, C.R.D.O. Leptin modulates norepinephrine-mediated melatonin synthesis in cultured rat pineal gland. Biomed Res. Int. 2013, 2013, 546516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gava, F.N.; da Silva, A.A.; Dai, X.; Harmancey, R.; Ashraf, S.; Omoto, A.C.; Salgado, M.C.; Moak, S.P.; Li, X.; Hall, J.E.; et al. Restoration of Cardiac Function After Myocardial Infarction by Long-Term Activation of the CNS Leptin-Melanocortin System. JACC Basic Transl Sci. 2021, 6, 55–70. [Google Scholar] [CrossRef]
- Lopaschuk, G.D. Targeting the Brain to Protect the Heart. JACC Basic Transl Sci. 2021, 6, 71–73. [Google Scholar] [CrossRef]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [Green Version]
- Abd Alkhaleq, H.; Kornowski, R.; Waldman, M.; Levy, E.; Zemel, R.; Nudelman, V.; Shainberg, A.; Miskin, R.; Hochhauser, E. Leptin modulates gene expression in the heart and cardiomyocytes towards mitigating ischemia-induced damage. Exp. Cell Res. 2020, 397, 112373. [Google Scholar] [CrossRef]
- Hall, M.E.; Maready, M.W.; Hall, J.E.; Stec, D.E. Rescue of cardiac leptin receptors in db/db mice prevents myocardial triglyceride accumulation. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E316–E325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shek, E.W.; Brands, M.W.; Hall, J.E. Chronic leptin infusion increases arterial pressure. Hypertension 1998, 31, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, Y.J. Is leptin protective against heart failure with preserved ejection fraction? A complex interrelationship among leptin, obesity, and left ventricular hypertrophy. Hypertens. Res. 2019, 42, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Tabucanon, T.; Wilcox, J.; Tang, W.H.W. Does Weight Loss Improve Clinical Outcomes in Overweight and Obese Patients with Heart Failure? Curr. Diab. Rep. 2020, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- McGaffin, K.R.; Witham, W.G.; Yester, K.A.; Romano, L.C.; O’Doherty, R.M.; McTiernan, C.F.; O’Donnell, C.P. Cardiac-specific leptin receptor deletion exacerbates ischaemic heart failure in mice. Cardiovasc. Res. 2011, 89, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanivel, R.; Eguchi, M.; Shuralyova, I.; Coe, I.; Sweeney, G. Distinct effects of short- and long-term leptin treatment on glucose and fatty acid uptake and metabolism in HL-1 cardiomyocytes. Metabolism 2006, 55, 1067–1075. [Google Scholar] [CrossRef]
- Witham, W.; Yester, K.; O’Donnell, C.P.; McGaffin, K.R. Restoration of glucose metabolism in leptin-resistant mouse hearts after acute myocardial infarction through the activation of survival kinase pathways. J. Mol. Cell. Cardiol. 2012, 53, 91–100. [Google Scholar] [CrossRef]
- Szczepaniak, L.S.; Victor, R.G.; Orci, L.; Unger, R.H. Forgotten but not gone: The rediscovery of fatty heart, the most common unrecognized disease in America. Circ. Res. 2007, 101, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Huang, J.; Hileman, S.; Martin, K.H.; Hull, R.; Davis, M.; Yu, H.G. Leptin decreases heart rate associated with increased ventricular repolarization via its receptor. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1731–H1739. [Google Scholar] [CrossRef] [Green Version]
- Satoh, N.; Ogawa, Y.; Katsuura, G.; Numata, Y.; Tsuji, T.; Hayase, M.; Ebihara, K.; Masuzaki, H.; Hosoda, K.; Yoshimasa, Y.; et al. Sympathetic activation of leptin via the ventromedial hypothalamus: Leptin-induced increase in catecholamine secretion. Diabetes 1999, 48, 1787–1793. [Google Scholar] [CrossRef]
- Lin, Y.C.; Huang, J.; Kan, H.; Castranova, V.; Frisbee, J.C.; Yu, H.G. Defective calcium inactivation causes long QT in obese insulin-resistant rat. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1013–H1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, M.; Zhang, S.; Fan, Y.; Tan, Y.; Guo, Z.; Chen, L.; Bai, L.; Jiang, D.; Hao, X.; Li, X.; et al. The Expression of microRNA in Adult Rat Heart with Isoproterenol-Induced Cardiac Hypertrophy. Cells 2020, 9, 1173. [Google Scholar] [CrossRef] [PubMed]
- Soci, U.P.R.; Cavalcante, B.R.R.; Improta-Caria, A.C.; Roever, L. The Epigenetic Role of MiRNAs in Endocrine Crosstalk Between the Cardiovascular System and Adipose Tissue: A Bidirectional View. Front. Cell Dev. Biol. 2022, 10, 910884. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Series | 60 min Conditioning Period, min | Reperfusion, min | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 15 | 30 | 45 | 60 | 5 | 30 | 60 | 90 | 120 | |
| Heart rate, beats/min | ||||||||||
| CON | 256 ± 16 | 250 ± 13 | 232 ± 21 | 260 ± 19 | 255 ± 23 | 201 ± 28 | 184 ± 18 | 216 ± 24 | 219 ± 21 | 251 ± 38 |
| LEP1.0 | 263 ± 22 | 258 ± 16 | 246 ± 17 | 257 ± 18 | 264 ± 21 | 183 ± 31 | 199 ± 22 | 234 ± 19 | 229 ± 28 | 247 ± 17 |
| LEP3.1 | 260 ± 19 | 242 ± 23 | 260 ± 14 | 268 ± 20 | 259 ± 17 | - | 192 ± 15 | 212 ± 33 | 227 ± 21 | 230 ± 25 |
| Coronary flow rate, mL/min | ||||||||||
| CON | 11.6 ± 1.3 | 10.8 ± 0.9 | 11.2 ± 2.4 | 12.1 ± 1.6 | 11.0 ± 1.7 | 3.6 ± 0.5 | 3.8 ± 1.0 | 4.2 ± 1.4 | 3.9 ± 0.7 | 4.2 ± 1.3 |
| LEP1.0 | 11.1 ± 1.0 | 11.7 ± 1.1 | 10.9 ± 1.3 | 12.2 ± 1.9 | 11.5 ± 0.8 | 4.6 ± 1.3 | 4.7 ± 0.9 | 5.3 ± 1.7 | 5.8 ± 1.1 | 5.7 ± 1.5 |
| LEP3.1 | 12.6 ± 2.1 | 12.3 ± 1.8 | 11.6 ± 2.3 | 11.9 ± 2.0 | 10.1 ± 1.1 | 2.7 ± 0.7 | 4.1 ± 1.5 | 4.2 ± 0.9 | 3.6 ± 0.8 | 3.4 ± 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polyakova, E.A.; Mikhaylov, E.N.; Minasian, S.M.; Galagudza, M.M.; Shlyakhto, E.V. Anti-Ischemic Effect of Leptin in the Isolated Rat Heart Subjected to Global Ischemia-Reperfusion: Role of Cardiac-Specific miRNAs. Cardiogenetics 2023, 13, 1-13. https://doi.org/10.3390/cardiogenetics13010001
Polyakova EA, Mikhaylov EN, Minasian SM, Galagudza MM, Shlyakhto EV. Anti-Ischemic Effect of Leptin in the Isolated Rat Heart Subjected to Global Ischemia-Reperfusion: Role of Cardiac-Specific miRNAs. Cardiogenetics. 2023; 13(1):1-13. https://doi.org/10.3390/cardiogenetics13010001
Chicago/Turabian StylePolyakova, Ekaterina A., Evgeny N. Mikhaylov, Sarkis M. Minasian, Mikhail M. Galagudza, and Evgeny V. Shlyakhto. 2023. "Anti-Ischemic Effect of Leptin in the Isolated Rat Heart Subjected to Global Ischemia-Reperfusion: Role of Cardiac-Specific miRNAs" Cardiogenetics 13, no. 1: 1-13. https://doi.org/10.3390/cardiogenetics13010001
APA StylePolyakova, E. A., Mikhaylov, E. N., Minasian, S. M., Galagudza, M. M., & Shlyakhto, E. V. (2023). Anti-Ischemic Effect of Leptin in the Isolated Rat Heart Subjected to Global Ischemia-Reperfusion: Role of Cardiac-Specific miRNAs. Cardiogenetics, 13(1), 1-13. https://doi.org/10.3390/cardiogenetics13010001