

Effect of Plant Growth Promoting Rhizobacteria on the Development and Biochemical Composition of Cucumber under Different Substrate Moisture Levels

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Site and Conditions
2.2. Morphometric Variables and Phytochemical Compounds
2.3. Total Phenol Contents, Flavonoids, and Antioxidant Capacity
2.4. Nitrogen Content in Plant
2.5. Phosphorus Content in Plant
2.6. Statistical Analyses
3. Results
3.1. Plant Height
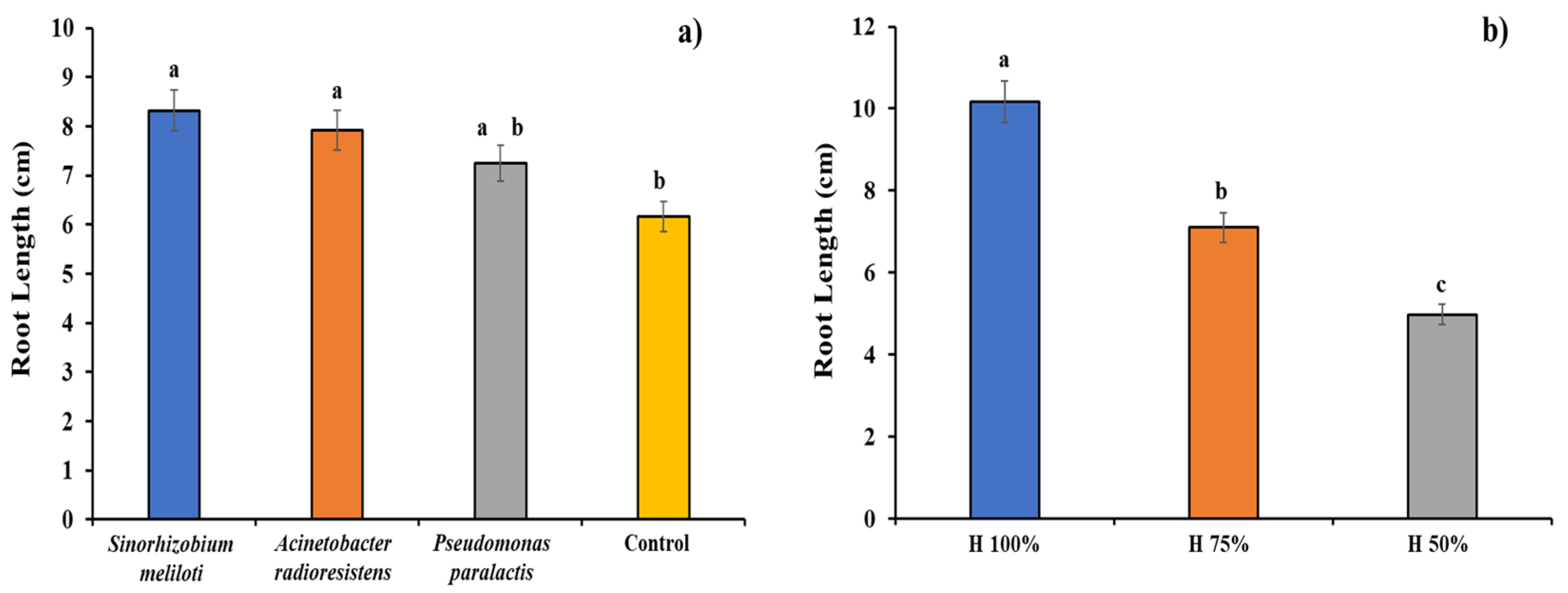
3.2. Root Length
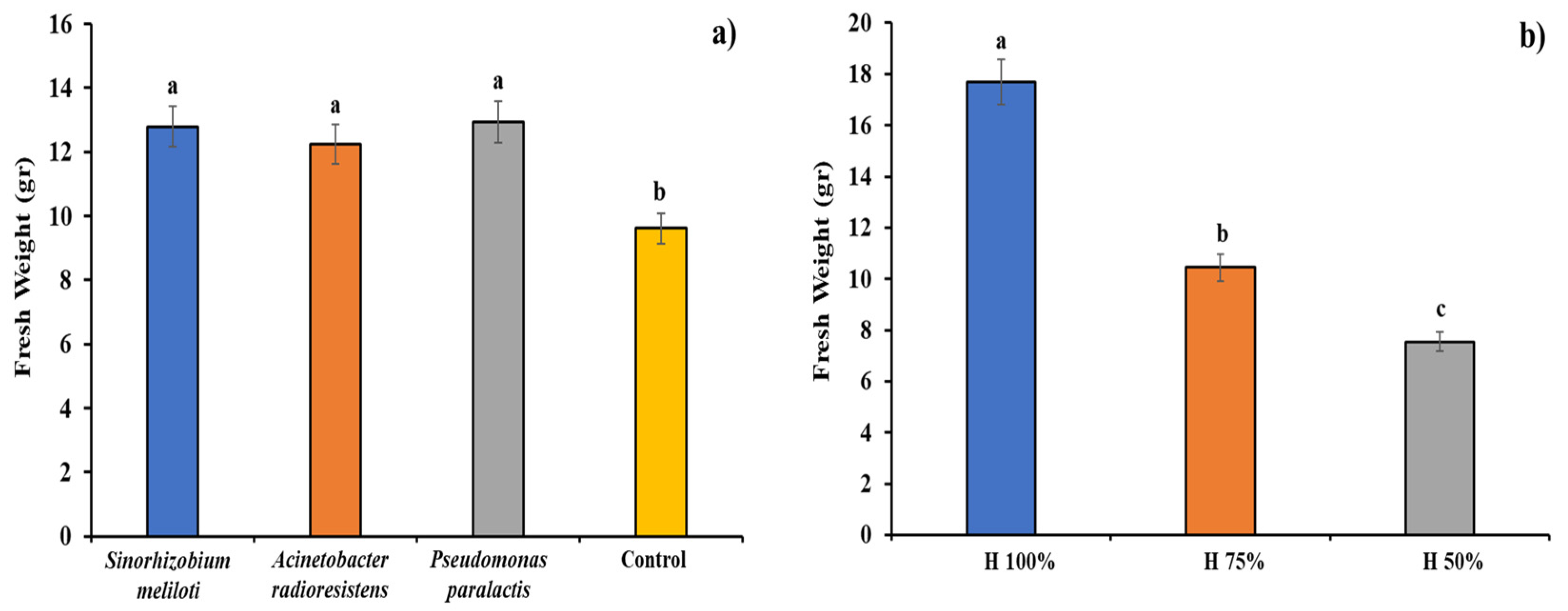
3.3. Fresh Weight
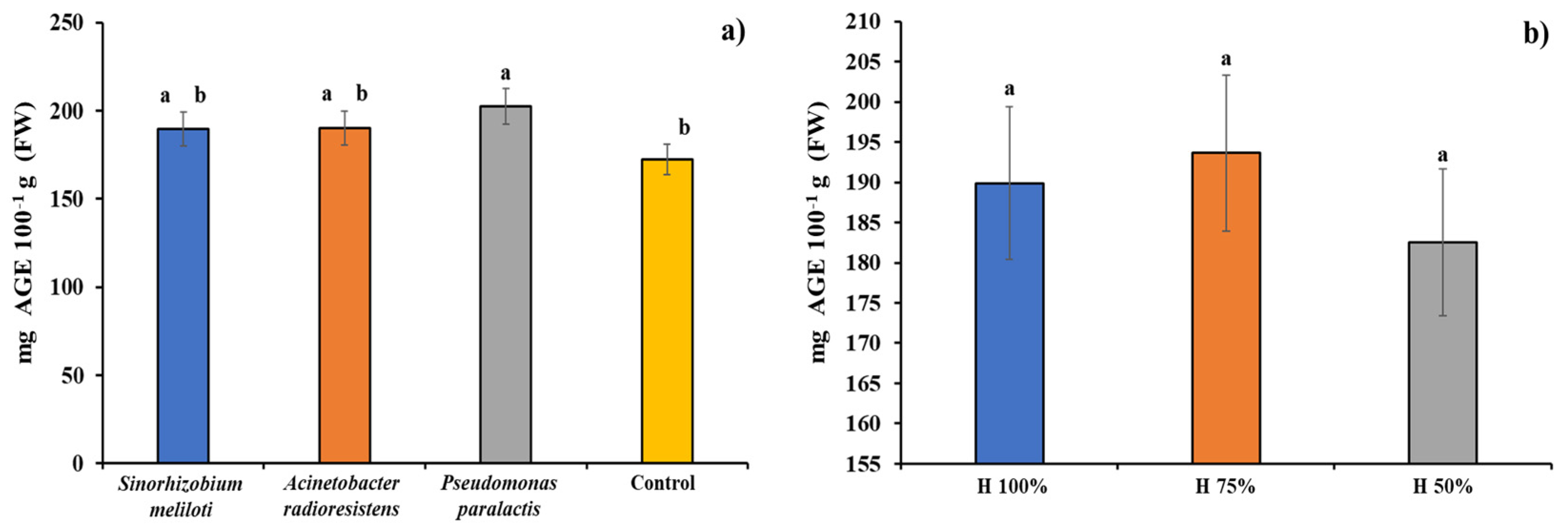
3.4. Total Phenols
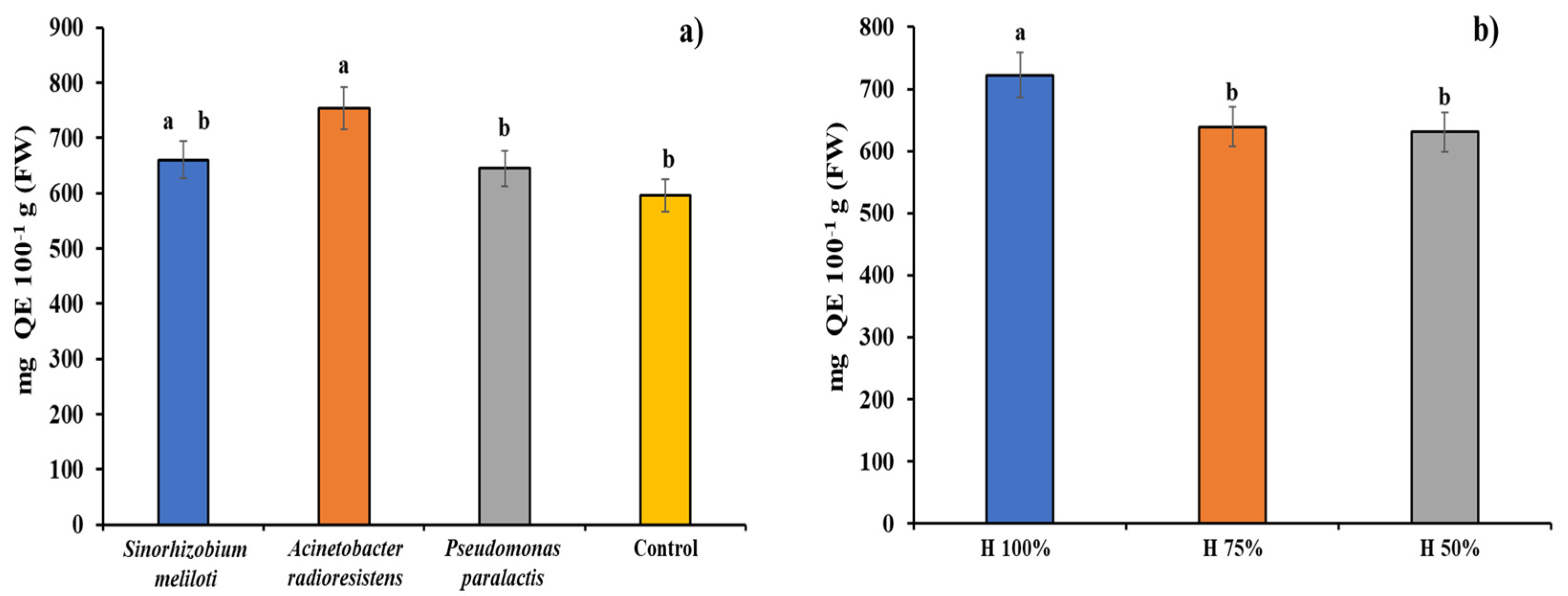
3.5. Flavonoids
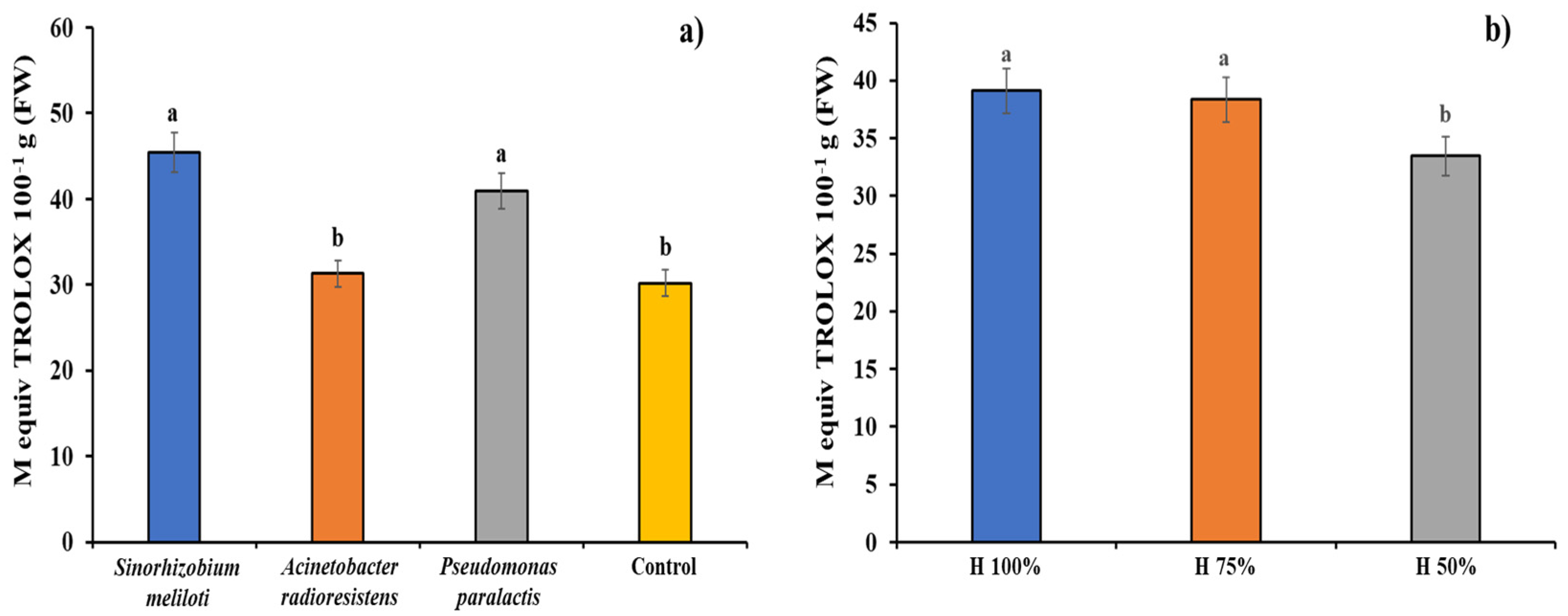
3.6. Antioxidant Capacity
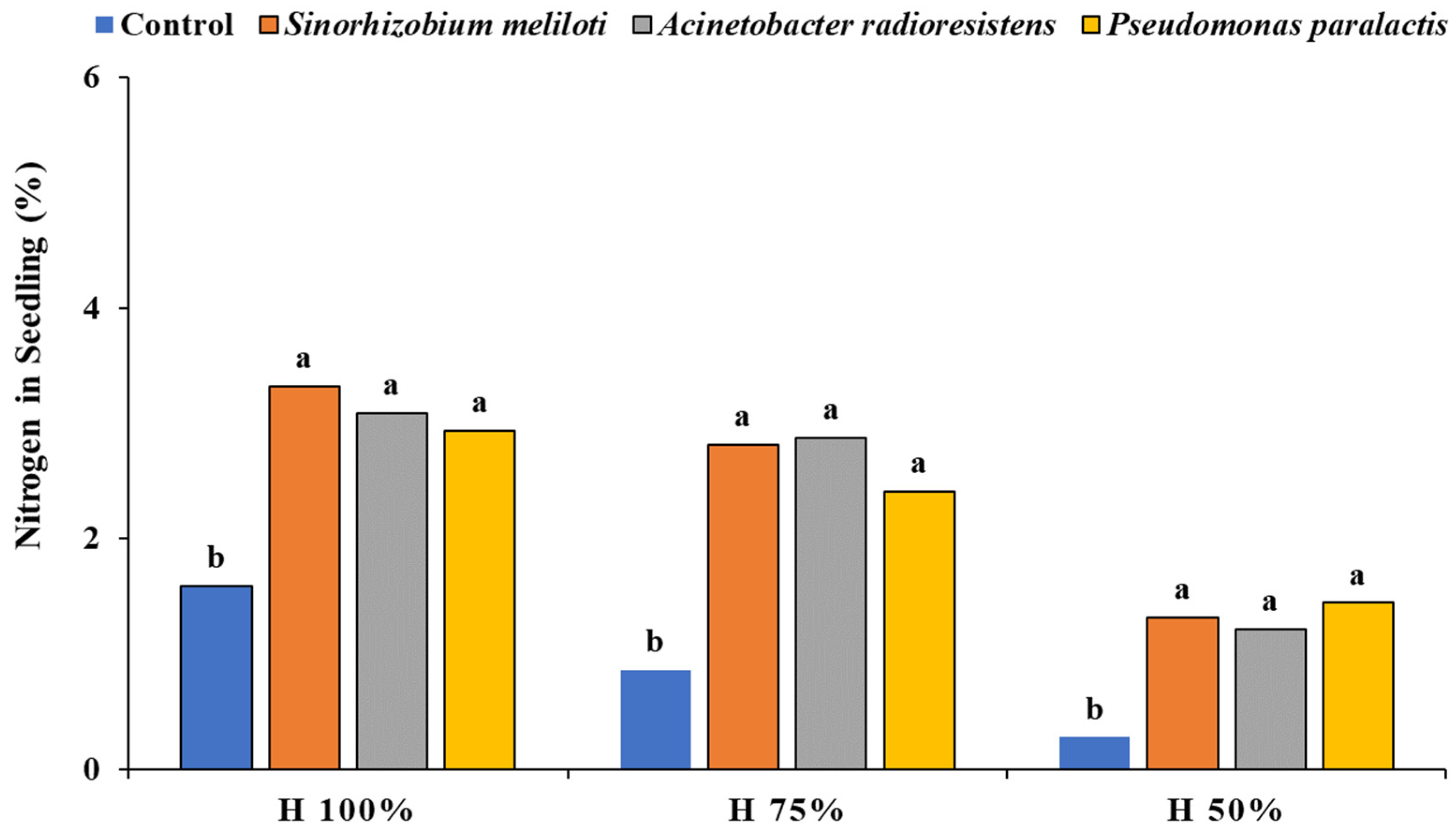
3.7. Nitrogen Content in Plant
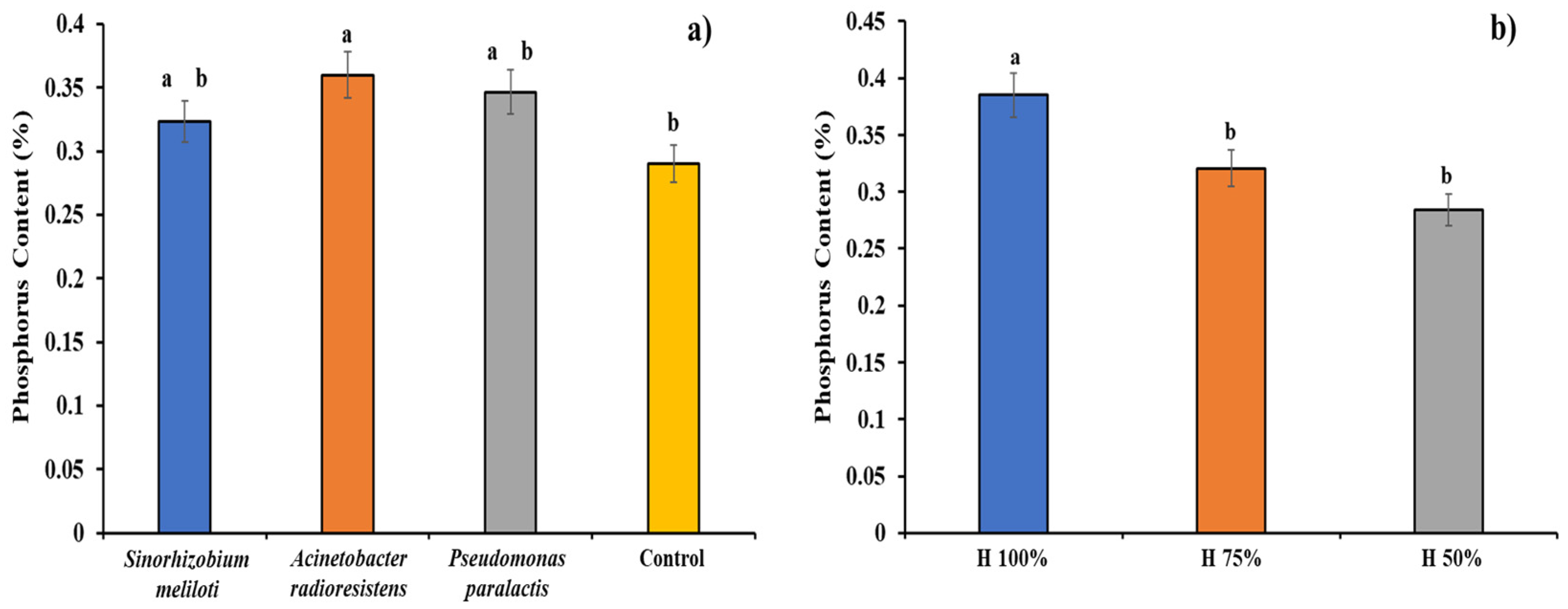
3.8. Phosphorus (P)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rai, A.; Chugh, V.; Pandey, S. Cucumber (Cucumis sativus L.): Genetic Improvement for Nutraceutical Traits. In Compendium of Crop Genome Designing for Nutraceuticals; Springer Nature: Singapore, 2023; pp. 1527–1544. [Google Scholar] [CrossRef]
- Aderinola, T.A.; Abaire, K.E. Quality acceptability, nutritional composition and antioxidant properties of carrot-cucumber juice. Beverages 2019, 5, 15. [Google Scholar] [CrossRef]
- Ji, F.; Wei, S.; Liu, N.; Xu, L.; Yang, P. Growth of cucumber seedlings in different varieties as affected by light environment. Int. J. Agric. Biol. Eng. 2020, 13, 73–78. [Google Scholar] [CrossRef]
- Roy, S.; Kapoor, R.; Mathur, P. Revisiting Changes in Growth, Physiology and Stress Responses of Plants under the Effect of Enhanced CO2 and Temperature. Plant Cell Physiol. 2024, 65, 4–19. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Oxygen and reactive oxygen species-dependent regulation of plant growth and development. Plant Physiol. 2021, 186, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Bagal, D.; Jain, D.; Singh, R.; Singh, I.K.; Singh, A. Biotic stresses on plants: Reactive oxygen species generation and antioxidant mechanism. In Frontiers in Plant-Soil Interaction; Academic Press: Cambridge, MA, USA, 2021; pp. 381–411. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Yadav, A.N. Bacterial Mitigation of Drought Stress in Plants: Current Perspectives and Future Challenges. Curr. Microbiol. 2022, 79, 248. [Google Scholar] [CrossRef] [PubMed]
- Yunusov, S.; Bolikulov, F.; Makhmanazarov, Z.; Yakhshiboev, D. Effect of growth regulators on cucumber seed fertility. E3S Web Conf. 2023, 421, 02013. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Singh, S.B.; Shilpa, N.; Aiyaz, M.; Nataraj, K.; Udayashankar, A.C.; Amruthesh, K.N.; Murali, M.; Poczai, P.; Gafur, A.; et al. Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants 2022, 11, 1763. [Google Scholar] [CrossRef]
- García-López, J.V.; Redondo-Gómez, S.; Flores-Duarte, N.J.; Zunzunegui, M.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Mateos-Naranjo, E. Exploring through the use of physiological and isotopic techniques the potential of a PGPR-based biofertilizer to improve nitrogen fertilization practices efficiency in strawberry cultivation. Front. Plant Sci. 2023, 14, 1243509. [Google Scholar] [CrossRef]
- Pranaw, K.; Kumawat, K.C.; Meena, V.S. Plant growth-promoting rhizobacteria (PGPR) and plant hormones: An approach for plant abiotic stress management and sustainable agriculture. Front. Microbiol. 2023, 14, 1285756. [Google Scholar] [CrossRef]
- Gul, A.; Nawaz, S. Role of phytohormones in biotic vs abiotic stresses with respect to PGPR and autophagy. In Phytohormones and Stress Responsive Secondary Metabolites; Academic Press: Cambridge, MA, USA, 2023; pp. 41–62. [Google Scholar]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Joshi, S.; Jaggi, V.; Tiwari, S.; Sah, V.K.; Sahgal, M. Multitrate phosphate solubilizing bacteria from Dalbergia sissoo Roxb. Rhizosphere in natural forests of Indian Central Himalayas. Environ. Ecol. 2019, 3, 894–908. [Google Scholar]
- Sharghi, A.; Badi, H.N.; Bolandnazar, S.; Mehrafarin, A.; Sarikhani, M.R. Morphophysiological and phytochemical responses of fenugreek to plant growth promoting rhizobacteria (PGPR) under different soil water levels. Folia Hortic. 2018, 30, 215–228. [Google Scholar] [CrossRef]
- Rosier, A.; Beauregard, P.B.; Bais, H.P. Quorum quenching activity of the PGPR Bacillus subtilis UD1022 alters nodulation efficiency of Sinorhizobium meliloti on Medicago truncatula. Front. Microbiol. 2021, 11, 596299. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, G.A. The role of salt-tolerant plant growth promoting bacteria in increasing the resistance of canola to salt-stress. J. Soil Sci. Agric. Eng. 2022, 13, 147–156. [Google Scholar] [CrossRef]
- Salazar-Ramírez, M.T.; Gallegos-López, D.A.; Hernández-Ordaz, G.; Preciado-Rangel, P.; Vidal, J.A.O. Inoculated growth promoting rhizobacteria in Serrano chili pepper (Capsicum annuum L.) seedlings under abiotic stress conditions. Agrociencia 2023, 57, 1–12. [Google Scholar]
- Akköprü, A.; Özaktan, H. Identification of rhizobacteria that increase yield and plant tolerance to angular leaf spot disease in cucumber. Plant Prot. Sci. 2018, 54, 67–73. [Google Scholar] [CrossRef]
- Carrillo, A.; Puente, M.; Castellanos, T.; Bashan, Y. Aplicaciones Biotecnológicas de Ecología Microbiana, Manual de Laboratorio. [Biotechnological Applications of Microbial Ecology. Laboratory Manual]; Pontificia Universidad Javeriana: Santa Fe de Bogotá, Colombia; Centro de Investigaciones Biológicas de Noroeste SC: La Paz, Mexico, 1998. [Google Scholar]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Villanueva, D.M.L. Eficacia de biofungicidas frente a la caída de plántula de pepino, inducida por Pythium aphanidermatum. Rev. Investig. Agroproducción Sustentable 2018, 2, 72–78. [Google Scholar] [CrossRef]
- Borges-Gómez, L.; Cervantes-Cárdenas, L.; Ruiz-Novelo, J.; Soria-Fregoso, M.; Reyes-Oregel, V.; Villanueva-Couoh, E. Capsaicinoides en chile habanero (Capsicum chinense Jacq.) bajo diferentes condiciones de humedad y nutrición. Terra Latinoam. 2010, 28, 35–41. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants using a folic-ciocalteu reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar]
- Lamaison, J.L.C.; Carnet, A. Contents in main flavonoid compounds of Crataegus Monogyna Jacq. and Crataegus laevigata (Poiret) D. C. flowers at different development stages. Pharm. Acta Helv. 1990, 65, 315–320. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bremmer, J.M.; Keeney, D.R. Steam distillation methods for determination of ammonium, nitrate and nitrite. Anal. Chim. Acta 1965, 32, 485–495. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus Soluble in Sodium Bicarbonate. In Methods of Soils Analysis. Part 2, Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA; Soil Science Society of America: Madison, WI, USA, 1982; pp. 404–430. [Google Scholar]
- Kour, D.; Rana, K.L.; Sheikh, I.; Kumar, V.; Yadav, A.N.; Dhaliwal, H.S.; Saxena, A.K. Alleviation of drought stress and plant growth promotion by Pseudomonas libanensis EU-LWNA-33, a drought-adaptive phosphorus-solubilizing bacterium. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 785–795. [Google Scholar] [CrossRef]
- Bouremani, N.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Baranov, O.; Belbahri, L. Plant growth-promoting rhizobacteria (PGPR): A rampart against the adverse effects of drought stress. Water 2023, 15, 418. [Google Scholar] [CrossRef]
- Ambreetha, S.; Chinnadurai, C.; Marimuthu, P.; Balachandar, D. Plant-associated Bacillus modulates the expression of auxin-responsive genes of rice and modifies the root architecture. Rhizosphere 2018, 5, 57–66. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, B.; Le Scornet, A.; Sauviac, L.; Rémy, A.; Bruand, C.; Meilhoc, E. The nitrate assimilatory pathway in Sinorhizobium meliloti: Contribution to NO production. Front. Microbiol. 2019, 10, 1526. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.; Hassan, S.E.D. Plant growth-promoting endophytic bacterial community inhabiting the leaves of Pulicaria incisa (Lam.) DC inherent to arid regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef]
- Calvo, P.; Zebelo, S.; McNear, D.; Kloepper, J.; Fadamiro, H. Plant growth-promoting rhizobacteria induce changes in Arabidopsis thaliana gene expression of nitrate and ammonium uptake genes. J. Plant Interact. 2019, 14, 224–231. [Google Scholar] [CrossRef]
- Ding, L.; Lu, Z.; Gao, L.; Guo, S.; Shen, Q. Is nitrogen a key determinant of water transport and photosynthesis in higher plants upon drought stress? Front. Plant Sci. 2018, 9, 1143. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Iqrar, I.; Shinwari, Z.K.; Yasmin, A. Plant growth-promoting potential of endophytic bacteria isolated from roots of wild Dodonaea viscosa L. Plant Growth Regul. 2017, 81, 399–408. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zapata-Sifuentes, G.; Fortis-Hernández, M.; Sáenz-Mata, J.; Silva-Martínez, C.; Lara-Capistran, L.; Preciado-Rangel, P.; Hernández-Montiel, L.G. Effect of Plant Growth Promoting Rhizobacteria on the Development and Biochemical Composition of Cucumber under Different Substrate Moisture Levels. Microbiol. Res. 2024, 15, 1505-1515. https://doi.org/10.3390/microbiolres15030102
Zapata-Sifuentes G, Fortis-Hernández M, Sáenz-Mata J, Silva-Martínez C, Lara-Capistran L, Preciado-Rangel P, Hernández-Montiel LG. Effect of Plant Growth Promoting Rhizobacteria on the Development and Biochemical Composition of Cucumber under Different Substrate Moisture Levels. Microbiology Research. 2024; 15(3):1505-1515. https://doi.org/10.3390/microbiolres15030102
Chicago/Turabian StyleZapata-Sifuentes, Gerardo, Manuel Fortis-Hernández, Jorge Sáenz-Mata, Christian Silva-Martínez, Liliana Lara-Capistran, Pablo Preciado-Rangel, and Luis Guillermo Hernández-Montiel. 2024. "Effect of Plant Growth Promoting Rhizobacteria on the Development and Biochemical Composition of Cucumber under Different Substrate Moisture Levels" Microbiology Research 15, no. 3: 1505-1515. https://doi.org/10.3390/microbiolres15030102
APA StyleZapata-Sifuentes, G., Fortis-Hernández, M., Sáenz-Mata, J., Silva-Martínez, C., Lara-Capistran, L., Preciado-Rangel, P., & Hernández-Montiel, L. G. (2024). Effect of Plant Growth Promoting Rhizobacteria on the Development and Biochemical Composition of Cucumber under Different Substrate Moisture Levels. Microbiology Research, 15(3), 1505-1515. https://doi.org/10.3390/microbiolres15030102