
Dietary Administration Effects of Microbial Exopolysaccharide from Bacillus subtilis P1 on Growth Performance and Immunity in Nile Tilapia (Oreochromis niloticus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening and Isolation of EPS-Producing Bacteria from Freshwater Scaled Fish Intestine
2.2. Exopolysaccharide Production, Purification, and Quantification
2.3. EPS Characterization of the Purified EPS
2.3.1. Molecular Mass Determination
2.3.2. Functional Groups Determination
2.3.3. Monosaccharide and Oligosaccharide Determination
2.3.4. Scanning Electron Microscopy (SEM) Analysis
2.4. Optimization Condition of EPS Production from the Selected Strain
2.5. Bacterial Identification
2.6. In Vitro Biological Activity
2.6.1. Prebiotic Properties
2.6.2. EPS Antioxidant Studies
2.6.3. Antibacterial Activity Studies
2.6.4. Cytotoxicity of EPS by Hemolytic Test
2.7. Experimental Animal
2.7.1. Acclimatization of Oreochromis niloticus Linn.
2.7.2. Experimental Fish Diet Formulation
2.7.3. Experimental Design and Sample Collection
2.8. Growth Performance Parameters
2.9. Measurement of Immune Response Parameters
2.9.1. Lysozyme Activity
2.9.2. Respiratory Burst Activity
2.9.3. Total Immunoglobulin
2.10. Measurement of Serum Biochemical Parameters
2.11. Challenge Test
2.12. Statistical Analysis
3. Results and Discussion
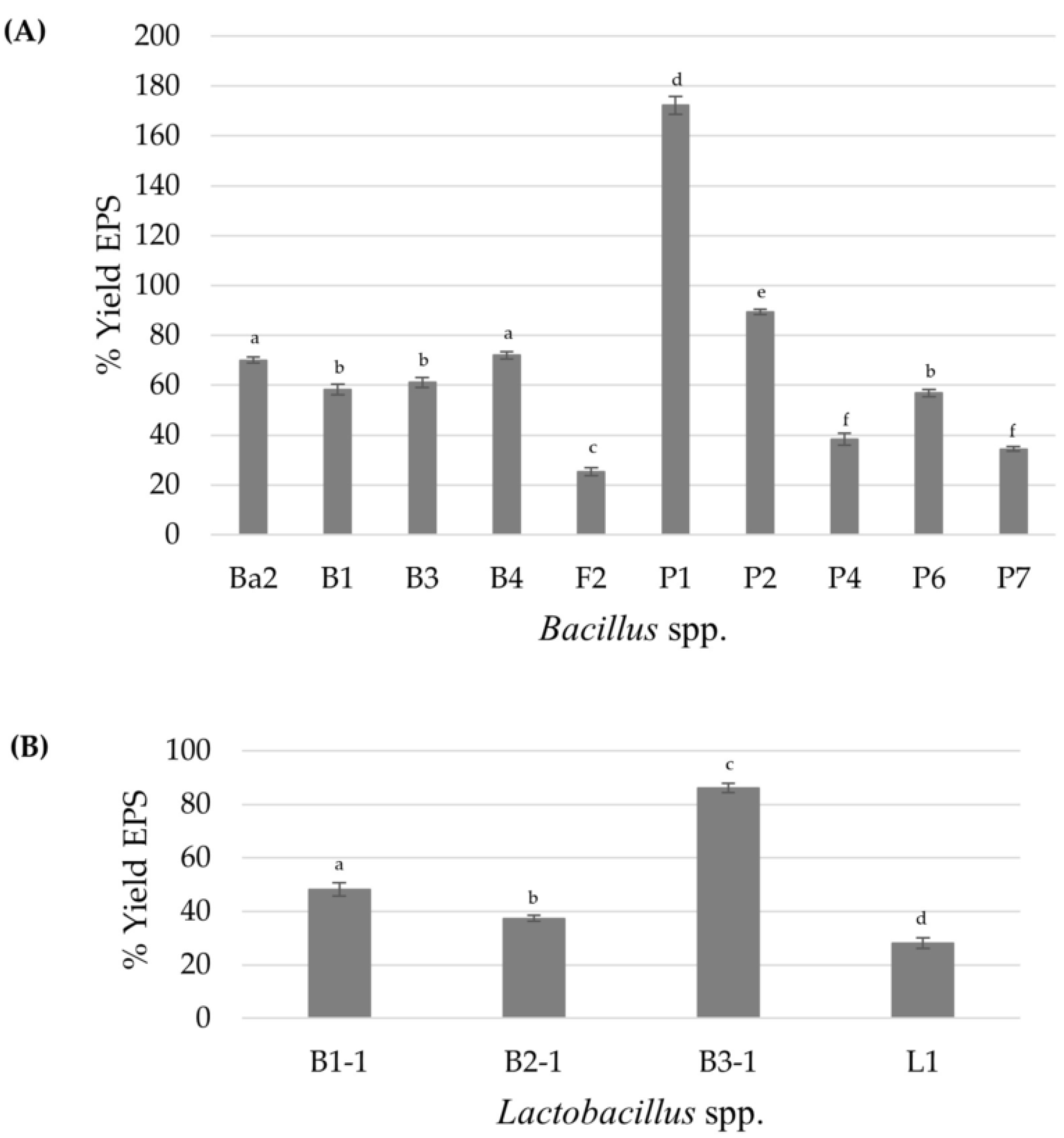
3.1. Screening and Isolation of EPS-Producing Bacteria
3.2. Characterization of the Purified EPS
3.2.1. Molecular Mass
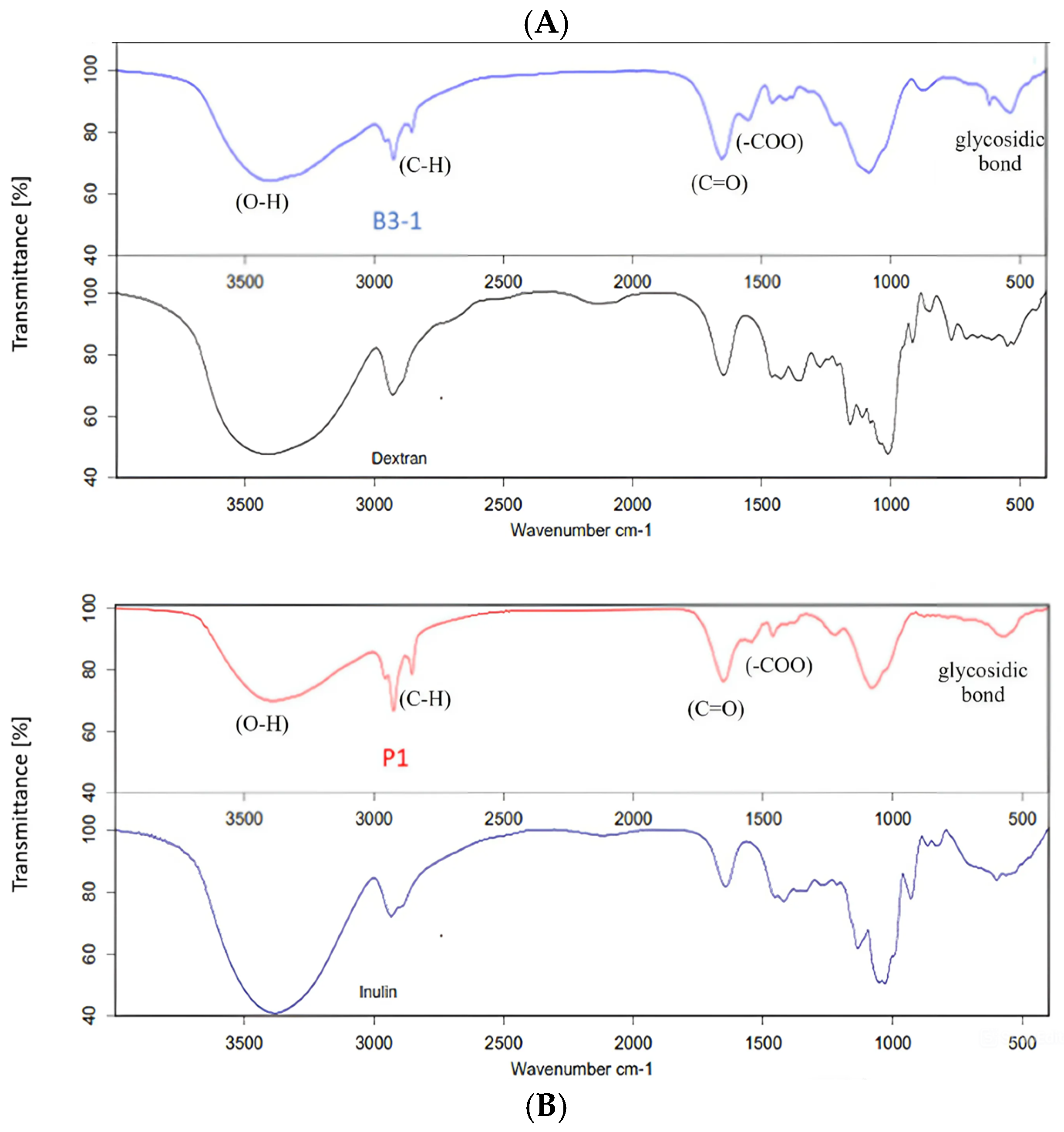
3.2.2. Functional Groups of EPS and Its Monosaccharide Compositions
3.3. Optimization Conditions for EPS Production
3.4. Bacterial Identification
3.5. In Vitro Biological Activity
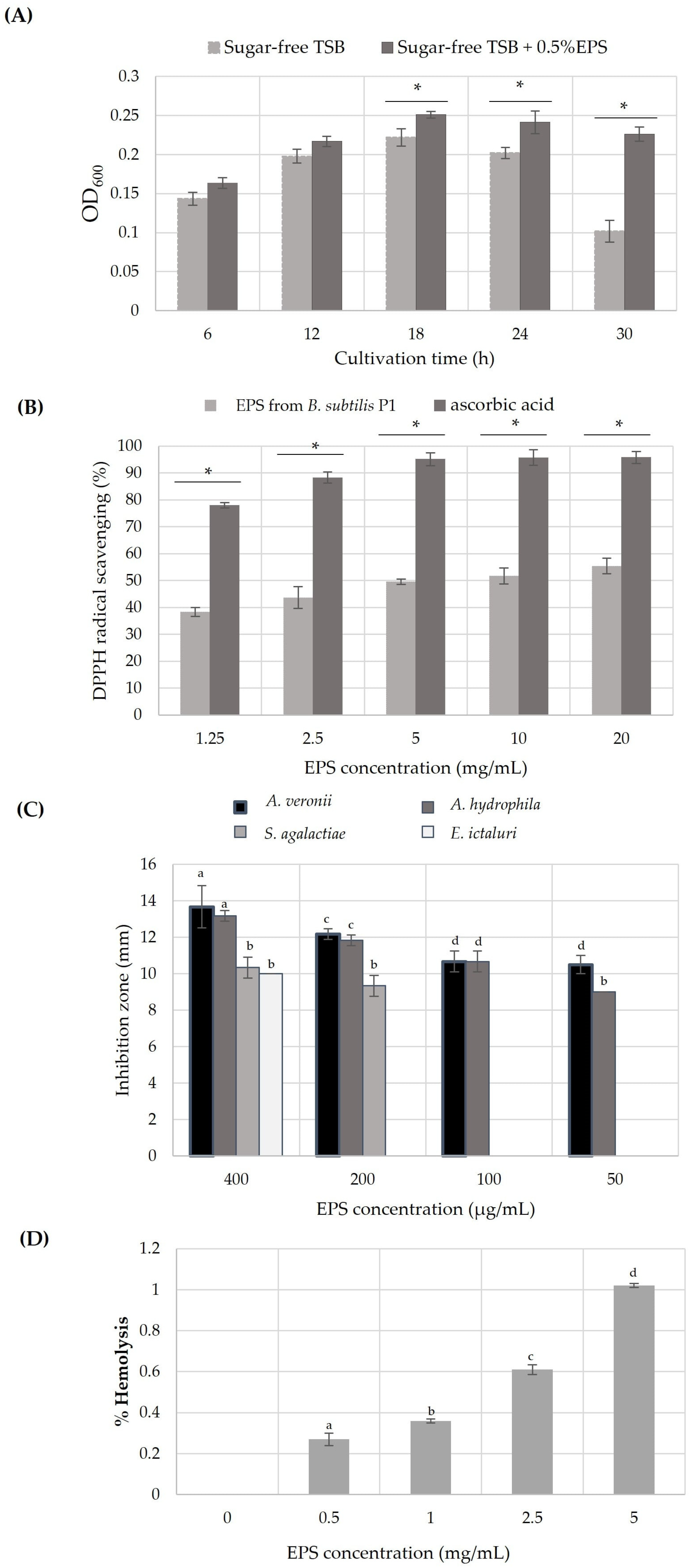
3.5.1. Prebiotic Properties of the Produced EPS
3.5.2. Anti-Oxidant Activity
3.5.3. Anti-Bacterial Activity
3.5.4. Cytotoxicity Test
3.6. In Vivo Study of the Effect of EPS Supplementation in Fish Diet on Growth Performance and Immune Response
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lertwanakarn, T.; Purimayata, T.; Luengyosluechakul, T.; Grimalt, P.B.; Pedrazzani, A.S.; Quintiliano, M.H.; Surachetpong, W. Assessment of Tilapia (Oreochromis spp.) Welfare in the Semi-Intensive and Intensive Culture Systems in Thailand. Animals 2023, 13, 2498. [Google Scholar] [CrossRef] [PubMed]
- Ibrahem, M.D.; Fathi, M.; Mesalhy, S.; El-Aty, A.A. Effect of dietary supplementation of inulin and vitamin C on the growth, hematology, innate immunity, and resistance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2010, 29, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.; Faggio, C. Importance of prebiotics in aquaculture as immunostimulants. Effects on immune system of Sparus aurata and Dicentrarchus labrax. Fish Shellfish Immunol. 2016, 54, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Esteban, M.Á.; Cuesta, A.; Sun, Y.-Z. Prebiotics and fish immune response: A review of current knowledge and future perspectives. Rev. Fish. Sci. Aquac. 2015, 23, 315–328. [Google Scholar] [CrossRef]
- Yousefian, M.; Amiri, M.S. A review of the use of prebiotic in aquaculture for fish and shrimp. Afr. J. Biotechnol. 2009, 8, 1–6. [Google Scholar]
- Camacho-Chab, J.C.; Lango-Reynoso, F.; Castañeda-Chávez, M.D.R.; Galaviz-Villa, I.; Hinojosa-Garro, D.; Ortega-Morales, B.O. Implications of extracellular polymeric substance matrices of microbial habitats associated with coastal aquaculture systems. Water 2016, 8, 369. [Google Scholar] [CrossRef]
- Gupta, S.; Das, P.; Singh, S.; Akhtar, M.; Meena, D.; Mandal, S. Microbial levari, an ideal prebiotic and immunonutrient in aquaculture. World Aquac. 2011, 42, 61. [Google Scholar]
- You, S.; Ma, Y.; Yan, B.; Pei, W.; Wu, Q.; Ding, C.; Huang, C. The promotion mechanism of prebiotics for probiotics: A review. Front. Nutr. 2022, 9, 1000517. [Google Scholar] [CrossRef]
- Netrusov, A.I.; Liyaskina, E.V.; Kurgaeva, I.V.; Liyaskina, A.U.; Yang, G.; Revin, V.V. Exopolysaccharides Producing Bacteria: A Review. Microorganisms 2023, 11, 1541. [Google Scholar] [CrossRef]
- Salimi, F.; Farrokh, P. Recent advances in the biological activities of microbial exopolysaccharides. World J. Microbiol. Biotechnol. 2023, 39, 213. [Google Scholar] [CrossRef]
- Wang, W.; Ju, Y.; Liu, N.; Shi, S.; Hao, L. Structural characteristics of microbial exopolysaccharides in association with their biological activities: A review. Chem. Biol. Technol. Agric. 2023, 10, 137. [Google Scholar] [CrossRef]
- Abinaya, M.; Shanthi, S.; Palmy, J.; Al-Ghanim, K.A.; Govindarajan, M.; Vaseeharan, B. Exopolysaccharides-mediated ZnO nanoparticles for the treatment of aquatic diseases in freshwater fish. Oreochromis mossambicus. Toxics 2023, 11, 313. [Google Scholar] [CrossRef]
- Sutthi, N.; Wangkahart, E.; Panase, P.; Karirat, T.; Deeseenthum, S.; Ma, N.L.; Luang-In, V. Dietary Administration Effects of Exopolysaccharide Produced by Bacillus tequilensis PS21 Using Riceberry Broken Rice, and Soybean Meal on Growth Performance, Immunity, and Resistance to Streptococcus agalactiae of Nile tilapia (Oreochromis niloticus). Animals 2023, 13, 3262. [Google Scholar] [CrossRef] [PubMed]
- Ruas-Madiedo, P.; Hugenholtz, J.; Zoon, P. An overview of the functionality of exopolysaccharides produced by lactic acid bacteria. Int. Dairy J. 2002, 12, 163–171. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2019, 487, 107881. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Mahdhi, A.; Chakroun, I.; Espinosa-Ruiz, C.; Messina, C.M.; Arena, R.; Majdoub, H.; Santulli, A.; Mzoughi, R.; Esteban, M.A. Dietary administration effects of exopolysaccharide from potential probiotic strains on immune and antioxidant status and nutritional value of European sea bass (Dicentrarchus labrax L.). Res. Vet. Sci. 2020, 131, 51–58. [Google Scholar] [CrossRef]
- Gobi, N.; Vaseeharan, B.; Chen, J.-C.; Rekha, R.; Vijayakumar, S.; Anjugam, M.; Iswarya, A. Dietary supplementation of probiotic Bacillus licheniformis Dahb1 improves growth performance, mucus and serum immune parameters, antioxidant enzyme activity as well as resistance against Aeromonas hydrophila in tilapia Oreochromis mossambicus. Fish Shellfish Immunol. 2018, 74, 501–508. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Jiang, Y.-Y.; Zhao, X.; Na Hao, X.; Li, L.; Yang, Z.-N. Characterization and antioxidant activity of the exopolysaccharide produced by Bacillus amyloliquefaciens GSBa-1. J. Microbiol. Biotechnol. 2018, 28, 1282–1292. [Google Scholar] [CrossRef]
- Petrova, P.; Arsov, A.; Ivanov, I.; Tsigoriyna, L.; Petrov, K. New Exopolysaccharides produced by Bacillus licheniformis 24 display substrate-dependent content and antioxidant activity. Microorganisms 2021, 9, 2127. [Google Scholar] [CrossRef]
- Moghannem, S.A.; Farag, M.M.; Shehab, A.M.; Azab, M.S. Exopolysaccharide production from Bacillus velezensis KY471306 using statistical experimental design. Braz. J. Microbiol. 2018, 49, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Berekaa, M.M. Improved exopolysaccharide production by Bacillus licheniformis strain-QS5 and application of statistical experimental design. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 876–886. [Google Scholar]
- Gangalla, R.; Sampath, G.; Beduru, S.; Sarika, K.; Govindarajan, R.K.; Ameen, F.; Alwakeel, S.; Thampu, R.K. Optimization and characterization of exopolysaccharide produced by Bacillus aerophilus rk1 and its in vitro antioxidant activities. J. King Saud Univ. Sci. 2021, 33, 101470. [Google Scholar] [CrossRef]
- Razack, S.A.; Velayutham, V.; Thangavelu, V. Medium optimization for the production of exopolysaccharide by Bacillus subtilis using synthetic sources and agro wastes. Turk. J. Biol. 2013, 37, 280–288. [Google Scholar] [CrossRef]
- Mgomi, F.C.; Yang, Y.-R.; Cheng, G.; Yang, Z.-Q. Lactic acid bacteria biofilms and their antimicrobial potential against pathogenic microorganisms. Biofilm 2023, 5, 100118. [Google Scholar] [CrossRef]
- Lee, M.-G.; Joeng, H.; Shin, J.; Kim, S.; Lee, C.; Song, Y.; Lee, B.-H.; Park, H.-G.; Lee, T.-H.; Jiang, H.-H.; et al. Potential probiotic properties of exopolysaccharide-producing Lacticaseibacillus paracasei EPS DA-BACS and prebiotic activity of its exopolysaccharide. Microorganisms 2022, 10, 2431. [Google Scholar] [CrossRef]
- Lim, E.S.; Nam, S.J.; Koo, O.K.; Kim, J.-S. Protective role of Acinetobacter and Bacillus for Escherichia coli O157:H7 in biofilms against sodium hypochlorite and extracellular matrix-degrading enzymes. Food Microbiol. 2023, 109, 104125. [Google Scholar] [CrossRef]
- Todhanakasem, T.; Sangsutthiseree, A.; Areerat, K.; Young, G.M.; Thanonkeo, P. Biofilm production by Zymomonas mobilis enhances ethanol production and tolerance to toxic inhibitors from rice bran hydrolysate. New Biotechnol. 2014, 31, 451–459. [Google Scholar] [CrossRef]
- Liu, C.; Lu, J.; Lu, L.; Liu, Y.; Wang, F.; Xiao, M. Isolation, structural characterization and immunological activity of an exopolysaccharide produced by Bacillus licheniformis 8-37-0-1. Bioresour. Technol. 2010, 101, 5528–5533. [Google Scholar] [CrossRef]
- Lin, T.; Chen, C.; Chen, B.; Shaw, J.; Chen, Y. Optimal economic productivity of exopolysaccharides from lactic acid bacteria with production possibility curves. Food Sci. Nutr. 2019, 7, 2336–2344. [Google Scholar] [CrossRef]
- Ziadi, M.; Bouzaiene, T.; M’hIr, S.; Zaafouri, K.; Mokhtar, F.; Hamdi, M.; Boisset-Helbert, C. Evaluation of the efficiency of ethanol precipitation and ultrafiltration on the purification and characteristics of exopolysaccharides produced by three lactic acid bacteria. BioMed Res. Int. 2018, 2018, 1896240. [Google Scholar] [CrossRef] [PubMed]
- Khongkool, K.; Prakit, B.; Chiyod, R.; Suttibul, T.; Lertworapreecha, M. Qualitative analysis of fibre-degrading enzymes production by Bacillus isolated from native swine manures. Burapha Sci. J. 2023, 28, 647–659. [Google Scholar]
- Yılmaz, T.; Şimşek, Ö. Potential health benefits of ropy exopolysaccharides produced by Lactobacillus plantarum. Molecules 2020, 25, 3293. [Google Scholar] [CrossRef] [PubMed]
- Zaghloul, E.H.; Ibrahim, M.I.A.; Zaghloul, H.A.H. Antibacterial activity of exopolysaccharide produced by bee gut-resident Enterococcus sp. BE11 against marine fish pathogens. BMC Microbiol. 2023, 23, 231. [Google Scholar] [CrossRef] [PubMed]
- Wanja, D.W.; Mbuthia, P.G.; Waruiru, R.M.; Bebora, L.C.; Ngowi, H.A.; Nyaga, P.N. Antibiotic and disinfectant susceptibility patterns of bacteria isolated from farmed fish in Kirinyaga County, Kenya. Int. J. Microbiol. 2020, 2020, 8897338. [Google Scholar] [CrossRef]
- Abinaya, M.; Vaseeharan, B.; Divya, M.; Vijayakumar, S.; Govindarajan, M.; Alharbi, N.S.; Khaled, J.M.; Al-Anbr, M.N.; Benelli, G. Structural characterization of Bacillus licheniformis Dahb1 exopolysaccharide—Antimicrobial potential and larvicidal activity on malaria and Zika virus mosquito vectors. Environ. Sci. Pollut. Res. 2018, 25, 18604–18619. [Google Scholar] [CrossRef]
- Figueiredo-Silva, C.; Lemme, A.; Sangsue, D.; Kiriratnikom, S. Effect of DL-methionine supplementation on the success of almost total replacement of fish meal with soybean meal in diets for hybrid tilapia (Oreochromis niloticus × Oreochromis mossambicus). Aquac. Nutr. 2015, 21, 234–241. [Google Scholar] [CrossRef]
- Biller, J.D.; Polycarpo, G.D.V.; Moromizato, B.S.; Sidekerskis, A.P.D.; da Silva, T.D.; dos Reis, I.C.; Fierro-Castro, C. Lysozyme activity as an indicator of innate immunity of tilapia (Oreochromis niloticus) when challenged with LPS and Streptococcus agalactiae. Rev. Bras. Zootec. 2021, 50, e20210053. [Google Scholar] [CrossRef]
- Stasiak, S.A.; Baumann, P.C. Neutrophil activity as a potential bioindicator for contaminant analysis. Fish Shellfish Immunol. 1996, 6, 537–539. [Google Scholar] [CrossRef]
- Hampton, L.M.T.; Jeffries, M.K.S.; Venables, B.J. A practical guide for assessing respiratory burst and phagocytic cell activity in the fathead minnow, an emerging model for immunotoxicity. MethodsX 2020, 7, 100992. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Acad. Bras. Cienc. 2018, 90, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Sewaka, M.; Trullas, C.; Chotiko, A.; Rodkhum, C.; Chansue, N.; Boonanuntanasarn, S.; Pirarat, N. Efficacy of synbiotic Jerusalem artichoke and Lactobacillus rhamnosus GG-supplemented diets on growth performance, serum biochemical parameters, intestinal morphology, immune parameters and protection against Aeromonas veronii in juvenile red tilapia (Oreochromis spp.). Fish Shellfish Immunol. 2019, 86, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Nuryastuti, T. Current in vitro assay to determine bacterial biofilm formation of clinical isolates. J. Med. Sci. Berk. Ilmu Kedokt. 2014, 46, 142–152. [Google Scholar] [CrossRef]
- Amao, J.A.; Omojasola, P.F.; Barooah, M. Isolation and characterization of some exopolysaccharide producing bacteria from cassava peel heaps. Sci. Afr. 2019, 4, e00093. [Google Scholar] [CrossRef]
- Marvasi, M.; Visscher, P.T.; Martinez, L.C. Exopolymeric substances (EPS) from Bacillus subtilis: Polymers and genes encoding their synthesis. FEMS Microbiol. Lett. 2010, 313, 1–9. [Google Scholar] [CrossRef]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef]
- Sánchez-León, E.; Huang-Lin, E.; Amils, R.; Abrusci, C. Production and characterisation of an exopolysaccharide by Bacillus amyloliquefaciens: Biotechnological applications. Polymers 2023, 15, 1550. [Google Scholar] [CrossRef]
- Huang-Lin, E.; Sánchez-León, E.; Amils, R.; Abrusci, C. Potential applications of an exopolysaccharide produced by Bacillus xiamenensis RT6 isolated from an acidic environment. Polymers 2022, 14, 3918. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides produced by lactic acid bacteria: From biosynthesis to health-promoting properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Dănăilă-Guidea, S.M.; Tamba-Berehoiu, R.M.; Popa, C.-N.; Toma, R.; Vișan, L.; Simion, V. Inulin: Unique among the polyglucides with significant functional properties and biotechnological perspectives. Rom. Biotechnol. Lett. 2020, 25, 1387–1395. [Google Scholar] [CrossRef]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Prete, R.; Alam, M.K.; Perpetuini, G.; Perla, C.; Pittia, P.; Corsetti, A. Lactic acid bacteria exopolysaccharides producers: A sustainable tool for functional foods. Foods 2021, 10, 1653. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, M.A.M.; Hassan, A.I.; Mahmoud, M.G.; Asker, M.S. Effect of polysaccharide from Bacillus subtilis sp. on cardiovascular diseases and atherogenic indices in diabetic rats. BMC Complement. Altern. Med. 2016, 16, 112. [Google Scholar] [CrossRef]
- Malick, A.; Khodaei, N.; Benkerroum, N.; Karboune, S. Production of exopolysaccharides by selected Bacillus strains: Optimization of media composition to maximize the yield and structural characterization. Int. J. Biol. Macromol. 2017, 102, 539–549. [Google Scholar] [CrossRef]
- Daneshazari, R.; Khorasgani, M.R.; Hosseini-Abari, A.; Kim, J.-H. Bacillus subtilis isolates from camel milk as probiotic candidates. Sci. Rep. 2023, 13, 3387. [Google Scholar] [CrossRef]
- Nayak, S.K. Multifaceted applications of probiotic Bacillus species in aquaculture with special reference to Bacillus subtilis. Rev. Aquac. 2020, 13, 862–906. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlishvili, E.; Chikindas, M.L. Recent advances in the physiology of spore formation for bacillus probiotic production. Probiotics Antimicrob. Proteins 2018, 11, 731–747. [Google Scholar] [CrossRef]
- Zhu, L.; Qin, S.; Zhai, S.; Gao, Y.; Li, L. Inulin with different degrees of polymerization modulates composition of intestinal microbiota in mice. FEMS Microbiol. Lett. 2017, 364, fnx075. [Google Scholar] [CrossRef]
- Karirat, T.; Saengha, W.; Deeseenthum, S.; Ma, N.L.; Sutthi, N.; Wangkahart, E.; Luang-In, V. Data on exopolysaccharides produced by Bacillus spp. from cassava pulp with antioxidant and antimicrobial properties. Data Brief 2023, 50, 109474. [Google Scholar] [CrossRef]
- Yones, A.-M.A.S. Effects of dietary inulin as prebiotic on growth performance, immuno-haematological indices and ectoparasitic infection of fingerlings Nile tilapia, Oreochromis niloticus. Egypt. J. Histol. 2019, 43, 88–103. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Ascencio, F.; Gracia-Lopez, V.; Macias, M.E.; Roa, M.C.; Esteban, M. Single or combined effects of Lactobacillus sakei and inulin on growth, non-specific immunity and IgM expression in leopard grouper (Mycteroperca rosacea). Fish Physiol. Biochem. 2014, 40, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Ghafarifarsani, H.; Rashidian, G.; Bagheri, T.; Hoseinifar, S.H.; Van Doan, H. Study on growth enhancement and the protective effects of dietary prebiotic inulin on immunity responses of rainbow trout (Oncorhynchus mykiss) fry infected with Aeromonas hydrophila. Ann. Anim. Sci. 2021, 21, 543–559. [Google Scholar] [CrossRef]
- Kumar, V.; Makkar, H.; Becker, K. Nutritional, physiological and haematological responses in rainbow trout (Oncorhynchus mykiss) juveniles fed detoxified Jatropha curcas kernel meal. Aquac. Nutr. 2010, 17, 451–467. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Mw (Da) | Mn (Da) | PDI | References |
|---|---|---|---|---|
| Lb. plantarum C7 | 3.37 × 104 | 0.85 × 104 | 3.96 | Ziadi et al. [31] |
| Lb. delbrueckii subsp. bulgaricus | 104 to 106 | n.r. | n.r | Jurášková et al. [50] |
| Lactobacillus sp. B3-1 | 8.31 × 105 | 3.38 × 105 | 2.46 | This study |
| B. amyloliquefaciens GSBa-1 | 5.40 × 104 | n.r. | 1.78 | Zhao et al. [19] |
| B. velezensis KY471306 | 1.29 × 105 | 1.14 × 105 | 1.13 | Moghannem et al. [21] |
| B. subtilis | 0.57 to 1.28 × 105 | n.r. | n.r | Marvasi et al. [45] |
| Bacillus sp. P1 | 9.81 × 105 | 6.04 × 105 | 1.62 | This study |
| Source | Sum of Squares | Mean Square | F-Value | p-Value * |
|---|---|---|---|---|
| Model | 6.268 × 105 | 69,645.11 | 20.23 | 0.0003 |
| A:sucrose | 33,303.93 | 33,303.93 | 9.67 | 0.0171 |
| B:yeast extract | 33,697.38 | 33,697.38 | 9.79 | 0.0166 |
| C:(NH4)2SO4 | 55,461.15 | 55,461.15 | 16.11 | 0.0051 |
| AB | 8689.97 | 8689.97 | 2.52 | 0.1561 |
| AC | 3863.24 | 3863.24 | 1.12 | 0.3246 |
| BC | 9259.25 | 9259.25 | 2.69 | 0.1450 |
| A2 | 59,541.62 | 59,541.62 | 17.30 | 0.0042 |
| B2 | 51,357.16 | 51,357.16 | 14.92 | 0.0062 |
| C2 | 3.325 × 105 | 3.325 × 105 | 96.59 | <0.0001 |
| Lack of Fit | 22,795.51 | 7598.50 | 23.36 | 0.0054 |
| Pure Error | 1301.35 | 325.34 | - | - |
| R2 | 0.9630 | - | - | |
| Adjusted R2 | 0.9154 | - | - | - |
| Duration (Week) | Experiment | Growth Performance Parameters | |||||
|---|---|---|---|---|---|---|---|
| Weight Gain | Specific Growth Rate (%) | Average Daily Gain | Feed Conversion Rate (g/g) | ||||
| av. per Tank (g) | av. per Fish (g) | av. per Tank (g/day) | av. per Fish (g/day) | ||||
| 2 | Control | 527.55 ± 13.35 a | 21.10 | 4.83 ± 0.08 a | 37.68 ± 0.95 a | 1.51 | 1.30 ± 0.06 a |
| Treatment | 529.40 ± 19.83 a | 21.18 | 4.98 ± 0.14 a | 37.81 ± 1.42 a | 1.51 | 1.36 ± 0.05 a | |
| 4 | Control | 990.28 ± 25.91 b | 39.61 | 3.69 ± 0.03 b | 35.37 ± 0.93 b | 1.41 | 0.66 ± 0.03 b |
| Treatment | 1047.00 ± 11.80 c | 41.88 | 3.92 ± 0.07 c | 37.40 ± 0.42 c | 1.50 | 0.71 ± 0.02 b | |
| 6 | Control | 1403.86 ± 34.17 d | 56.15 | 3.03 ± 0.03 d | 33.43 ± 0.81 d | 1.34 | 0.42 ± 0.04 d |
| Treatment | 1464.57 ± 29.22 d | 58.58 | 3.17 ± 0.09 d | 34.87 ± 0.70 d | 1.40 | 0.43 ± 0.00 d | |
| Parameters | Control | Treatment |
|---|---|---|
| Lysozyme activity (U/min) | 3.37 ± 0.13 | 6.88 ± 0.16 * |
| Respiratory burst activity a (OD630) | 0.10 | 0.44 |
| Total immunoglobulin (mg/dL) | 3 ± 0 | 3 ± 0 |
| Relative level of protection (%RLP) after 2 weeks challenge test | 0 | 50 |
| Analysis | Parameters | Control | Treatment |
|---|---|---|---|
| Blood chemistry | Blood urea nitrogen (mg/dL) | 3 ± 1 | 3 ± 0 |
| Glucose (mg/dL) | 98.33 ± 15.89 a | 118.67 ± 11.59 ab | |
| ALT (U/L) | 12.67 ± 7.77 | 15.33 ± 3.51 | |
| AST (U/L) | 68.33 ± 32.59 | 58.67 ± 30.29 | |
| Total bilirubin (mg/dL) | 0.23 ± 0.06 | 0.20 ± 0.00 | |
| Direct bilirubin (mg/dL) | 0.26 ± 0.05 | 0.23 ± 0.05 | |
| Total cholesterol (mg/dL) | 197.33 ± 5.03 | 190.67 ± 16.29 | |
| Total protein (g/dL) | 4.30 ± 0.26 | 4.43 ± 0.06 | |
| Albumin (mg/dL) | 1.47 ± 0.06 | 1.43 ± 0.06 | |
| Proximate chemical compositions (AOAC 2000 *) | Crude protein ** | 17.80 ± 0.04 | 19.02 ± 0.10 |
| Crude fat | 0.33 ± 0.01 | 0.83 ± 0.03 | |
| Moisture content | 71.91 ± 0.50 | 69.86 ± 0.20 | |
| Ash | 1.85 ± 0.01 | 2.30 ± 0.02 | |
| Carbohydrate | 8.11 ± 0.02 | 7.99 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunkaew, K.; Yenraksa, A.; Pewhom, A.; Kiriratnikom, S.; Sewaka, M.; Lertworapreecha, M.; Chanasit, W. Dietary Administration Effects of Microbial Exopolysaccharide from Bacillus subtilis P1 on Growth Performance and Immunity in Nile Tilapia (Oreochromis niloticus). Microbiol. Res. 2024, 15, 2210-2231. https://doi.org/10.3390/microbiolres15040148
Bunkaew K, Yenraksa A, Pewhom A, Kiriratnikom S, Sewaka M, Lertworapreecha M, Chanasit W. Dietary Administration Effects of Microbial Exopolysaccharide from Bacillus subtilis P1 on Growth Performance and Immunity in Nile Tilapia (Oreochromis niloticus). Microbiology Research. 2024; 15(4):2210-2231. https://doi.org/10.3390/microbiolres15040148
Chicago/Turabian StyleBunkaew, Kannika, Auraiwan Yenraksa, Akkanee Pewhom, Suphada Kiriratnikom, Mariya Sewaka, Monthon Lertworapreecha, and Wankuson Chanasit. 2024. "Dietary Administration Effects of Microbial Exopolysaccharide from Bacillus subtilis P1 on Growth Performance and Immunity in Nile Tilapia (Oreochromis niloticus)" Microbiology Research 15, no. 4: 2210-2231. https://doi.org/10.3390/microbiolres15040148
APA StyleBunkaew, K., Yenraksa, A., Pewhom, A., Kiriratnikom, S., Sewaka, M., Lertworapreecha, M., & Chanasit, W. (2024). Dietary Administration Effects of Microbial Exopolysaccharide from Bacillus subtilis P1 on Growth Performance and Immunity in Nile Tilapia (Oreochromis niloticus). Microbiology Research, 15(4), 2210-2231. https://doi.org/10.3390/microbiolres15040148