
Phenolic Compounds Synthesized by Trichoderma longibrachiatum Native to Semi-Arid Areas Show Antifungal Activity against Phytopathogenic Fungi of Horticultural Interest

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms Used and Culture Conditions
2.2. Obtaining the Crude Extract of T. longibrachiatum in Submerged Culture
2.3. Extraction of Phenolic Compounds from Extract of T. longibrachiatum
2.4. Quantification of Total Phenolics and Flavonoids
2.5. Determination of the Profile of Phenolic Compounds by HPLC
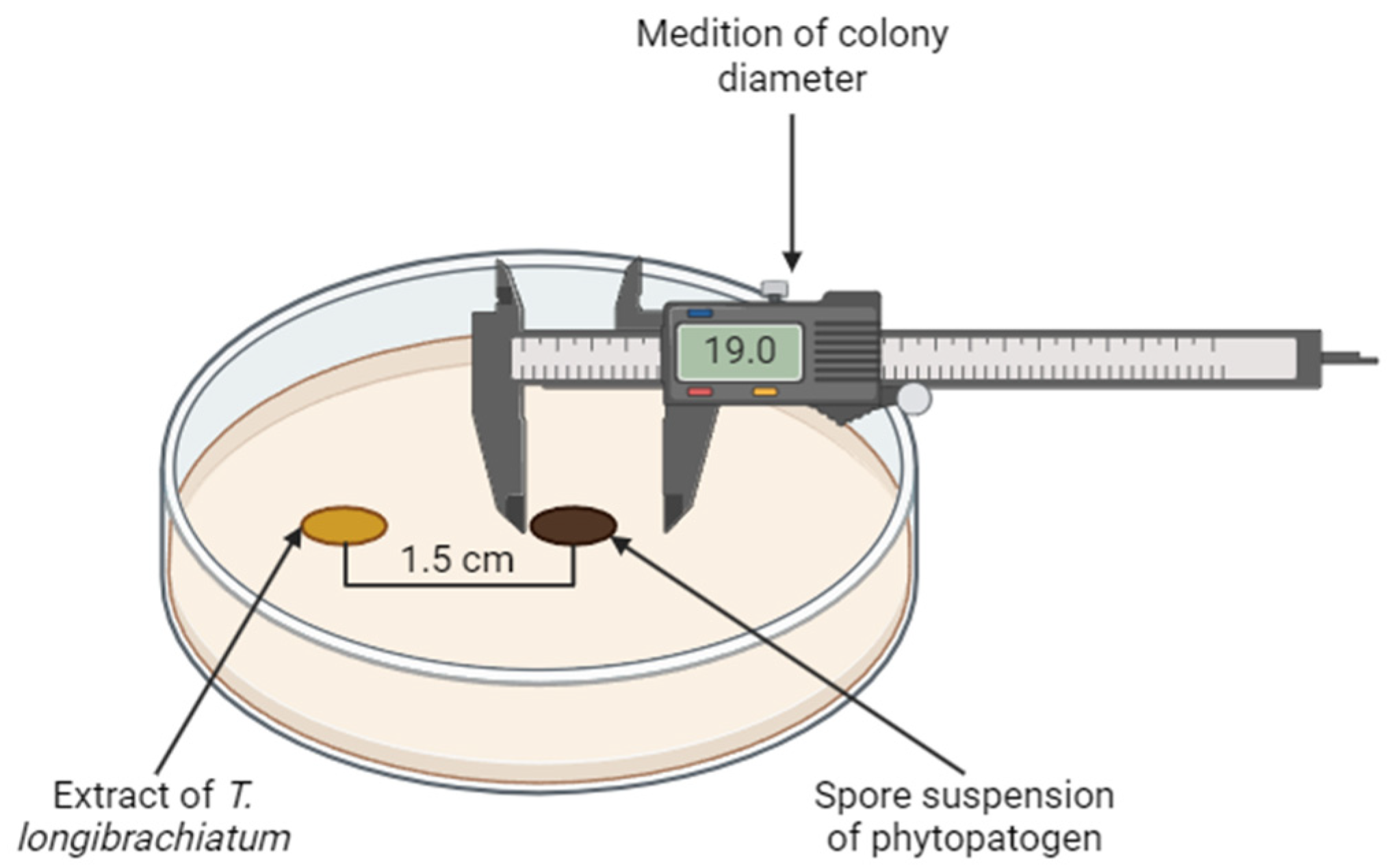
2.6. Antifungal Capacity of the Ethanolic Extract on Mycelial Growth
2.7. Antifungal Capacity of the Ethanolic Extract on Spore Germination
2.8. Antifungal Activity of Phenolic Acids Identified on Spore Germination
2.9. Statistical Analysis
3. Results
3.1. Effect of Extraction Method on the Concentration of Phenolic Compounds
3.2. Phenolic Profile of T. longibrachiatum Extract
3.3. Effect of the Ethanolic Extract of T. longibrachiatum on Mycelial Growth of A. alternata and F. oxysporum
3.4. Effect of the Ethanolic Extract of T. longibrachiatum on Spore Germination of A. alternata and F. oxysporum
3.5. Antifungal Activity of Phenolic Acids on Spore Germination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guzmán-Guzmán, P.; Kumar, A.; de los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.D.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma Species: Our Best Fungal Allies in the Biocontrol of Plant Diseases—A Review. Plants 2023, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Guo, T.; Wang, Y.; Wang, A.; Zhang, M. Carbendazim: Ecological risks, toxicities, degradation pathways and potential risks to human health. Chemosphere 2023, 314, 137723. [Google Scholar] [CrossRef] [PubMed]
- Kiiker, R.; Juurik, M.; Heick, T.M. Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020. Microorganisms 2021, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhou, R.; Zhu, G.; Xie, X.; Chai, A.; Li, L.; Fan, T.; Li, B.; Shi, Y. Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola. Microorganisms 2023, 11, 2849. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive Secondary Metabolites from Trichoderma spp. against Phytopathogenic Fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.R.; De Oliveira, F.S.; Bráz, A.F.; Oliveira, J.S.; Lima-Santos, J.; Dias, A.A.M. Unveiling the anticancer potential of the ethanolic extract from Trichoderma asperelloides. Front. Pharmacol. 2024, 15, 1398135. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Sahoo, S.A.; Barvkar, V.T.; Srivastava, A.K.; Mukherjee, P.K. Trichoderma virens exerts herbicidal effect on Arabidopsis thaliana via modulation of amino acid metabolism. Plant Sci. 2023, 332, 111702. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.G.; Kos, T.; Puškarić, J.; Vrandečić, K.; Benković-Lačić, T.; Brmež, M. Soil ecosystem functioning through interactions of nematodes and fungi Trichoderma sp. Sustainability 2024, 16, 2885. [Google Scholar] [CrossRef]
- Tang, P.; Huang, D.; Zheng, K.X.; Hu, D.; Dai, P.; Li, C.H.; Qin, S.Y.; Chen, G.D.; Yao, X.S.; Gao, H. Thirteen new peptaibols with antimicrobial activities from Trichoderma sp. Chin. J. Nat. Med. 2023, 21, 868–880. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Mendoza-Mendoza, A.; Zeilinger, S.; Horwitz, B.A. Mycoparasitism as a mechanism of Trichoderma-mediated suppression of plant diseases. Fungal Biol. Rev. 2022, 39, 15–33. [Google Scholar] [CrossRef]
- Keswani, C.; Bisen, K.; Chitara, M.; Sarma, B.; Singh, H. Exploring the role of secondary metabolites of Trichoderma in tripartite interaction with plant and pathogens. In Agro-Environmental Sustainability; Singh, J., Seneviratne, G., Eds.; Springer: Cham, Switzerland, 2017; pp. 63–79. [Google Scholar] [CrossRef]
- Molina, M.M.; Pereira, A.G.; García, N.M. Propuesta de rangos de Índice de Color según estados de maduración en frutas. Rev. Cienc. Técnicas Agropecu. 2019, 28, 19–27. [Google Scholar]
- Dal Bello, G.; Lampugnani, G.; Abramoff, C.; Fusé, C.; Perelló, A. Postharvest control of Botrytis gray mould in tomato by antagonists and biorational compounds. IOBC-WPRS 2015, 111, 417–425. [Google Scholar]
- Intana, W.; Kheawleng, S.; Sunpapao, A. Trichoderma asperellum T76-14 released volatile organic compounds against postharvest fruit rot in Muskmelons (Cucumis melo) caused by Fusarium incarnatum. J. Fungi 2021, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Ruangwong, O.-U.; Pornsuriya, C.; Pitija, K.; Sunpapao, A. Biocontrol mechanisms of Trichoderma koningiopsis PSU3-2 against postharvest anthracnose of chili pepper. J. Fungi 2021, 7, 276. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.A.; Najeeb, S.; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms 2020, 8, 401. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H. Antagonism of Trichoderma spp. against Macrophomina phaseolina: Evaluation of coiling and cell wall degrading enzymatic activities. J. Plant Pathol. Microbiol. 2012, 3, 149. [Google Scholar] [CrossRef]
- Al-Askar, A.A. Bioactive compounds produced by Trichoderma harzianum 1-SSR for controlling Fusarium verticillioides (Sacc.) nirenberg and growth promotion of Sorghum vulgare. Egypt. J. Biol. Pest Control. 2016, 26, 379–386. [Google Scholar]
- Tchameni, S.N.; Cotârleț, M.; Ghinea, I.O.; Bedine, M.A.B.; Sameza, M.L.; Borda, D.; Bahrim, G.; Dinică, R.M. Involvement of lytic enzymes and secondary metabolites produced by Trichoderma spp. in the biological control of Pythium myriotylum. Int. Microbiol. 2020, 23, 179–188. [Google Scholar] [CrossRef]
- Priego-Capote, F. 6—Solid–liquid extraction techniques. In Analytical Sample Preparation with Nano- and Other High-Performance Materials; Lucena, R., Cárdenas, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 111–130. [Google Scholar]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Soxhlet extraction of phenolic compounds from Vernonia cinerea leaves and its antioxidant activity. J. Appl. Res. Med. Aromat. Plants 2018, 11, 12–17. [Google Scholar] [CrossRef]
- Cablé, P.-A.; Le Brech, Y.; Mutelet, F. Liquid-liquid extraction of phenolic compounds from aqueous solution using hydrophobic deep eutectic solvents. J. Mol. Liq. 2022, 366, 120266. [Google Scholar] [CrossRef]
- Barp, L.; Višnjevec, A.M.; Moret, S. Pressurized liquid extraction: A powerful tool to implement extraction and purification of food contaminants. Foods 2023, 12, 2017. [Google Scholar] [CrossRef] [PubMed]
- Le, A.; Parks, S.; Nguyen, M.; Roach, P. Effect of solvents and extraction methods on recovery of bioactive compounds from defatted gac (Momordica cochinchinensis Spreng.) seeds. Separations 2018, 5, 39. [Google Scholar] [CrossRef]
- Socas-Rodríguez, B.; Torres-Cornejo, M.V.; Álvarez-Rivera, G.; Mendiola, J.A. Deep eutectic solvents for the extraction of bioactive compounds from natural sources and agricultural by-products. Appl. Sci. 2021, 11, 4897. [Google Scholar] [CrossRef]
- Cravotto, C.; Fabiano-Tixier, A.-S.; Claux, O.; Abert-Vian, M.; Tabasso, S.; Cravotto, G.; Chemat, F. Towards substitution of hexane as extraction solvent of food products and ingredients with no regrets. Foods 2022, 11, 3412. [Google Scholar] [CrossRef] [PubMed]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Ngo, M.T.; Nguyen, M.V.; Han, J.W.; Park, M.S.; Kim, H.; Choi, G.J. In vitro and in vivo antifungal activity of sorbicillinoids produced by Trichoderma longibrachiatum. J. Fungi 2021, 7, 428. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Gonzalez-Mendoza, D.; Zayed, O. Cell-free culture filtrate of Trichoderma longibrachiatum AD-1 as alternative approach to control Fusarium solani and induce defense response Phaseolus vulgaris L. plants. Rhizosphere 2023, 25, 100648. [Google Scholar] [CrossRef]
- Gonzalez-Soto, T.; Moreno-Ramírez, L.; Troncoso-Rojas, R.; Gonzalez-Mendoza, D.; Sánchez, A.; Grimaldo, O.; Camacho, O.; Ceceña-Duram, C. Inoculación de Trichoderma longibrachiatum en algodón transgénico: Cambios en compuestos fenólicos y enzimas de estrés oxidativo. Idesia 2017, 35, 19–24. [Google Scholar] [CrossRef]
- Cabral-Miramontes, J.P.; Olmedo-Monfil, V.; Lara-Banda, M.; Zúñiga-Romo, E.R.; Aréchiga-Carvajal, E.T. Promotion of plant growth in arid zones by selected Trichoderma spp. Strains with adaptation plasticity to alkaline ph. Biology 2022, 11, 1206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, B.; Zhang, J.; Gan, Y. Identification of the antifungal activity of Trichoderma longibrachiatum T6 and assessment of bioactive substances in controlling phytopathgens. Pestic. Biochem. Physiol. 2018, 147, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Angulo, J.C.; Mendez-Trujillo, V.; González-Mendoza, D.; Morales-Trejo, A.; Grimaldo-Juarez, O.; Cervantes-Díaz, L. A rapid and inexpensive method for isolation of total DNA from Trichoderma spp. (Hypocreaceae). Genet. Mol. Res. GMR 2012, 11, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Khan, S.A.; Waheed, U.; Raheel, M.; Khan, Z.; Alrefaei, A.F.; Alkhamis, H.H. Morphological and genetic characterization of Fusarium oxysporum and its management using weed extracts in cotton. J. King Saud Univ. Sci. 2021, 33, 101299. [Google Scholar] [CrossRef]
- Uysal-Morca, A.; Kinay-Teksür, P.; Egercï, Y. Morphological and phylogenetic identification of Botrytis cinerea causing blossom blight and fruit rot of sweet cherries in Aegean region, Turkey. J. Plant Dis. Prot. 2021, 128, 1051–1060. [Google Scholar] [CrossRef]
- He, L.; Cheng, H.; Zhao, L.; Htun, A.A.; Yu, Z.H.; Deng, J.X. Morphological and molecular identification of two new Alternaria species (Ascomycota, Pleosporaceae) in section Radicina from China. MycoKeys 2021, 78, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lopez, T.; Corbin, C.; Falguieres, A.; Doussot, J.; Montguillon, J.; Hagège, D.; Hano, C.; Lainé, É. Secondary metabolite accumulation, antibacterial and antioxidant properties of in vitro propagated Clidemia hirta L. extracts are influenced by the basal culture medium. Comptes Rendus Chim. 2016, 19, 1071–1076. [Google Scholar] [CrossRef]
- Kamali, H.; Khodaverdi, E.; Hadizadeh, F.; Ghaziaskar, S.H. Optimization of phenolic and flavonoid content and antioxidants capacity of pressurized liquid extraction from Dracocephalum kotschyi via circumscribed central composite. J. Supercrit. Fluids 2016, 107, 307–314. [Google Scholar] [CrossRef]
- Mradu, G.; Saumyakanti, S.; Sohini, M.; Arup, M. HPLC profiles of standard phenolic compounds present in medicinal plants. Int. J. Pharmacogn. Phytochem. Res. 2012, 4, 162–167. [Google Scholar]
- National Committee for Clinical Laboratory, Standards; Pfaller, M.A. Reference Method for Broth Dilution Antifungal Testing of Filamentous Fungi; NCCLS: Wayne, PA, USA, 2018. [Google Scholar]
- Irakli, M.N.; Samanidou, V.F.; Biliaderis, C.G.; Papadoyannis, I.N. Simultaneous determination of phenolic acids and flavonoids in rice using solid-phase extraction and RP-HPLC with photodiode array detection. J. Sep. Sci. 2012, 35, 1603–1611. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, S.A.; Ijaz, M.U.; Ullah, S.; Muhammad, Z.; Suleria, H.A.R.; Song, Y. Antioxidant activity of polyphenolic extracts of filamentous fungus Mucor circinelloides (WJ11): Extraction, characterization and storage stability of food emulsions. Food Biosci. 2020, 34, 100525. [Google Scholar] [CrossRef]
- Liberal, Â.; Cardoso, R.V.C.; Heleno, S.A.; Fernandes, Â.; Barros, L.; Martins, A. Phenolic Acids from Fungi. In Natural Secondary Metabolites: From Nature, through Science, to Industry; Carocho, M., Heleno, S.A., Barros, L., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 475–495. [Google Scholar]
- da Silva, M.H.R.; Cueva-Yesquén, L.G.; Júnior, S.B.; Garcia, V.L.; Sartoratto, A.; de Angelis, D.d.F.; de Angelis, D.A. Endophytic fungi from Passiflora incarnata: An antioxidant compound source. Arch. Microbiol. 2020, 202, 2779–2789. [Google Scholar] [CrossRef] [PubMed]
- Takó, M.; Kerekes, E.B.; Zambrano, C.; Kotogán, A.; Papp, T.; Krisch, J.; Vágvölgyi, C. Plant phenolics and phenolic-enriched extracts as antimicrobial agents against food-contaminating microorganisms. Antioxidants 2020, 9, 165. [Google Scholar] [CrossRef] [PubMed]
- El-Nagar, A.; Elzaawely, A.A.; Taha, N.A.; Nehela, Y. The antifungal activity of gallic acid and its derivatives against Alternaria solani, the causal agent of tomato early blight. Agronomy 2020, 10, 1402. [Google Scholar] [CrossRef]
- Ecevit, K.; Barros, A.A.; Silva, J.M.; Reis, R.L. Preventing microbial infections with natural phenolic compounds. Future Pharmacol. 2022, 2, 460–498. [Google Scholar] [CrossRef]
- Troncoso-Rojas, R.; Sánchez-Estrada, A.; Carvallo, T.; González-León, A.; Ojeda-Contreras, J.; Aguilar-Valenzuela, A.; Tiznado-Hernández, M.-E. A fungal elicitor enhances the resistance of tomato fruit to Fusarium oxysporum infection by activating the phenylpropanoid metabolic pathway. Phytoparasitica 2013, 41, 133–142. [Google Scholar] [CrossRef]
- Ruelas, C.; Tiznado-Hernández, M.E.; Sánchez-Estrada, A.; Robles-Burgueño, M.R.; Troncoso-Rojas, R. Changes in phenolic acid content during Alternaria alternata infection in tomato fruit. J. Phytopathol. 2006, 154, 236–244. [Google Scholar] [CrossRef]
- Peleg, M.; Corradini, M.G. Microbial growth curves: What the models tell us and what they cannot. Crit. Rev. Food Sci. Nutr. 2011, 51, 917–945. [Google Scholar] [CrossRef]
- Imran, M.; Abo-Elyousr, K.A.M.; Mousa, M.A.A.; Saad, M.M. Use of Trichoderma culture filtrates as a sustainable approach to mitigate early blight disease of tomato and their influence on plant biomarkers and antioxidants production. Front. Plant Sci. 2023, 14, 1192818. [Google Scholar] [CrossRef]
- Rauf, A.; Subhani, M.N.; Siddique, M.; Shahid, H.; Chattha, M.B.; Alrefaei, A.F.; Hasan Naqvi, S.A.; Ali, H.; Lucas, R.S. Cultivating a greener future: Exploiting trichoderma derived secondary metabolites for fusarium wilt management in peas. Heliyon 2024, 10, e29031. [Google Scholar] [CrossRef]
- Villao-Uzho, L.; Espinoza-Lozano, F.; Galarza-Romero, L.; Santos-Ordóñez, E. Biotechnological tools for genetic improvement of Trichoderma. Sci. Agropecu. 2024, 15, 213–223. [Google Scholar] [CrossRef]
- Zabka, M.; Pavela, R. Antifungal efficacy of some natural phenolic compounds against significant pathogenic and toxinogenic filamentous fungi. Chemosphere 2013, 93, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wu, J.; Wang, M.; Jiao, W.; Chen, Q.; Du, Y.; Chen, X.; Yang, X.; Fu, M. p-coumaric acid prevents Colletotrichum gloeosporioides by inhibiting membrane targeting and organic acid metabolism. Postharvest Biol. Technol. 2023, 204, 112447. [Google Scholar] [CrossRef]
- Yan, H.; Meng, X.; Lin, X.; Duan, N.; Wang, Z.; Wu, S. Antifungal activity and inhibitory mechanisms of ferulic acid against the growth of Fusarium graminearum. Food Biosci. 2023, 52, 102414. [Google Scholar] [CrossRef]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhao, P.; Ge, X.; Tian, P. Overview of Alternaria alternata Membrane Proteins. Indian J. Microbiol. 2020, 60, 269–282. [Google Scholar] [CrossRef] [PubMed]
- O’Mara, S.P.; Broz, K.; Schwister, E.M.; Singh, L.; Dong, Y.; Elmore, J.M.; Kistler, H.C. The Fusarium graminearum transporters abc1 and abc6 are important for xenobiotic resistance, trichothecene accumulation, and virulence to wheat. Phytopathology 2023, 113, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Mendoza, L.; Cotoras, M. Alteration of oxidative phosphorylation as a possible mechanism of the antifungal action of p-coumaric acid against Botrytis cinerea. J. Appl. Microbiol. 2017, 123, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Possamai Rossatto, F.C.; Tharmalingam, N.; Escobar, I.E.; d’Azevedo, P.A.; Zimmer, K.R.; Mylonakis, E. Antifungal activity of the phenolic compounds ellagic acid (ea) and caffeic acid phenethyl ester (cape) against drug-resistant Candida auris. J. Fungi 2021, 7, 763. [Google Scholar] [CrossRef]
- Soto, M.; Estevez-Braun, A.; Amesty, A.; Kluepfel, J.; Restrepo, S.; Diaz, K.; Espinoza, L.; Olea, A.F.; Taborga, L. Synthesis and fungicidal activity of hydrated geranylated phenols against Botrytis cinerea. Molecules 2021, 26, 6815. [Google Scholar] [CrossRef]
- Shirai, A.; Kawasaka, K.; Tsuchiya, K. Antimicrobial action of phenolic acids combined with violet 405-nm light for disinfecting pathogenic and spoilage fungi. J. Photochem. Photobiol. B Biol. 2022, 229, 112411. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytopathogen | Treatment | µmax (mm/h) * | R2 | p Value |
|---|---|---|---|---|
| A. alternata | Control | 0.39 ± 0.02 a | 0.9912 | 0.0001 |
| Extract | 0.24 ± 0.00 b | 0.9913 | ||
| F. oxysporum | Control | 0.41 ± 0.00 a | 0.9898 | 0.0001 |
| Extract | 0.28 ± 0.00 b | 0.9985 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-García, E.; Valenzuela-Quintanar, A.I.; Sánchez-Estrada, A.; González-Mendoza, D.; Tiznado-Hernández, M.E.; Islas-Rubio, A.R.; Troncoso-Rojas, R. Phenolic Compounds Synthesized by Trichoderma longibrachiatum Native to Semi-Arid Areas Show Antifungal Activity against Phytopathogenic Fungi of Horticultural Interest. Microbiol. Res. 2024, 15, 1425-1440. https://doi.org/10.3390/microbiolres15030096
Díaz-García E, Valenzuela-Quintanar AI, Sánchez-Estrada A, González-Mendoza D, Tiznado-Hernández ME, Islas-Rubio AR, Troncoso-Rojas R. Phenolic Compounds Synthesized by Trichoderma longibrachiatum Native to Semi-Arid Areas Show Antifungal Activity against Phytopathogenic Fungi of Horticultural Interest. Microbiology Research. 2024; 15(3):1425-1440. https://doi.org/10.3390/microbiolres15030096
Chicago/Turabian StyleDíaz-García, Enis, Ana Isabel Valenzuela-Quintanar, Alberto Sánchez-Estrada, Daniel González-Mendoza, Martín Ernesto Tiznado-Hernández, Alma Rosa Islas-Rubio, and Rosalba Troncoso-Rojas. 2024. "Phenolic Compounds Synthesized by Trichoderma longibrachiatum Native to Semi-Arid Areas Show Antifungal Activity against Phytopathogenic Fungi of Horticultural Interest" Microbiology Research 15, no. 3: 1425-1440. https://doi.org/10.3390/microbiolres15030096
APA StyleDíaz-García, E., Valenzuela-Quintanar, A. I., Sánchez-Estrada, A., González-Mendoza, D., Tiznado-Hernández, M. E., Islas-Rubio, A. R., & Troncoso-Rojas, R. (2024). Phenolic Compounds Synthesized by Trichoderma longibrachiatum Native to Semi-Arid Areas Show Antifungal Activity against Phytopathogenic Fungi of Horticultural Interest. Microbiology Research, 15(3), 1425-1440. https://doi.org/10.3390/microbiolres15030096