

Properties of Potential Plant-Growth-Promoting Bacteria and Their Effect on Wheat Growth Promotion (Triticum aestivum) and Soil Characteristics
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection
2.2. Isolation of Bacteria
2.3. Molecular Characterization
2.4. Quantitative Assessment of Potential Properties That Promote Plant Growth
2.4.1. Solubilization of Insoluble Phosphate
2.4.2. Production of Ammonia
2.4.3. Nitrogen Fixation
2.4.4. Production of Indole-3-Acetic Acid
2.4.5. Siderophore Production
2.4.6. Antifungal Activity against Fusarium oxisporum in Dual Culture Plate Method
2.5. Plant Inoculation and Experimental Design of Pot Trial
2.6. Soil Analysis
- (1)
- Soil organic carbon (SOC)—0.1–0.2 g soil, reaction with 0.4 N K2Cr2O7 in mixture with H2SO4;
- (2)
- Soil total nitrogen (STN), determined by the Kjeldahl technique—4 g soil, digestion with 20 mL 95.6% H2SO4;
- (3)
- Available phosphorus (AP)—20 g soil extracted by 0.03 N K2SO4, 5 min reaction time;
- (4)
- Exchangeable potassium (Ex-K)—5 g soil < 1.0 mm, extracted by 50 mL of CH3COONH4, pH 7, 1 h reaction time.
2.7. Statistical Analysis
3. Results
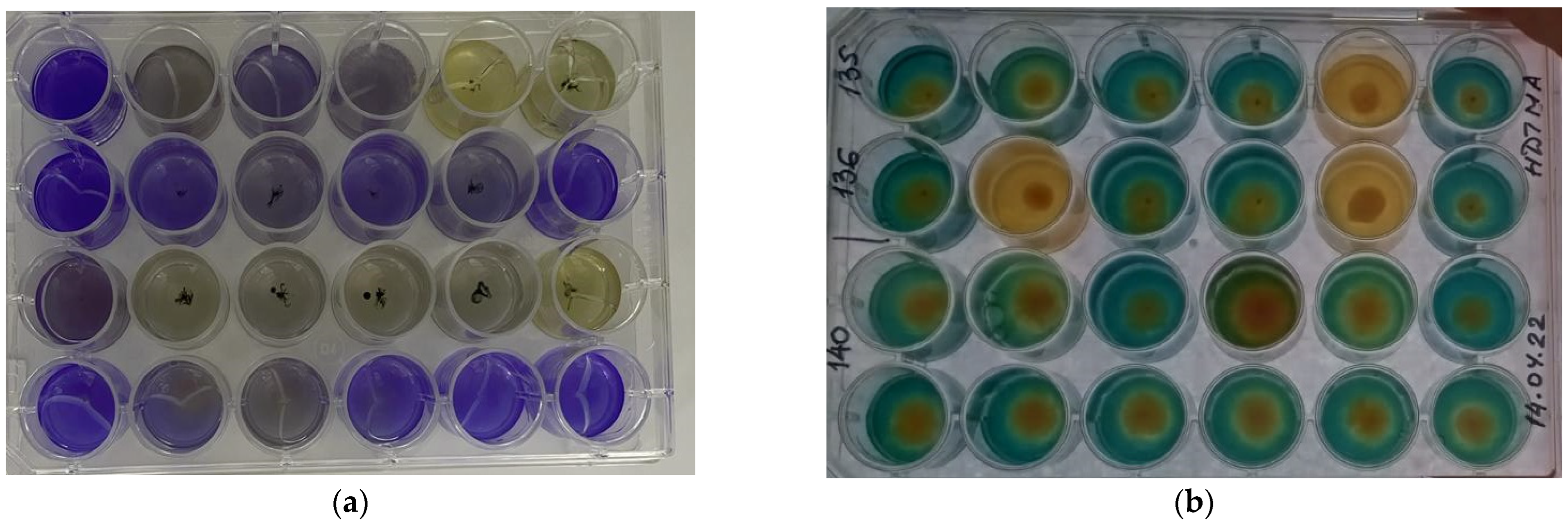
3.1. Isolation and Characterization of PGPB Isolates
3.2. Identification of Bacterial Isolates
3.3. Plant-Growth-Promoting Traits of Bacterial Strains
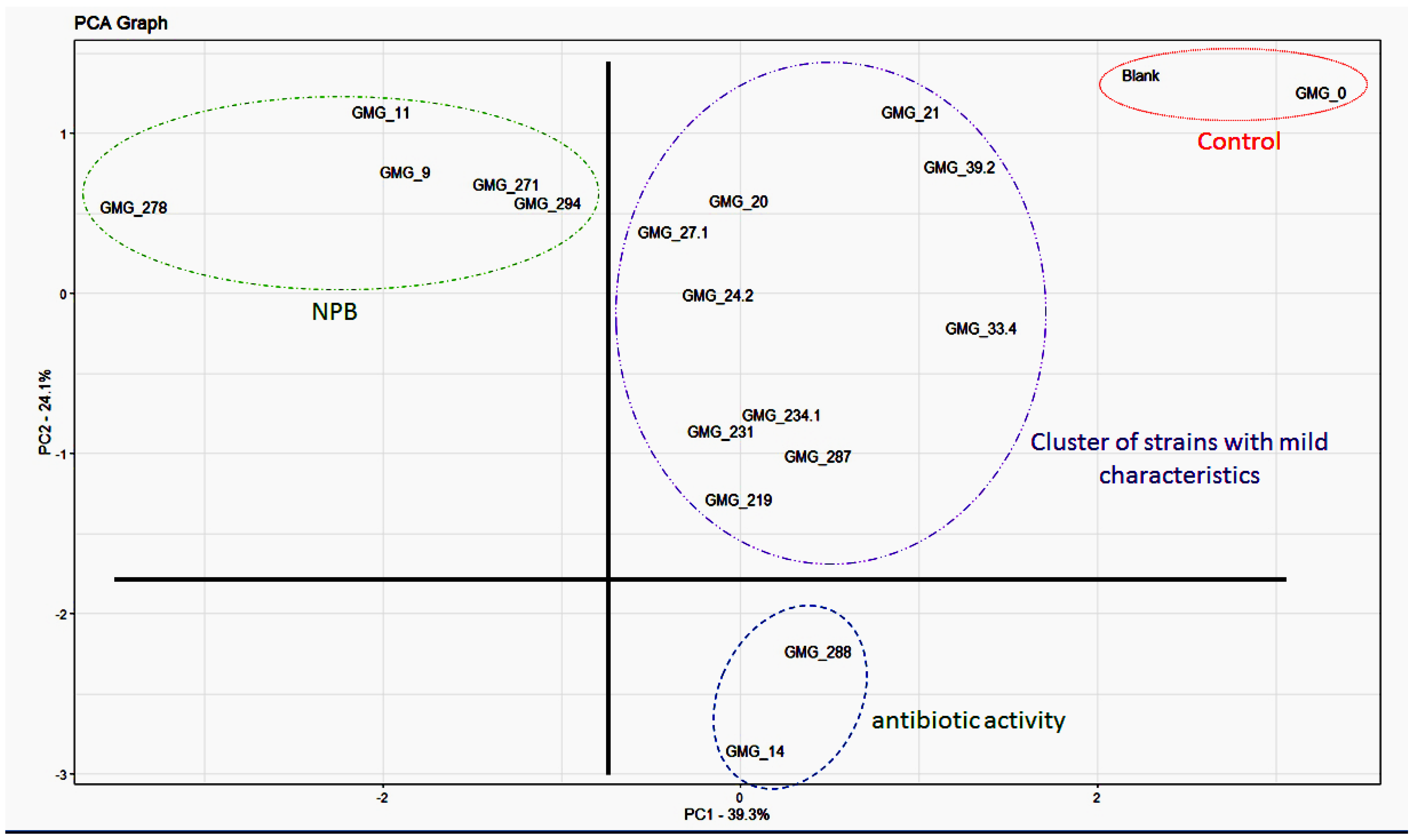
3.4. Principal Component Analysis (PCA)
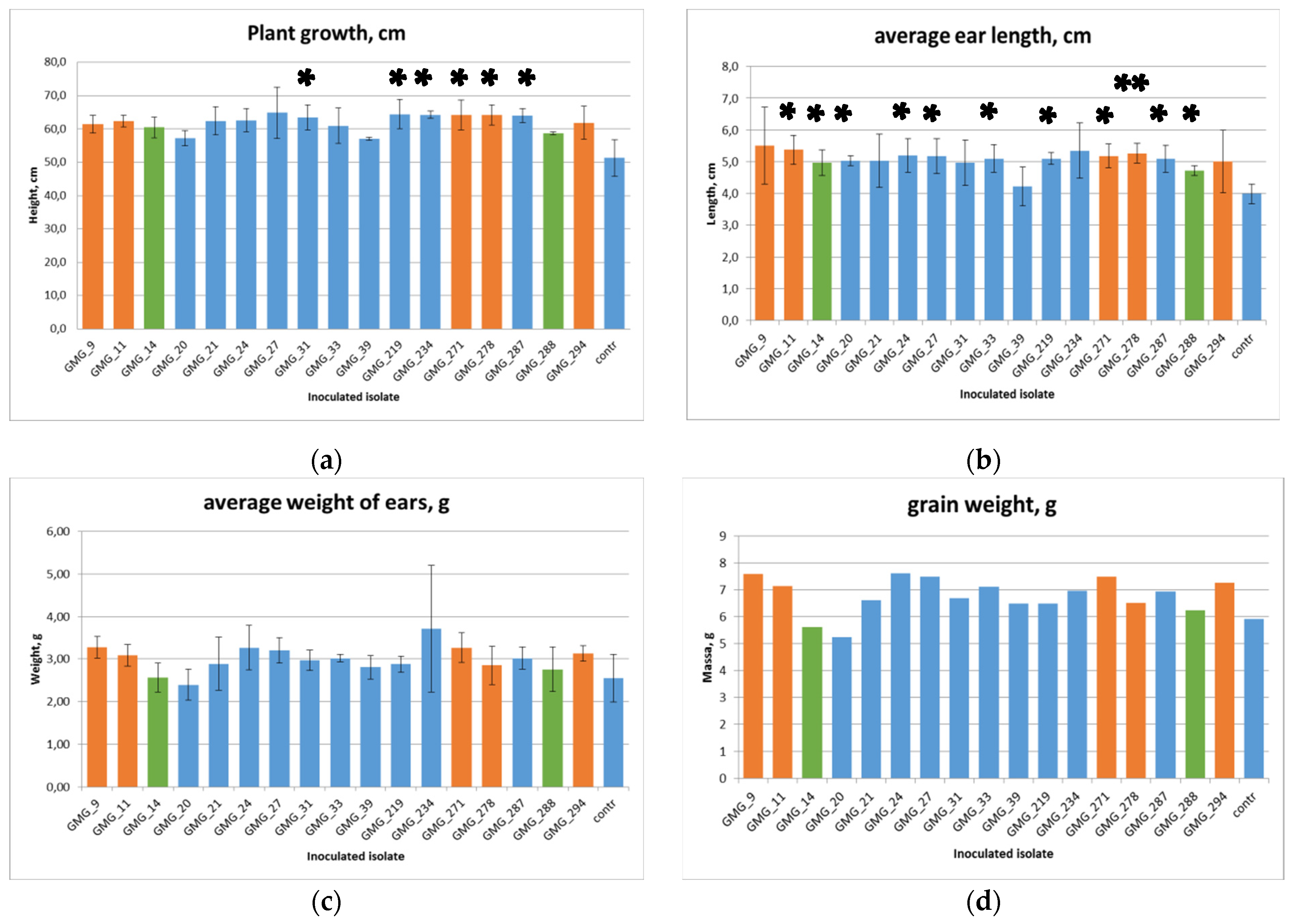
3.5. Plant Growth Parameters
3.6. Soil Characteristics
4. Discussion
5. Conclusions
- (1)
- It was not possible to identify 1–2 of the most important indicators for the primary selection of strains beneficial for plant growth. To effectively search for beneficial bacterial growth strains, it is insufficient to use only indicators such as growth on nitrogen-free media, phosphate mobilization, ammonium production, and auxin production. The development of additional effective and simple methods for screening other bacterial properties that may be beneficial to plants (e.g., ACC deaminase, enzymes, etc.) is required.
- (2)
- The selection of strains should be carried out independently based on the characteristics of “plant nutrition” and “antimicrobial activity”. Furthermore, bacteria that exhibit the maximum values of these factors can be combined into consortia.
- (3)
- Characteristics beneficial for plant growth can be revealed in almost any species of soil microorganism. In such a way, a strain of Rothia endophytica was first discovered which showed high growth ability on a nitrogen-free medium, ammonium production, and phosphate-mobilizing abilities.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Lu, K.; Liu, X.; Zhu, Y.; Liu, C. Comparative Functional Genome Analysis Reveals the Habitat Adaptation and Biocontrol Characteristics of Plant Growth-Promoting Bacteria in NCBI Databases. Microbiol. Spectr. 2023, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Lehtiniemi, M.; Outinen, O.; Puntila-Dodd, R. Citizen science provides added value in the monitoring for coastal non-indigenous species. J. Environ. Manag. 2020, 267, 110608. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.D.A.; Volpiano, C.G.; Vargas, L.K.; Granada, C.E.; Lisboa, B.B.; Passaglia, L.M.P. Use of Mineral Weathering Bacteria to Enhance Nutrient Availability in Crops: A Review. Front. Plant Sci. 2020, 11, 590774. [Google Scholar] [CrossRef] [PubMed]
- Tsukanova, K.A.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting Rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Jha, C.K.; Saraf, M. Hormonal signaling by PGPR improves plant health under stress conditions. In Bacteria in Agrobiology: Stress Management; Springer: Berlin/Heidelberg, Germany, 2011; pp. 119–140. [Google Scholar]
- Sheikh, S.; Amin, A.R.; Asra, M.; Bhagyalakshmi, N. Microbes and Their Role in Alleviation of Abiotic and Biotic Stress Tolerance in Crop Plants. In Microbial Symbionts and Plant Health: Trends and Applications for Changing Climate; Springer Nature: Singapore, 2023; pp. 109–126. [Google Scholar]
- Ruiu, L. Plant-Growth-Promoting Bacteria (PGPB) against Insects and Other Agricultural Pests. Agronomy 2020, 10, 861. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Miniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta. 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Reddy, C.N.; Arunasri, K.; Kumar, Y.D.; Krishna, K.V.; Mohan, S.V. Qualitative in vitro Evaluation of Plant Growth Promoting Activity of Electrogenic Bacteria from Biohydrogen Producing Microbial Electrolysis Cell towards Biofertilizer Application. J. Energy Environ. Sustain. 2016, 1, 47–51. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils. 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Tank, N.; Rajendran, N.; Patel, B.; Saraf, M. Evaluation and biochemical characterization of a distinctive pyoverdin from a pseudomonas isolated from chickpea rhizosphere. Braz. J. Microbiol. 2012, 43, 639. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.H.; Paul, N.C.; Deng, J.X.; Kim, Y.S.; Yun, B.S.; Yu, S.H. Biocontrol Activity of Bacillus amyloliquefaciens CNU114001 against Fungal Plant Diseases. Mycobiology 2013, 41, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.V. Agrochemical Methods of Soil Studies; Nauka: Moscow, Russia, 1975. [Google Scholar]
- Maynard, D.G.; Kalra, Y.P.; Crumbaugh, J.A. Nitrate and exchangeable ammonium nitrogen. In Soil Sampling and Methods of analysis, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 71–80. [Google Scholar]
- Wang, Q.; Li, Y.; Wang, Y. Optimizing the weight loss-on-ignition methodology to quantify organic and carbonate carbon of sediments from diverse sources. Environ. Monit. Assess. 2011, 174, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, N.A.; Campaniello, D.; Bevilacqua, A.; Cataldi, M.P.; Sinigaglia, M.; Flagella, Z.; Corbo, M.R. Isolation, Screening, and Characterization of Plant-Growth-Promoting Bacteria from Durum Wheat Rhizosphere to Improve N and P Nutrient Use Efficiency. Microorganisms 2019, 7, 541. [Google Scholar] [CrossRef] [PubMed]
- Jha, C.K.; Aeron, A.; Patel, B.V.; Maheshwari, D.K.; Saraf, M. Enterobacter: Role in Plant Growth Promotion. In Bacteria in Agrobiology: Plant Growth Metadata of the Chapter That Will Be Visualized Online; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Sarron, E.; Clément, N.; Pawlicki-Jullian, N.; Gaillard, I.; Boitel-Conti, M. Stimulating effects of Two Plant Growth-Promoting Bacteria, Enterobacter ludwigii Ez-185-17 and Raoultella terrigena Ez-555-6, on Flax Culture. In Proceedings of the 2nd International Conference on Applied Sciences (ICAS-2), Ho Chi Minh City, Vietnam, 24–25 May 2018; American Institute of Physics Inc.: College Park, MR, USA, 2018; p. 1954. [Google Scholar]
- Singh, R.P.; Mishra, S.; Jha, P.; Raghuvanshi, S.; Jha, P.N. Effect of inoculation of zinc-resistant bacterium Enterobacter ludwigii CDP-14 on growth, biochemical parameters and zinc uptake in wheat (Triticum aestivum L.) plant. Ecol. Eng. 2018, 116, 163–173. [Google Scholar] [CrossRef]
- Shoebitz, M.; Ribaudo, C.M.; Pardo, M.A.; Cantore, M.L.; Ciampi, L.; Curá, J.A. Plant growth promoting properties of a strain of Enterobacter ludwigii isolated from Lolium perenne rhizosphere. Soil Biol. Biochem. 2009, 41, 1768–1774. [Google Scholar] [CrossRef]
- Yagmur, B.; Gunes, A. Evaluation of the Effects of Plant Growth Promoting Rhizobacteria (PGPR) on Yield and Quality Parameters of Tomato Plants in Organic Agriculture by Principal Component Analysis (PCA). Gesunde Pflanz. 2021, 73, 219–228. [Google Scholar] [CrossRef]
- Tahir, M.; Mirza, M.S.; Zaheer, A.; Dimitrov, M.R.; Smidt, H.; Hameed, S. Isolation and identification of phosphate solubilizer Azospirillum, Bacillus and Enterobacter strains by 16SrRNA sequence analysis and their effect on growth of wheat (Triticum aestivum L.). Aust. J. Crop Sci. 2013, 7, 1284–1292. [Google Scholar]
- Kuzina, E.; Mukhamatdyarova, S.; Sharipova, Y.; Makhmutov, A.; Belan, L.; Korshunova, T. Influence of Bacteria of the Genus Pseudomonas on Leguminous Plants and Their Joint Application for Bioremediation of Oil Contaminated Soils. Plants 2022, 11, 3396. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, R.K.; Li, H.B.; Guo, D.J.; Sharma, A.; Verma, K.K.; Solanki, M.K.; Upadhyay, S.K.; Lakshmanan, P.; Yang, L.T.; et al. Nitrogen fixation and phytohormone stimulation of sugarcane plant through plant growth promoting diazotrophic Pseudomonas. Biotechnol. Genet. Eng. Rev. 2023, 22, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Ma, Y.N.; Wang, J.; Xia, Z.; Wei, H.L. Genomic insights into a plant growth-promoting Pseudomonas koreensis strain with cyclic lipopeptide-mediated antifungal activity. Microbiologyopen 2020, 9, 1–15. [Google Scholar] [CrossRef]
- Sawada, H.; Fujikawa, T.; Nishiwaki, Y.; Horita, H. Pseudomonas kitaguniensis sp. Nov., a pathogen causing bacterial rot of welsh onion in Japan. Int. J. Syst. Evol. Microbiol. 2020, 70, 3018–3026. [Google Scholar] [CrossRef]
- Lee, S.D.; Jeon, D.; Kim, I.S.; Choe, H.; Kim, J.S. Rahnella aceris sp. nov., isolated from sap drawn from Acer pictum. Arch. Microbiol. 2020, 202, 2411–2417. [Google Scholar] [CrossRef]
- Xu, S.; Zhao, Y.; Peng, Y.; Shi, Y.; Xie, X.; Chai, A.; Li, B.; Li, L. Comparative Genomics Assisted Functional Characterization of Rahnella aceris ZF458 as a Novel Plant Growth Promoting Rhizobacterium. Front. Microbiol. 2022, 13, 850084. [Google Scholar] [CrossRef]
- Landa-Acuña, D.; Toro, M.; Santos-Mendoza, R.; Zúñiga-Dávila, D. Role of Rahnella aquatilis AZO16M2 in Phosphate Solubilization and Ex Vitro Acclimatization of Musa acuminata var. Valery. Microorg. 2023, 11, 1596. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.J.; Zhang, J.-L.; Zhang, D.-F.; Zhou, Z.-L.; Liu, M.-J.; Zhu, W.-Y.; Zhao, L.-X.; Xu, L.-H.; Li, W.-J. Rothia endophytica sp. nov., an actinobacterium isolated from Dysophylla stellata (Lour.) Benth. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 11, 3964–3969. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No | Strain | Nitrogen Fixation, % | Phosphate Solubilization, µg/ml | Ammonia Production, µmol/mL | Siderophore Production, % | IAA Production, µg/mL | Antifungal Activity, mm |
|---|---|---|---|---|---|---|---|
| GMG_9 | Rothia endophytica | 41.8 ± 4.6 | 248.3 ± 35.5 | 8.5 ± 0.3 | 7.1 ± 4.0 | 0.0 ± 0.3 | 0.0 |
| GMG_11 | Pseudomonas koreensis | 41.5 ± 10.1 | 294.8 ± 28.1 | 8.0 ± 0.7 | 0.0 ± 0.7 | 0.0 ± 0.1 | 0.0 |
| GMG_14 | Pseudomonas silesiensis | 3.9 ± 1.6 | 231.4 ± 39.9 | 4.8 ± 1.3 | 76.8 ± 5.2 | 0.0 ± 0.4 | 16.2 ± 5.1 |
| GMG_20 | Pantoea agglomerans | 3.1 ± 1.2 | 105.1 ± 18.8 | 6.5 ± 1.6 | 0.0 ± 0.3 | 6.7 ± 0.3 | 0.0 |
| GMG_21 | Rhodococcus erythropolis | 5.5 ± 1.3 | 54.3 ± 7.0 | 4.7 ± 1.4 | 0.0 ± 0.3 | 0.0 ± 0.1 | 0.0 |
| GMG_24 | Enterobacter cloacae | 0.6 ± 0.4 | 94.2 ± 18.1 | 8.5 ± 0.2 | 8.4 ± 4.6 | 5.2 ± 0.6 | 0.0 |
| GMG_27 | Variovorax paradoxus | 1.9 ± 1.2 | 194.1 ± 24.8 | 6.7 ± 0.8 | 0.0 ± 0.2 | 5.6 ± 0.4 | 0.0 |
| GMG_31 | Hylemonella gracilis | 0.4 ± 0.4 | 0.0 ± 10.9 | 8.5 ± 0.5 | 42.8 ± 12.5 | 4.9 ± 0.1 | 0.0 |
| GMG_33.4 | Agrobacterium arsenijevicii | 4.1 ± 1.5 | 0.0 ± 8.6 | 5.9 ± 0.6 | 0.0 ± 0.2 | 0.0 ± 0.3 | 11.7 ± 3.2 |
| GMG_39.2 | Azotobacter chroococcum | 0.9 ± 0.2 | 128.8 ± 20.4 | 6.7 ± 0.8 | 0.0 ± 0.3 | 0.0 ± 0.3 | 0.0 |
| GMG_219 | Pseudomonas kitaguniensis | 1.8 ± 0.9 | 75.5 ± 12.9 | 8.0 ± 0.2 | 61.7 ± 5.3 | 0.0 ± 0.2 | 0.0 |
| GMG_234 | Acinetobacter oryzae | 9.2 ± 4.1 | 65.8 ± 25.2 | 7.8 ± 0.6 | 3.5 ± 0.7 | 0.2 ± 0.4 | 12.3 ± 2.7 |
| GMG_271 | Pseudomonas kitaguniensis | 26.9 ± 3.2 | 151.2 ± 36.3 | 6.4 ± 0.5 | 7.8 ± 3.7 | 12.3 ± 4.6 | 0.0 |
| GMG_278 | Enterobacter ludwigii | 31.5 ± 8.6 | 287.3 ± 31.9 | 8.1 ± 0.3 | 7.8 ± 1.2 | 37.8 ± 0.9 | 0.0 |
| GMG_287 | Rahnella aquatilis | 9.3 ± 4.0 | 18.3 ± 10.8 | 8.5 ± 0.9 | 0.0 ± 0.4 | 1.6 ± 0.5 | 18.2 ± 3.5 |
| GMG_288 | Enterobacter amnigenus | 2.5 ± 1.6 | 52.3 ± 14.3 | 7.5 ± 1.2 | 39.8 ± 2.3 | 3.0 ± 0.3 | 17.5 ± 2.6 |
| GMG_294 | Rahnella aceris | 21.2 ± 4.9 | 214.0 ± 33.2 | 7.4 ± 0.5 | 3.1 ± 0.3 | 1.0 ± 0.7 | 0.0 |
| GMG_0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| PCA1 | PCA2 | |
|---|---|---|
| NFM | −0.50 | 0.27 |
| PM | −0.58 | −0.06 |
| Amm | −0.45 | −0.29 |
| IAA | −0.42 | −0.003 |
| Syd | −0.07 | −0.66 |
| AntiF | 0.12 | −0.63 |
| Ammonia Nitrogen (N-NH4), mg per kg of soil | Nitrate Nitrogen (N-NO3), mg per kg of soil | Soil Total Nitrogen (STN), % | Soil Humus (SH), % | Soil Organic Carbon (SOC), % | Available Phosphorus (AP), mg per kg of soil | Exchangeable Potassium (Ex-K), mg per kg of soil | |
|---|---|---|---|---|---|---|---|
| GMG_9 | 1.81 ± 0.27 | 6.33 ± 1.9 | 0.19 ± 0.02 | 3.58 ± 0.54 | 3.94 ± 0.24 | 0.24 ± 0.01 | 158 ± 15.8 |
| GMG_11 | 1.79 ± 0.27 | 4.29 ± 1.29 | 0.21 ± 0.02 | 3.55 ± 0.53 | 4.11 ± 0.25 | 0.18 ± 0.01 | 158 ± 15.8 |
| GMG_14 | 1.29 ± 0.19 | 3.89 ± 1.17 | 0.2 ± 0.02 | 3.37 ± 0.51 | 4 ± 0.25 | 0.1 ± 0.02 | 158 ± 15.8 |
| GMG_20 | 1.02 ± 0.15 | 3.7 ± 1.11 | 0.2 ± 0.02 | 4.32 ± 0.65 | 3.87 ± 0.25 | 0.18 ± 0.01 | 172 ± 17.2 |
| GMG_21 | 1.4 ± 0.21 | 3.53 ± 1.06 | 0.2 ± 0.02 | 4.34 ± 0.65 | 3.97 ± 0.28 | 0.21 ± 0.02 | 186 ± 18.6 |
| GMG_24 | 0.89 ± 0.13 | 3.36 ± 1.01 | 0.2 ± 0.02 | 4.25 ± 0.64 | 4.01 ± 0.25 | 0.42 ± 0.02 | 158 ± 15.8 |
| GMG_27 | 3.62 ± 0.54 | 3.36 ± 1.01 | 0.18 ± 0.02 | 3.96 ± 0.59 | 3.93 ± 0.27 | 0.11 ± 0.01 | 144 ± 14.4 |
| GMG_31 | 1.55 ± 0.23 | 3.89 ± 1.17 | 0.2 ± 0.02 | 4.08 ± 0.61 | 4.07 ± 0.06 | 0.09 ± 0.02 | 158 ± 15.8 |
| GMG_33.4 | 2.42 ± 0.36 | 3.36 ± 1.01 | 0.2 ± 0.02 | 3.64 ± 0.55 | 4.15 ± 0.26 | 0.13 ± 0.01 | 158 ± 15.8 |
| GMG_39.2 | 1.59 ± 0.24 | 3.7 ± 1.11 | 0.2 ± 0.02 | 3.61 ± 0.54 | 3.97 ± 0.27 | 0.13 ± 0.01 | 144 ± 14.4 |
| GMG_219 | 0.94 ± 0.14 | 3.36 ± 1.01 | 0.2 ± 0.02 | 4.15 ± 0.62 | 4.12 ± 0.25 | 0.12 ± 0.02 | 172 ± 17.2 |
| GMG_234 | 0.83 ± 0.12 | 3.53 ± 1.06 | 0.22 ± 0.02 | 3.83 ± 0.57 | 4.27 ± 0.25 | 0.18 ± 0.02 | 186 ± 18.6 |
| GMG_271 | 0.72 ± 0.11 | 3.2 ± 0.96 | 0.2 ± 0.02 | 3.05 ± 0.46 | 4.01 ± 0.24 | 0.17 ± 0.01 | 158 ± 15.8 |
| GMG_278 | 1.16 ± 0.17 | 4.08 ± 1.22 | 0.2 ± 0.02 | 3.27 ± 0.49 | 3.93 ± 0.26 | 0.14 ± 0 | 172 ± 17.2 |
| GMG_287 | 2.88 ± 0.43 | 3.36 ± 1.01 | 0.21 ± 0.02 | 4.18 ± 0.63 | 4.18 ± 0.26 | 0.1 ± 0.01 | 158 ± 15.8 |
| GMG_288 | 1.31 ± 0.2 | 3.89 ± 1.17 | 0.19 ± 0.02 | 3.5 ± 0.53 | 2.21 ± 0.25 | 0.17 ± 0.01 | 158 ± 15.8 |
| GMG_294 | 1.92 ± 0.29 | 3.53 ± 1.06 | 0.19 ± 0.02 | 4.6 ± 0.69 | 4.02 ± 0.25 | 0.26 ± 0.02 | 158 ± 15.8 |
| GMG_0 | 4.82 ± 0.72 | 3.37 ± 1.01 | 0.22 ± 0.02 | 3.8 ± 0.57 | 4.16 ± 0.24 | 0.2 ± 0 | 200 ± 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronina, E.; Sokolova, E.; Tromenschleger, I.; Mishukova, O.; Hlistun, I.; Miroshnik, M.; Savenkov, O.; Buyanova, M.; Ivanov, I.; Galyamova, M.; et al. Properties of Potential Plant-Growth-Promoting Bacteria and Their Effect on Wheat Growth Promotion (Triticum aestivum) and Soil Characteristics. Microbiol. Res. 2024, 15, 20-32. https://doi.org/10.3390/microbiolres15010002
Voronina E, Sokolova E, Tromenschleger I, Mishukova O, Hlistun I, Miroshnik M, Savenkov O, Buyanova M, Ivanov I, Galyamova M, et al. Properties of Potential Plant-Growth-Promoting Bacteria and Their Effect on Wheat Growth Promotion (Triticum aestivum) and Soil Characteristics. Microbiology Research. 2024; 15(1):20-32. https://doi.org/10.3390/microbiolres15010002
Chicago/Turabian StyleVoronina, Elena, Ekaterina Sokolova, Irina Tromenschleger, Olga Mishukova, Inna Hlistun, Matvei Miroshnik, Oleg Savenkov, Maria Buyanova, Ilya Ivanov, Maria Galyamova, and et al. 2024. "Properties of Potential Plant-Growth-Promoting Bacteria and Their Effect on Wheat Growth Promotion (Triticum aestivum) and Soil Characteristics" Microbiology Research 15, no. 1: 20-32. https://doi.org/10.3390/microbiolres15010002
APA StyleVoronina, E., Sokolova, E., Tromenschleger, I., Mishukova, O., Hlistun, I., Miroshnik, M., Savenkov, O., Buyanova, M., Ivanov, I., Galyamova, M., & Smirnova, N. (2024). Properties of Potential Plant-Growth-Promoting Bacteria and Their Effect on Wheat Growth Promotion (Triticum aestivum) and Soil Characteristics. Microbiology Research, 15(1), 20-32. https://doi.org/10.3390/microbiolres15010002