
Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815





Abstract
:1. Introduction
2. Methodology
3. Mycochemistry
3.1. Primary Metabolites
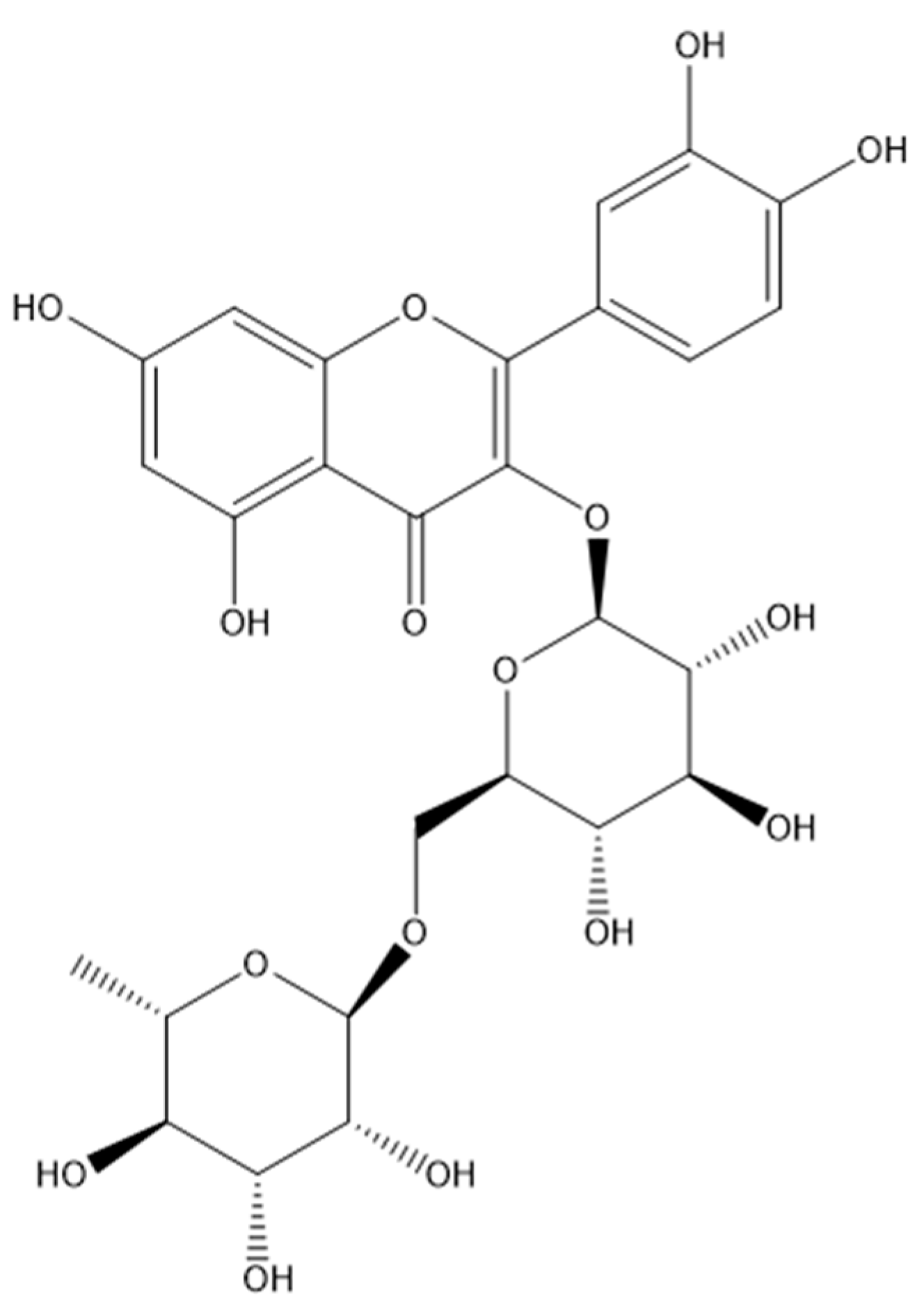
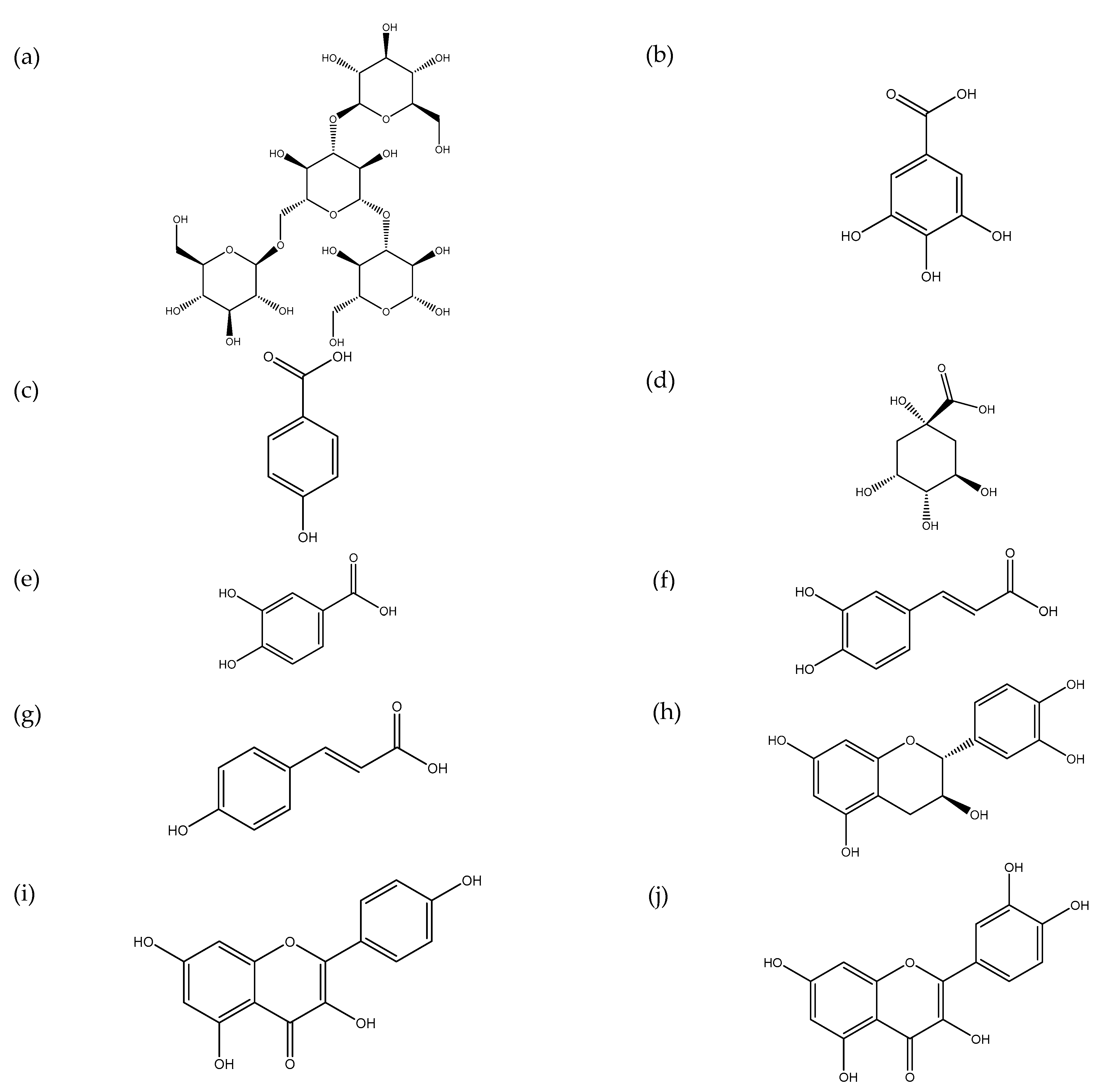
3.2. Secondary Metabolites
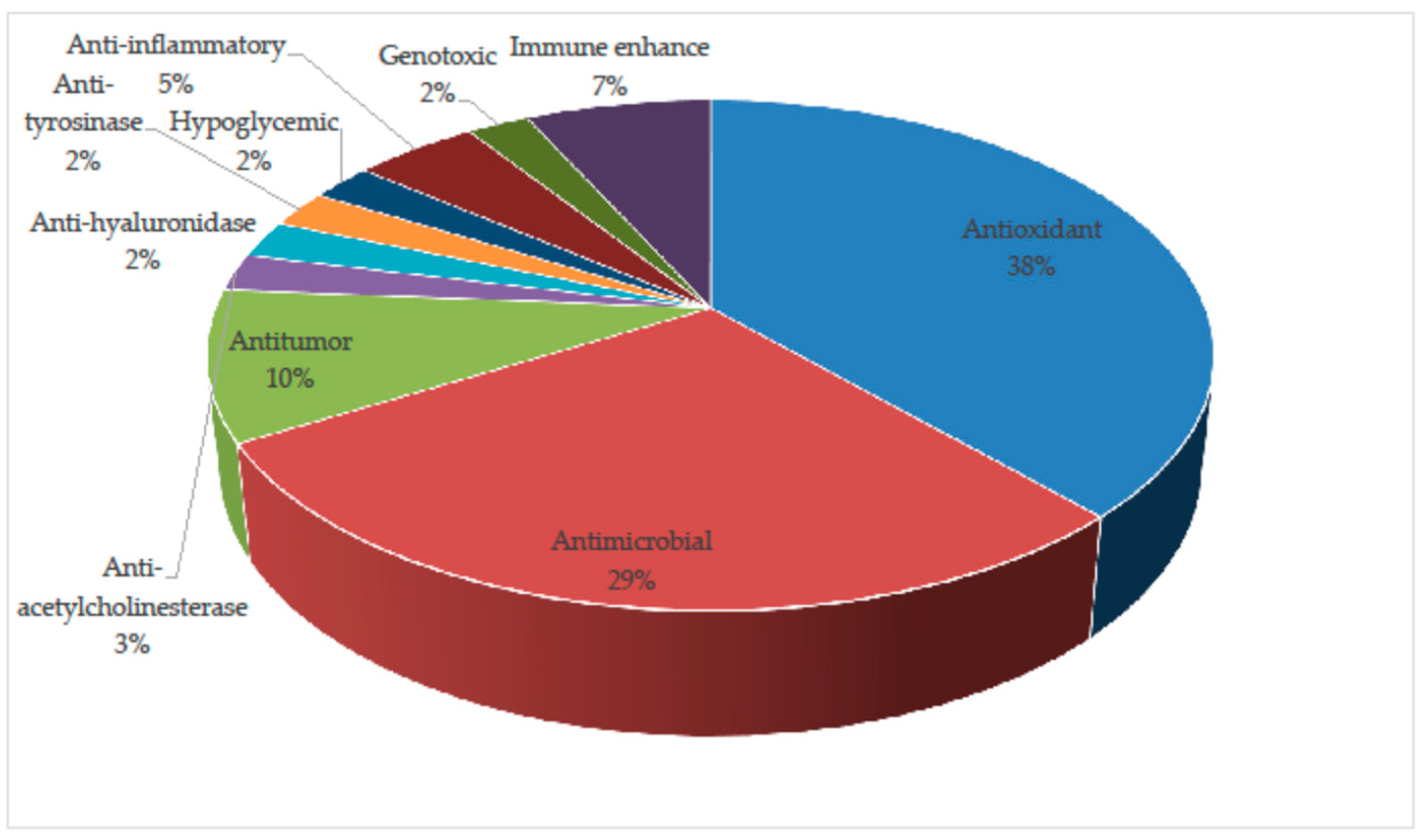
4. Biological Activities
4.1. Antioxidant Activity
4.2. Anti-Acetylcholinesterase Activity
4.3. Antimicrobial Activity
4.4. Antitumor Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AAC | total amino acid content |
| AB | antibacterial |
| ABTS | and 2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid |
| AChE | acetylcholinesterase |
| AF | antifungal |
| AM | antimicrobial |
| AO | antioxidant |
| Aq | aqueous, water |
| AV | antiviral |
| BHA | butylated hydroxyanisole |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| EPS | exopolysaccharide |
| EtOH | ethanolic |
| F | fermentation broth |
| FRAP | ferric reducing–antioxidant power |
| GC-MS | gas chromatography–mass spectrometry |
| HPLC-RI | high-performance liquid chromatography with refraction index detection |
| IR | infrared |
| IT | Italy |
| LC-MS/MS | liquid chromatography–mass spectrometry |
| M | mycelial biomass |
| MBC | minimum bactericidal concentration |
| MeOH | methanol |
| MIC | minimum inhibitory concentration |
| MW | molecular weight |
| NO | nitric oxide radical |
| OH | hydroxyl radical |
| PH | phenolic |
| PSH | polysaccharide |
| SEPS | water-soluble exopolysaccharide |
| SIPS | water-soluble intrapolysaccharide |
| SOA | superoxide anion radical |
| SPG | schizophyllan |
| SRB | Serbia |
| TCC | total carbohydrate content |
| TF | total flavonoid content |
| TP | total phenolic content |
| VOC | volatile organic compounds |
References
- López-Legarda, X.; Rostro-Alanís, M.D.; Parra-Saldívar, R.; Villa-Pulgarín, J.A.; Segura-Sánchez, F. Submerged cultivation, characterization and in vitro antitumor activity of polysaccharides from Schizophyllumradiatum. Int. J. Biol. Macromol. 2021, 186, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Robledo, G.L.; Popoff, O.F.; Amarilla, L.D.; Moncalvo, J.M.; Urcelay, C. Schizophyllumleprieurii and the Schizophyllumumbrinum lineage (Agaricales, Basidiomycota) in Argentina. Lilloa 2014, 51, 87–96. [Google Scholar]
- Available online: http://www.speciesfungorum.org/Names/Names.asp?strGenus=Schizophyllum (accessed on 30 January 2023).
- Available online: https://www.mycobank.org/ (accessed on 4 February 2023).
- Siqueira, J.P.; Sutton, D.A.; Gené, J.; García, D.; Guevara-Suarez, M.; Decock, C.A.; Wiederhold, N.P.; Guarro, J. Schizophyllumradiatum, an emerging fungus from human respiratory tract. J. Clin. Microbiol. 2016, 54, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- Sammut, C.; Alvarado, P.; Saar, I. Schizophyllumamplum (Agaricales, Schizophyllaceae): A rare Basidiomycete from Malta and Estonia. Ital. J. Mycol. 2019, 48, 50–56. [Google Scholar]
- Cooke, W.B. The genus Schizophyllum. Mycologia 1961, 53, 575–599. [Google Scholar] [CrossRef]
- Carreño-Ruiz, S.D.; Lázaro, A.A.; García, S.C.; Hernández, R.G.; Chen, J.; Navarro, G.K.; Fajardo, L.V.; Pérez, N.D.; Cruz, M.T.; Blanco, J.C.; et al. New record of Schizophyllum (Schizophyllaceae) from Mexico and the confirmation of its edibility in the humid tropics. Phytotaxa 2019, 413, 137–148. [Google Scholar] [CrossRef]
- Debnath, S.; Saha, A.K.; Panna, D. Biological activities of Schizophyllum commune Fr.: A wild edible mushroom of Tripura, North East India. J. Mycopathol. Res. 2017, 54, 469–475. [Google Scholar]
- Du, B.; Yang, Y.; Bian, Z.; Xu, B. Molecular weight and helix conformation determine intestinal anti-inflammatory effects of exopolysaccharide from Schizophyllum commune. Carbohydr. Polym. 2017, 172, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Yang, Y.D.; Bian, Z.X.; Xu, B.J. Characterization and anti-inflammatory potential of an exopolysaccharide from submerged mycelial culture of Schizophyllum commune. Front. Pharmacol. 2017, 8, 252. [Google Scholar] [CrossRef] [PubMed]
- Ellan, K.; Thayan, R.; Raman, J.; Hidari, K.I.; Ismail, N.; Sabaratnam, V. Anti-viral activity of culinary and medicinal mushroom extracts against dengue virus serotype 2: An in-vitro study. BMC Complement. Altern. Med. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Phuket, S.R.N.; Sangkaew, T.; Chanapan, P.; Techaoei, S. Biological activity of SS-glucans from edible mushroom, Schizophyllum commune in Thailand. Int. J. Appl. Pharm. 2019, 11, 110–112. [Google Scholar] [CrossRef]
- Wongaem, A.; Reamtong, O.; Srimongkol, P.; Sangtanoo, P.; Saisavoey, T.; Karnchanatat, A. Antioxidant properties of peptides obtained from the split gill mushroom (Schizophyllum commune). J. Food. Sci. Technol. 2021, 58, 680–691. [Google Scholar] [CrossRef]
- Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two Schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef]
- Karaman, M.; Čapelja, E.; Rašeta, M.; Rakić, M. Diversity, Chemistry, and Environmental Contamination of Wild Growing Medicinal Mushroom Species as Sources of Biologically Active Substances (Antioxidants, Anti-Diabetics, and AChE Inhibitors). In Biology, Cultivation and Applications of Mushrooms; Arya, A., Rusevska, K., Eds.; Springer: Singapore, 2022; Volume 8, pp. 203–257. [Google Scholar]
- Elkhateeb, W.A.; El Ghwas, D.E.; Gundoju, N.R.; Somasekhar, T.; Akram, M.; Daba, G.M. Chicken of the woods Laetiporussulphureus and Schizophyllum commune treasure of medicinal mushrooms. J. Microbiol. Biotechnol. 2021, 6, 000201. [Google Scholar]
- Acanto, R.B.; Van Helen, S.C.; Gimoto, P.H. Phytochemical screening, cytotoxic activity, and proximate analysis of split gill mushroom (Schizophyllum commune). J. Multidiscip. Res. 2022, 47, 15–29. [Google Scholar] [CrossRef]
- Nonting, S.; Satirapipathkul, C.; Charnvanich, D. The influences of extraction conditions on the content of β-glucan extracted from Schizophyllum commune processed-product residue. Int. J. Biosci. Biochem. Bioinform. 2020, 11, 8–13. [Google Scholar]
- Kumar, A.; Bharti, A.K.; Bezie, Y. Schizophyllum commune: A fungal cell-factory for production of valuable metabolites and enzymes. BioResources 2022, 17, 5420–5436. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Lee, C.; Palanisamy, S.; Prabhu, N.M.; Lee, J.; You, S. Studies on structural properties and immune-enhancing activities of glycomannans from Schizophyllum commune. Carbohydr. Polym. 2019, 218, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, C.; Fan, X.; Ma, K.; Yao, F.; Zhou, R.; Gao, H. Characterization of physicochemical and biological properties of Schizophyllum commune polysaccharide extracted with different methods. Int. J. Biol. Macromol. 2020, 156, 1425–1434. [Google Scholar] [CrossRef]
- Smirnou, D.; Knotek, P.; Nešporová, K.; Šmejkalová, D.; Pavlík, V.; Franke, L.; Velebný, V. Ultrasound-assisted production of highly-purified β-glucan schizophyllan and characterization of its immune properties. Process Biochem. 2017, 58, 313–319. [Google Scholar] [CrossRef]
- Abd Razak, D.L.; Jamaluddin, A.; Abd Rashid, N.Y.; Sani, N.A.; Abdul Manan, M. Assessment of cosmeceutical potentials of selected mushroom fruit body extracts through evaluation of antioxidant, anti-hyaluronidase and anti-tyrosinase activity. J 2020, 3, 329–342. [Google Scholar]
- Basso, V.; Schiavenin, C.; Mendonça, S.; de Siqueira, F.G.; Salvador, M.; Camassola, M. Chemical features and antioxidant profile by Schizophyllum commune produced on different agroindustrial wastes and byproducts of biodiesel production. Food Chem. 2020, 329, 127089. [Google Scholar] [CrossRef]
- Deka, A.C.; Sarma, I.D.; Dey, S.; Sarma, T.C. Antimicrobial properties and phytochemical screening of some wild macrofungi of Rani-Garbhanga reserve forest area of Assam, India. Adv. Appl. Sci. Res. 2017, 8, 17–22. [Google Scholar]
- Kaur, M.; Chadha, P.; Kaur, S.; Kaur, A.; Kaur, R.; Yadav, A.K.; Kaur, R. Schizophyllum commune induced genotoxic and cytotoxic effects in Spodoptera litura. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Woo, E.E.; Kim, J.Y.; Kim, J.S.; Kwon, S.W.; Lee, I.K.; Yun, B.S. Mannonerolidol, a new nerolidol mannoside from culture broth of Schizophyllum commune. J. Antibiot. 2019, 72, 178–180. [Google Scholar] [CrossRef]
- Hastuty, A.; Mairani, R.; Rosada, K.K. Antibacterial and anti-biofilm activities of culture filtrates from Schizophyllum commune, Coniothyrium sp., and Fusarium sp. Makara J. Sci. 2020, 24, 4. [Google Scholar]
- Khardziani, T.; Metreveli, E.; Didebulidze, K.; Elisashvili, V.I. Screening of Georgian medicinal mushrooms for their antibacterial activity and optimization of cultivation conditions for the split gill medicinal mushroom, Schizophyllum commune BCC64 (Agaricomycetes). Int. J. Med. Mushrooms 2020, 22, 659–669. [Google Scholar] [CrossRef]
- Pasailiuk, M. Total flavonoid content, lipid peroxidation and total antioxidant activity of Hericiumcoralloides, Fomes fomentarius and Schizophyllum commune cultivated by the method of direct confrontation. IJM 2020, 49, 25–37. [Google Scholar]
- Boonthatui, Y.; Chongsuwat, R.; Kittisakulnam, S. Production of antioxidant bioactive compounds during mycelium growth of Schizophyllum commune on different cereal media. CMUJ. Nat. Sci. 2021, 20, e2021032. [Google Scholar] [CrossRef]
- Boonthatui, Y.; Thakhiew, W.; Kittisakulnam, S. Quantitative changes of phenolics, flavonoids, β-glucans and antioxidant activities during the first and second harvests of split-gill mushroom (Schizophyllum commune). Agr. Nat. Resour. 2021, 5, 995–1004. [Google Scholar]
- Al Azad, S.; Ai Ping, V.C. Comparison of protein and amino acids in the extracts of two edible mushroom, Pleurotussajor-caju and Schizophyllum commune. Adv. Biosci. Biotechnol. 2021, 12, 286–296. [Google Scholar] [CrossRef]
- Mongkontanawat, N.; Thumrongchote, D. Effect of strains and extraction methods on β-glucan production, antioxidant properties, and FTIR spectra from mushroom fruiting bodies of Schizophyllum commune Fr. in Thailand. Food Res. 2021, 5, 410–415. [Google Scholar] [CrossRef]
- Wirth, S.; Krause, K.; Kunert, M.; Broska, S.; Paetz, C.; Boland, W.; Kothe, E. Function of sesquiterpenes from Schizophyllum commune in interspecific interactions. PLoS ONE 2021, 16, e0245623. [Google Scholar] [CrossRef] [PubMed]
- Toyotome, T.; Takino, M.; Takaya, M.; Yahiro, M.; Kamei, K. Identification of volatile sulfur compounds produced by Schizophyllum commune. J. Fungi 2021, 7, 465. [Google Scholar] [CrossRef] [PubMed]
- Al Azad, S.; Ping, V.C.A. Antioxidant properties and antimicrobial activity in the extracts of two edible mushroom, Pleurotussajorcaju and Schizophyllum commune. Adv. Biosci. Biotechnol. 2022, 13, 352–361. [Google Scholar] [CrossRef]
- Chen, G.G.; Zhu, Q.F.; Long, X.M.; Lu, Q.; Li, K.Y.; Chen, Q.; Xu, G.B. Antibacterial activities of the chemical constituents of Schizophyllum commune MST7-3 collected from the coal area. Nat. Prod. Res. 2022, 36, 4645–4654. [Google Scholar] [CrossRef]
- Debnath, S.; Hore, S.; Das, P.; Saha, A.K. The evaluation of selected bioactivities of fungal mycelium of a wild Schizophyllum commune Fr. from submerged cultures. J. Microbiol. Biotechnol. Food Sci. 2022, 12, e3250. [Google Scholar] [CrossRef]
- Saetang, N.; Amornlerdpison, D.; Rattanapot, T.; Ramaraj, R.; Unpaprom, Y. Processing of split gill mushroom as a biogenic material for functional food purpose. Biocatal. Agric. Biotechnol. 2022, 41, 102314. [Google Scholar] [CrossRef]
- Tosun, N.G.; Kaplan, Ö.; Türkekul, İ.; Gökçe, İ.; Özgür, A. Green synthesis of silver nanoparticles using Schizophyllum commune and Geoporasumneriana extracts and evaluation of their anticancer and antimicrobial activities. Part. Sci. Technol. 2022, 40, 801–811. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, G.; Ling, J. Medicinal fungi with antiviral effect. Molecules 2022, 27, 4457. [Google Scholar] [CrossRef] [PubMed]
- Al-Salihi, A.S.R.; Lau, B.F. Antioxidant activities and chemical composition of hot water-soluble extracts and fractions of selected Schizophyllum commune Fr. strains. Int. J. Health Sci. 2022, 6, 3976–3994. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Beara, I.N.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-inflammatory, antioxidant and enzyme inhibition activities in correlation with mycochemical profile of selected indigenous Ganoderma spp. from Balkan region (Serbia). Chem. Biodivers. 2020, 17, e2000828. [Google Scholar] [CrossRef]
- Rašeta, M.J.; Rakić, M.S.; Čapelja, E.V.; Karaman, M.A. Update on research data on the nutrient composition of mushrooms and their potentials in future human diets. In Food Chemistry, Function and Analysis, Edible Fungi: Chemical Composition, Nutrition and Health Effects; Stojković, D., Barros, L., Eds.; Royal Society of Chemistry: London, UK, 2022; Volume 2, pp. 27–67. [Google Scholar]
- Li, X.; Cheng, J.; Liu, X.; Guo, X.; Liu, Y.; Fan, W.; Lu, L.; Ma, Y.; Liu, T.; Tao, S.; et al. Origin and evolution of fusidane-type antibiotics biosynthetic pathway through multiple horizontal gene transfers. Genome Biol. Evol. 2020, 12, 1830–1840. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, J.; Meng, Y.; Aihemaiti, A.; Xu, Y.; Xiang, H.; Gao, Y.; Chen, X. Preparation, environmental application and prospect of biochar-supported metal nanoparticles: A review. J. Hazard. 2020, 388, 122026. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D. Anticholinesterase activity of selected phenolic acids and flavonoids-interaction testing in model solutions. Ann. Agric. Environ. Med. 2015, 22, 690–694. [Google Scholar] [CrossRef]
- Mandal, P. Screening, isolation and documentation of pharmaceutical importance macro fungi of kulikraiganj wildlife sanctuary, west bengal, india Parimal. Rev. Res. 2019, 7631, 1–9. [Google Scholar]
- Li, N.; Wang, C.; Georgiev, M.I.; Bajpai, V.K.; Tundis, R.; Simal-Gandara, J.; Lu, X.; Xiao, J.; Tang, X.; Qiao, X. Advances in dietary polysaccharides as anticancer agents: Structure-activity relationship. Trends Food Sci. Technol. 2021, 111, 360–377. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Species | Region | Year of Publication | Primary Metabolites | Content/Molecular Weight | Secondary Metabolites | Content | Biological Activity | Reference |
|---|---|---|---|---|---|---|---|---|
| S. commune | India | 2017 | total carbohydrate | 42.0% | - | - | Antimicrobial and antioxidant | [9] |
| crude fiber | 30.0% | |||||||
| crude protein | 15.55% | |||||||
| crude fat | 9.0% | |||||||
| lipids | 0.4% | |||||||
| S. commune | China | 2017 | low-MW EPS | 197 kDa | - | - | Anti-inflammatory | [10] |
| medium-MW EPS | 936 kDa | |||||||
| high-MW EPS | 1437 kDa | |||||||
| S. commune | China | 2017 | EPS | 2900 kDa | - | - | Anti-inflammatory | [11] |
| S. commune | India | 2017 | - | - | flavonoids saponins steroids tannins triterpenoids | qualitative mycochemical analysis | Antimicrobial | [26] |
| S. commune | Czech Republic | 2017 | ultrasonicated schizophyllan | - | - | - | Immunomodulatory | [23] |
| S. commune | India | 2018 | proteins | qualitative mycochemical analysis | glycosides | qualitative mycochemical analysis | Antioxidant, cytotoxic and genotoxic | [27] |
| flavonoids: catechin epicatechin rutin quercetin kaempferol | ||||||||
| phenolics: gallic acid chlorogenic acid caffeic acid coumaric acid | ||||||||
| triterpenoids | ||||||||
| S. commune | Korea | 2019 | - | - | nerolidol mannoside, (mannonerolidol) | - | Antibacterial and antifungal | [28] |
| schizostatin | ||||||||
| nerolidol | ||||||||
| S. commune | Malaysia | 2019 | α-glucan | 1.4 ± 0.1% | total phenolics | 0.31 ± 0.05% | Antiviral (anti- dengue) | [12] |
| β-glucan | 34.4 ± 0.4% | |||||||
| protein | 15 ± 0.4% | |||||||
| S. commune | Thailand | 2019 | β-glucan | - | total phenolics | 284.41 ± 1.22 mg GAE/g d.w. | Antioxidant | [13] |
| S. commune | Laos | 2019 | crude-PSH MWs | 710.3 × 103 g/mol 170.6 × 103 g/mol 136.3 × 103 g/mol | - | - | Immune enhancing | [21] |
| fraction MWs | 255.0 × 103 g/mol 44.8 × 103 g/mol 533.7 × 103 g/mol 93.5 × 103 g/mol 79.8 × 103 g/mol | |||||||
| carbohydrates | 50.3–65.8% | |||||||
| proteins | 1.46–20.1% | |||||||
| sulphates | 1.33–7.01% | |||||||
| glucose | 17.0–88.2% | |||||||
| mannose | 2.70–55.2% | |||||||
| galactose | 6.2–19.1% | |||||||
| xylose | 0.90–8.57% | |||||||
| S. commune | Malaysia | 2020 | polysaccharides | 66.7–76.3 mg GE/g extract | total phenolics | 14.65 mg GAE/g extract | Antioxidant, anti-hyaluronidase, and anti-tyrosinase | [24] |
| total glucan | 34.72 ± 1.94% | |||||||
| α-glucan | 4.30 ± 0.40% | |||||||
| β-glucan | 29.97 ± 1.55% | |||||||
| S. commune | Brazil | 2020 | β-glucan | 13.14% | total phenolics | 299.31 ± 3.67 mg GAE/100 g | Antioxidant | [25] |
| S. commune | China | 2020 | polysaccharides | 45.09–67.96% | - | - | Antioxidant and hypoglycemic | [22] |
| protein | 2.67–3.53% | |||||||
| uronic acid | 1.92–3.0% | |||||||
| S. commune | Indonesia | 2020 | - | - | - | - | Antibacterial | [29] |
| S. commune | Georgia | 2020 | - | - | - | - | Antibacterial | [30] |
| S. commune | Thailand | 2020 | β-glucan | 5.04–32.94% | - | - | - | [19] |
| S. commune | Ukraine | 2020 | - | - | total flavonoids | 0.64 ± 0.16 QE/g | Antioxidant | [31] |
| S. commune | India | 2020 | peptides | - | Antioxidant | [14] | ||
| MYSEKHGSGGT | 1153.23 Da | |||||||
| PGTRGAIAASSPQV | 1311.44 Da | |||||||
| MVSTLAVLGIREP | 1385.67 Da | |||||||
| EKEAAELGKGSF | 1265.37 Da | |||||||
| MSVTLLLLFISLVWVTISGLN | 2319.84 Da | |||||||
| S. commune | Thailand | 2021 | - | - | total phenolics | 0.12–8.56 mg GAE/g d.w. | Antioxidant | [32] |
| total flavonoids | 53.06–577.35 μg CE/g d.w. | |||||||
| S. commune | Thailand | 2021 | β-1,3-glucans | 64.43–114.70 mg CE/g d.w. | total phenolics | 15.15–18.92 mg GAE/g d.w.), | Antioxidant | [33] |
| β-1,3-1,6-glucans | 5.42–7.79 mg SE/g d.w | total flavonoids | 422.54–910.54 CE/g d.w | |||||
| S. commune | Malaysia | 2021 | total protein | 1.30–2.17% | - | - | - | [34] |
| total amino acid | 308.65–443.84 mg/g | |||||||
| essential amino acid | 51.44–88.56 mg/g | |||||||
| nonessential amino acid | 151.48–265.32 mg/g | |||||||
| S. commune | Italy and Serbia | 2021 | Polysaccharides | - | total phenolics | 42.74–77.52 mg GAE/g d.w. (IT) | Antioxidant antibacterial, and antiacetylcholinesterase | [15] |
| 2.09–84.60 mg GAE/g d.w. (SRB) | ||||||||
| phenolics: | ||||||||
| p-hydroxybenzoic acid | 1.87–22.19 μg/g.d.w. | |||||||
| protocatechuic acid | 1.90–5.41 μg/g.d.w. | |||||||
| gallic acid | 75.77 μg/g d.w. | |||||||
| quinic acid | 4.29–20.06 μg/g d.w. | |||||||
| S. commune | Thailand | 2021 | α-glucan | 0.41–0.57% | total phenolics | 2.54–2.64 mg GAE/g d.w. | Antioxidant | [35] |
| β-glucan | 47.94–49.20% | |||||||
| S. commune | Germany | 2021 | - | - | sesquiterpenes: | - | Antibacterial and antifungal | [36] |
| (-)-(1R,2S)-β-bisabolol | ||||||||
| β-bisabolene | ||||||||
| (E)-γ-bisabolene | ||||||||
| S. commune | Japan | 2021 | - | - | volatile sulfur compounds: | - | - | [37] |
| dimethyl disulfide | ||||||||
| dimethyl trisulfide | ||||||||
| dimethyl tetrasulfide | ||||||||
| methyl ethyl disulfide | ||||||||
| diethyl disulfide | ||||||||
| H2S | ||||||||
| S. radiatum | Colombia | 2021 | total carbohydrate | 73–79% | - | - | Antitumor and immunostimulant | [1] |
| total glucans | 11–14% | |||||||
| α-glucan | 0.4–1.7% | |||||||
| β-glucan | 11–14% | |||||||
| proteins | 1% | |||||||
| S. commune | Philippines | 2022 | proteins | saponins tannins alkaloids, flavonoids, terpenoids | - | Cytotoxic | [18] | |
| carbohydrates | ||||||||
| S. commune | Malaysia | 2022 | - | - | - | - | Antioxidant and antimicrobial | [38] |
| S. commune | China | 2022 | - | - | fusidane-type antibiotics: | - | Antibacterial | [39] |
| (E)-4-(4-hydroxy-3-methylbut-2-en-1-yl) oxybenzoic acid | ||||||||
| methyl 4-(2,3-dihydroxy-3-methylbutoxy) benzoate | ||||||||
| ethyl 4-(2,3-dihydroxy-3-methylbutoxy) benzoate | ||||||||
| (R/S)-3-hydroxy-3-((R/S)-1-hydroxyethyl) indolin-2-one | ||||||||
| helvolinic acid | ||||||||
| 3,7-diketo-cephalosporin P1 | ||||||||
| (24R)-6β-hydroxy-24-ethylcholest-4-en-3-one | ||||||||
| dankasterone A | ||||||||
| (22E,24R)-3β-hydroxyergosta-7,22-diene-6-one | ||||||||
| (3β,22E)-ergosta-5,7,22-trien-3-ol | ||||||||
| juglansnoid B | ||||||||
| trans-ferulic acid | ||||||||
| (S)-3-hydroxy-3-(2-oxopropyl) indolin-2-one | ||||||||
| schizostatin | ||||||||
| (13S)-8-oxo-(9E,11E)-8-oxo-octadeca-9,11-dien-13-olide | ||||||||
| S. commune | India | 2022 | total carbohydrate | 0.597 ± 0.06 g/100 g d.w. | total phenolics | 2.00 ± 0.05 mg GAE/g d.w. | Antioxidant and antibacterial | [40] |
| total flavonoids | 0.04 ± 0.00 mg QE/g d.w. | |||||||
| S. commune | Thailand | 2022 | schizophyllan | - | total phenolics in schizophyllan, | 6.22 ± 0.06 mg GAE/g | Antioxidant | [41] |
| total phenolics in supernatant | 12.85 ± 0.12 mg GAE/g | |||||||
| total phenolics in mushroom essence | 14.07 ± 0.09 mg GAE/g of dry extract | |||||||
| S. commune | Turkey | 2022 | - | - | - | - | Antimicrobial and anticancer | [42] |
| S. commune | Review | 2022 | hemolysin monomer ribonuclease schizophyllan | 29 kDa 20 kDa | - | - | Antiviral | [43] |
| S. commune | Malaysia and Thailand | 2022 | total carbohydrate | 229.95–596.22 mg carbohydrate/g extract | total phenolics | 6.43–37.87 mg GAE/g extract | Antioxidant | [44] |
| total proteins | 395.75–539.69 mg protein/g extract | total flavonoids | 5.77–84.55 mg RE/g extract |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mišković, J.; Rašeta, M.; Krsmanović, N.; Karaman, M. Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815. Microbiol. Res. 2023, 14, 409-429. https://doi.org/10.3390/microbiolres14010031
Mišković J, Rašeta M, Krsmanović N, Karaman M. Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815. Microbiology Research. 2023; 14(1):409-429. https://doi.org/10.3390/microbiolres14010031
Chicago/Turabian StyleMišković, Jovana, Milena Rašeta, Nenad Krsmanović, and Maja Karaman. 2023. "Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815" Microbiology Research 14, no. 1: 409-429. https://doi.org/10.3390/microbiolres14010031
APA StyleMišković, J., Rašeta, M., Krsmanović, N., & Karaman, M. (2023). Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815. Microbiology Research, 14(1), 409-429. https://doi.org/10.3390/microbiolres14010031