
Does DHN-Melanin Always Protect Fungi against Antifungal Drugs? The Fonsecaea/Micafungin Paradigm


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. DHN-Melanin Inhibition
2.3. Antifungal Susceptibility Test
2.4. Survival Assay
2.5. Chitin Quantification
2.6. Statistical Analyses
3. Results
3.1. High Tricyclazole Concentration Is Necessary to Partially Inhibit Fonsecaea spp. Melanization
3.2. Tricyclazole Affects Differently the MIC of Drugs with Low Antifungal Activity against Fonsecaea spp.
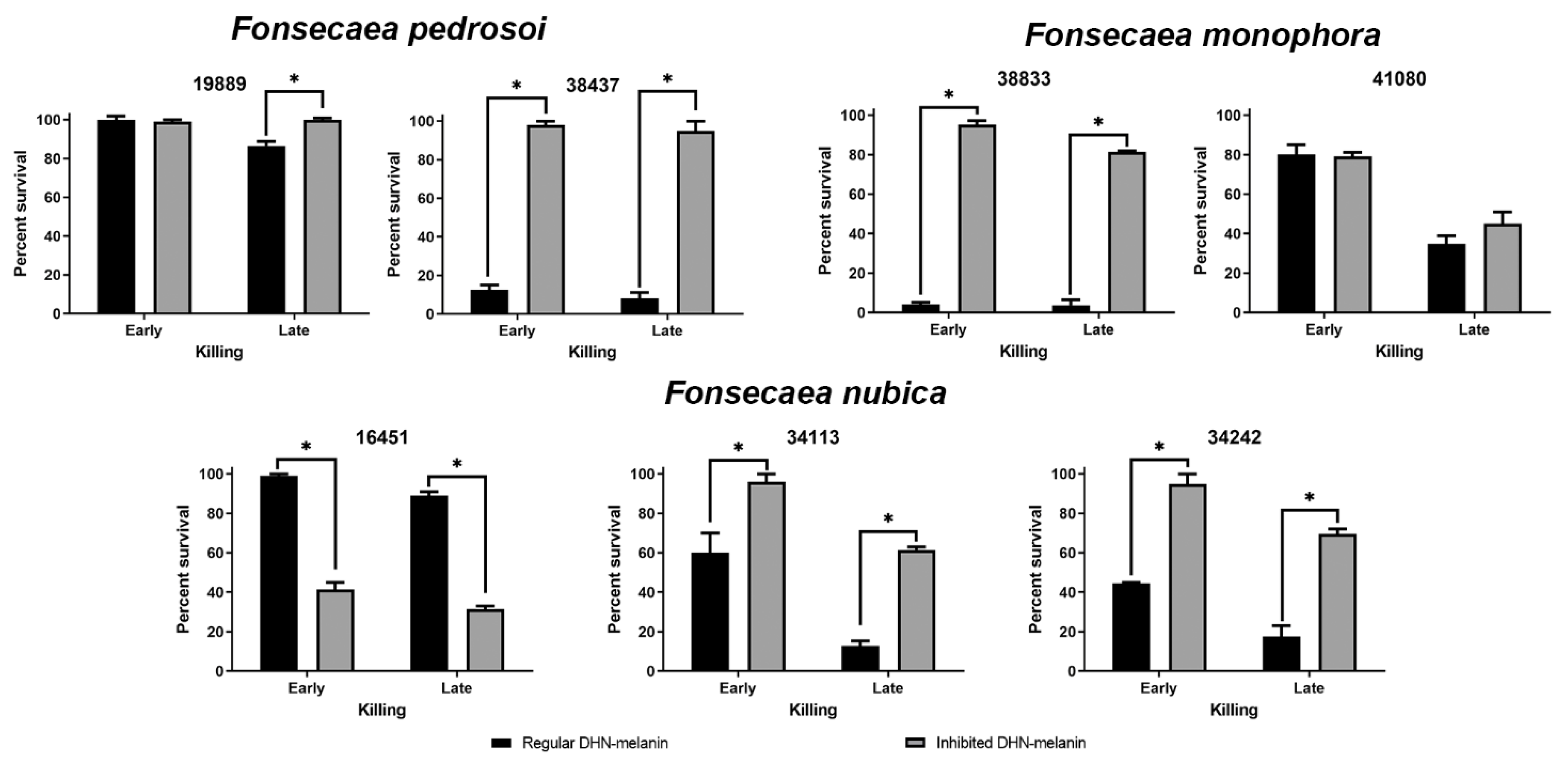
3.3. Most Fonsecaea spp. Strains with Inhibited DHN-Melanin Production Are Resistant to Micafungin Killing
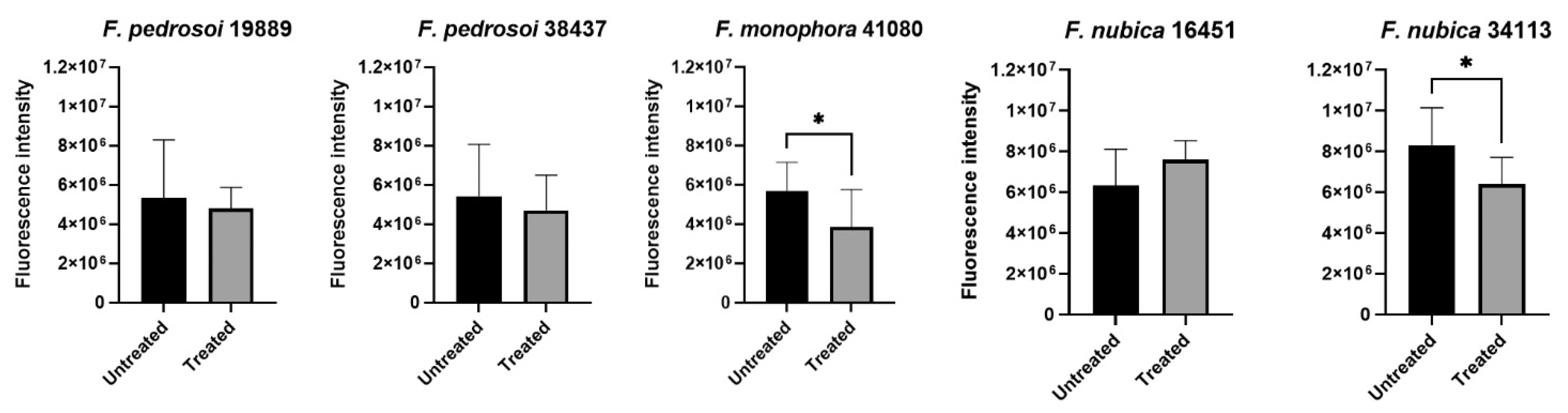
3.4. Chitin Is not Associated with Micafungin Resistance of Tricyclazole Treated Fonsecaea spp. Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Revankar, S.G. Dematiaceous fungi. Mycoses 2007, 50, 91–101. [Google Scholar] [CrossRef]
- Revankar, S.G.; Sutton, D.A. Melanized fungi in human disease. Clin. Microbiol. Rev. 2010, 23, 884–928. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O. Basic principles of the virulence of Cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo-Carvalho, M.H.G.; dos Santos, F.B.; Nosanchuk, J.D.; Zancope-Oliveira, R.M.; Almeida-Paes, R. L-dihydroxyphenylalanine induces melanin production by members of the genus Trichosporon. FEMS Yeast Res. 2014, 14, 988–991. [Google Scholar] [CrossRef] [Green Version]
- Chamilos, G.; Carvalho, A. Aspergillus fumigatus DHN-melanin. Curr. Top. Microbiol. Immunol. 2020, 425, 17–28. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; Borba-Santos, L.P.; Rozental, S.; Marco, S.; Zancopé-Oliveira, R.M.; da Cunha, M.M.L. Melanin biosynthesis in pathogenic species of Sporothrix. Fungal Biol. Rev. 2017, 31, 50–59. [Google Scholar] [CrossRef]
- Taborda, C.P.; da Silva, M.B.; Nosanchuk, J.D.; Travassos, L.R. Melanin as a virulence factor of Paracoccidioides brasiliensis and other dimorphic pathogenic fungi: A minireview. Mycopathologia 2008, 165, 331–339. [Google Scholar] [CrossRef] [Green Version]
- van Duin, D.; Casadevall, A.; Nosanchuk, J.D. Melanization of Cryptococcus neoformans and Histoplasma capsulatum reduces their susceptibilities to amphotericin B and caspofungin. Antimicrob. Agents Chemother. 2002, 46, 3394–3400. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.; Da Silva, L.B.; Gerfen, G.; SAraújo, G.R.D.; Frases, S.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Candida glabrata produces a melanin-like pigment that protects against stress conditions encountered during parasitism. Future Microbiol. 2021, 16, 509–520. [Google Scholar] [CrossRef]
- Paolo, W.F.; Dadachova, E.; Mandal, P.; Casadevall, A.; Szaniszlo, P.J.; Nosanchuk, J.D. Effects of disrupting the polyketide synthase gene WdPKS1 in Wangiella [Exophiala] dermatitidis on melanin production and resistance to killing by antifungal compounds, enzymatic degradation, and extremes in temperature. BMC Microbiol. 2006, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Alviano, C.S.; Farbiarz, S.R.; De Souza, W.; Angluster, J.; Travassos, L.R. Characterization of Fonsecaea pedrosoi melanin. J. Gen. Microbiol. 1991, 137, 837–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzen, A.J.; Cunha, M.M.L.; Miranda, K.; Hentschel, J.; Plattner, H.; da Silva, M.B.; Salgado, C.G.; de Souza, W.; Rozental, S. Ultrastructural characterization of melanosomes of the human pathogenic fungus Fonsecaea pedrosoi. J. Struct. Biol. 2008, 162, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Rozental, S.; Alviano, C.S.; de Souza, W. The in vitro susceptibility of Fonsecaea pedrosoi to activated macrophages. Mycopathologia 1994, 126, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Alviano, D.S.; Franzen, A.J.; Travassos, L.R.; Holandino, C.; Rozental, S.; Ejzemberg, R.; Alviano, C.S.; Rodrigues, M.L. Melanin from Fonsecaea pedrosoi induces production of human antifungal antibodies and enhances the antimicrobial efficacy of phagocytes. Infect. Immun. 2004, 72, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, M.M.L.; Franzen, A.J.; Alviano, D.S.; Zanardi, E.; Alviano, C.S.; De Souza, W.; Rozental, S. Inhibition of melanin synthesis pathway by tricyclazole increases susceptibility of Fonsecaea pedrosoi against mouse macrophages. Microsc. Res. Tech. 2005, 68, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.M.L.; Franzen, A.J.; Seabra, S.H.; Herbst, M.H.; Vugman, N.V.; Borba, L.P.; de Souza, W.; Rozental, S. Melanin in Fonsecaea pedrosoi: A trap for oxidative radicals. BMC Microbiol. 2010, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Zhou, X.; McCallum, N.C.; Hu, Z.; Ni, Q.Z.; Kapoor, U.; Heil, C.M.; Cay, K.S.; Zand, T.; Mantanona, A.J.; et al. Unraveling the structure and function of melanin through synthesis. J. Am. Chem. Soc. 2021, 143, 2622–2637. [Google Scholar] [CrossRef]
- Vicente, V.A.; Weiss, V.A.; Bombassaro, A.; Moreno, L.F.; Costa, F.F.; Raittz, R.T.; Leão, A.C.; Gomes, R.R.; Bocca, A.L.; Fornari, G.; et al. Comparative genomics of sibling species of Fonsecaea associated with human chromoblastomycosis. Front. Microbiol. 2017, 8, 1924. [Google Scholar] [CrossRef]
- Romero-Martinez, R.; Wheeler, M.; Guerrero-Plata, A.; Rico, G.; Torres-Guerrero, H. Biosynthesis and functions of melanin in Sporothrix schenckii. Infect. Immun. 2000, 68, 3696–3703. [Google Scholar] [CrossRef] [Green Version]
- Franzen, A.J.; Cunha, M.M.L.; Batista, E.J.O.; Seabra, S.H.; De Souza, W.; Rozental, S. Effects of tricyclazole (5-methyl-1,2,4-triazol[3,4]benzothiazole), a specific DHN-melanin inhibitor, on the morphology of Fonsecaea pedrosoi conidia and sclerotic cells. Microsc. Res. Tech. 2006, 69, 729–737. [Google Scholar] [CrossRef]
- Heidrich, D.; Pagani, D.M.; Koehler, A.; Alves, K.D.O.; Scroferneker, M.L. Effect of melanin biosynthesis inhibition on the antifungal susceptibility of chromoblastomycosis agents. Antimicrob. Agents Chemother. 2021, 65, e0054621. [Google Scholar] [CrossRef] [PubMed]
- Queiroz-Telles, F.; Purim, K.S.; Fillus, J.N.; Bordignon, G.F.; Lameira, R.P.; Van Cutsem, J.; Cauwenbergh, G. Itraconazole in the treatment of chromoblastomycosis due to Fonsecaea pedrosoi. Int. J. Derm. 1992, 31, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Silva-Rocha, W.P.; Cardoso, F.J.R.; Colalto, W.; Melo, A.S.A.; Chaves, G.M. Clinical improvement of chromoblastomycosis refractory to itraconazole successfully treated with high dose of terbinafine. J. Derm. 2013, 40, 775–776. [Google Scholar] [CrossRef] [PubMed]
- Esterre, P.; Andriantsimahavandy, A.; Ramarcel, E.R.; Pecarrere, J.L. Forty years of chromoblastomycosis in Madagascar: A review. Am. J. Trop. Med. Hyg. 1996, 55, 45–47. [Google Scholar] [CrossRef]
- Esterre, P.; Queiroz-Telles, F. Management of chromoblastomycosis: Novel perspectives. Curr. Opin. Infect. Dis. 2006, 19, 148–152. [Google Scholar] [CrossRef]
- Sawaya, B.P.; Briggs, J.P.; Schnermann, J. Amphotericin B nephrotoxicity: The adverse consequences of altered membrane properties. J. Am. Soc. Nephrol. 1995, 6, 154–164. [Google Scholar] [CrossRef]
- Coelho, R.A.; Brito-Santos, F.; Figueiredo-Carvalho, M.H.G.; Silva, J.V.D.S.; Gutierrez-Galhardo, M.C.; do Valle, A.C.F.; Zancopé-Oliveira, R.M.; Trilles, L.; Meyer, W.; Freitas, D.F.S.; et al. Molecular identification and antifungal susceptibility profiles of clinical strains of Fonsecaea spp. isolated from patients with chromoblastomycosis in Rio de Janeiro, Brazil. PLoS Negl. Trop. Dis. 2018, 12, e0006675. [Google Scholar] [CrossRef] [Green Version]
- CLSI Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi 2008; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.G.; Brito-Santos, F.; Almeida-Silva, F.; Oliveira, M.M.E.; Zancopé-Oliveira, R.M. Melanins protect Sporothrix brasiliensis and Sporothrix schenckii from the antifungal effects of terbinafine. PLoS ONE 2016, 11, e0152796. [Google Scholar] [CrossRef]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef]
- Smith, D.F.Q.; Casadevall, A. The role of melanin in fungal pathogenesis for animal hosts. Curr. Top. Microbiol. Immunol. 2019, 422, 1–30. [Google Scholar] [CrossRef]
- Gómez, B.L.; Nosanchuk, J.D. Melanin and fungi. Curr. Opin. Infect. Dis. 2003, 16, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Casadevall, A. Cryptococcal therapies and drug targets: The old, the new and the promising. Cell Microbiol. 2016, 18, 792–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, L.A.; Grisolia, J.C.; Burger, E.; de Araujo Paula, F.B.; Dias, A.L.T.; Malaquias, L.C.C. Virulence factors of Paracoccidioides brasiliensis as therapeutic targets: A review. Antonie Van Leeuwenhoek 2020, 113, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.E.; Almeida-Paes, R.; Corrêa-Moreira, D.; Borba, C.D.M.; Menezes, R.C.; Freitas, D.F.S.; Do Valle, A.C.F.; Schubach, A.D.O.; Barros, M.B.D.L.; Nosanchuk, J.D.; et al. A case of sporotrichosis caused by different Sporothrix brasiliensis strains: Mycological, molecular, and virulence analyses. Mem. Inst. Oswaldo Cruz 2019, 114, e190260. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, J.; Najafzadeh, M.J.; Badali, H.; Li, X.; Xi, L.; de Hoog, G.S. Melanization of a meristematic mutant of Fonsecaea monophora increases tolerance to stress factors while no effects on antifungal susceptibility. Mycopathologia 2011, 172, 373–380. [Google Scholar] [CrossRef]
- Polak, A.; Dixon, D.M. Loss of melanin in Wangiella dermatitidis does not result in greater susceptibility to antifungal agents. Antimicrob. Agents Chemother. 1989, 33, 1639–1640. [Google Scholar] [CrossRef] [Green Version]
- Queiroz-Telles, F.; Esterre, P.; Perez-Blanco, M.; Vitale, R.G.; Salgado, C.G.; Bonifaz, A. Chromoblastomycosis: An overview of clinical manifestations, diagnosis and treatment. Med. Mycol. 2009, 47, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.; Prados-Rosales, R.; Silva, B.M.A.; Nakouzi-Naranjo, A.; Zuzarte, M.; Chatterjee, S.; Stark, R.E.; Casadevall, A.; Gonçalves, T. Activation of melanin synthesis in Alternaria infectoria by antifungal drugs. Antimicrob. Agents Chemother. 2015, 60, 1646–1655. [Google Scholar] [CrossRef] [Green Version]
- Nakai, T.; Uno, J.; Ikeda, F.; Tawara, S.; Nishimura, K.; Miyaji, M. In vitro antifungal activity of micafungin (FK463) against dimorphic fungi: Comparison of yeast-like and mycelial forms. Antimicrob. Agents Chemother. 2003, 47, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Breda, L.C.D.; Menezes, I.G.; Paulo, L.N.M.; de Almeida, S.R. Immune sensing and potential immunotherapeutic approaches to control chromoblastomycosis. J. Fungi 2020, 7, 3. [Google Scholar] [CrossRef]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strain | Species | DHN-Melanin | Amphotericin B | Fluconazole | Flucytosine | Micafungin |
|---|---|---|---|---|---|---|
| 19889 | F. pedrosoi | regular | 4 mg/L | 16 mg/L | 16 mg/L | 8 mg/L |
| inhibited | 4 mg/L | 16 mg/L | 16 mg/L | ≥16 mg/L | ||
| 38437 | F. pedrosoi | regular | 8 mg/L | 16 mg/L | 32 mg/L | 1 mg/L |
| inhibited | 1 mg/L | 16 mg/L | 16 mg/L | 4 mg/L | ||
| 38833 | F. monophora | regular | 8 mg/L | 16 mg/L | 8 mg/L | 8 mg/L |
| inhibited | 2 mg/L | 16 mg/L | 16 mg/L | ≥16 mg/L | ||
| 41080 | F. monophora | regular | 8 mg/L | 16 mg/L | 4 mg/L | 8 mg/L |
| inhibited | 2 mg/L | 16 mg/L | 2 mg/L | 4 mg/L | ||
| 16451 | F. nubica | regular | 4 mg/L | 16 mg/L | 4 mg/L | 8 mg/L |
| inhibited | 1 mg/L | 32 mg/L | 8 mg/L | ≥16 mg/L | ||
| 34113 | F. nubica | regular | 4 mg/L | 32 mg/L | 4 mg/L | 8 mg/L |
| inhibited | 1 mg/L | 32 mg/L | 8 mg/L | ≥16 mg/L | ||
| 34242 | F. nubica | regular | 8 mg/L | 16 mg/L | 4 mg/L | 8 mg/L |
| inhibited | 2 mg/L | 32 mg/L | 8 mg/L | ≥16 mg/L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, R.A.; Figueiredo-Carvalho, M.H.G.; Silva, J.V.d.S.; Correa-Junior, D.; Frases, S.; Zancopé-Oliveira, R.M.; Freitas, D.F.S.; Almeida-Paes, R. Does DHN-Melanin Always Protect Fungi against Antifungal Drugs? The Fonsecaea/Micafungin Paradigm. Microbiol. Res. 2022, 13, 201-209. https://doi.org/10.3390/microbiolres13020017
Coelho RA, Figueiredo-Carvalho MHG, Silva JVdS, Correa-Junior D, Frases S, Zancopé-Oliveira RM, Freitas DFS, Almeida-Paes R. Does DHN-Melanin Always Protect Fungi against Antifungal Drugs? The Fonsecaea/Micafungin Paradigm. Microbiology Research. 2022; 13(2):201-209. https://doi.org/10.3390/microbiolres13020017
Chicago/Turabian StyleCoelho, Rowena Alves, Maria Helena Galdino Figueiredo-Carvalho, Juliana Vitória dos Santos Silva, Dario Correa-Junior, Susana Frases, Rosely Maria Zancopé-Oliveira, Dayvison Francis Saraiva Freitas, and Rodrigo Almeida-Paes. 2022. "Does DHN-Melanin Always Protect Fungi against Antifungal Drugs? The Fonsecaea/Micafungin Paradigm" Microbiology Research 13, no. 2: 201-209. https://doi.org/10.3390/microbiolres13020017
APA StyleCoelho, R. A., Figueiredo-Carvalho, M. H. G., Silva, J. V. d. S., Correa-Junior, D., Frases, S., Zancopé-Oliveira, R. M., Freitas, D. F. S., & Almeida-Paes, R. (2022). Does DHN-Melanin Always Protect Fungi against Antifungal Drugs? The Fonsecaea/Micafungin Paradigm. Microbiology Research, 13(2), 201-209. https://doi.org/10.3390/microbiolres13020017