
Skeletal Muscle Insulin Resistance in a Novel Fetal Growth Restriction Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
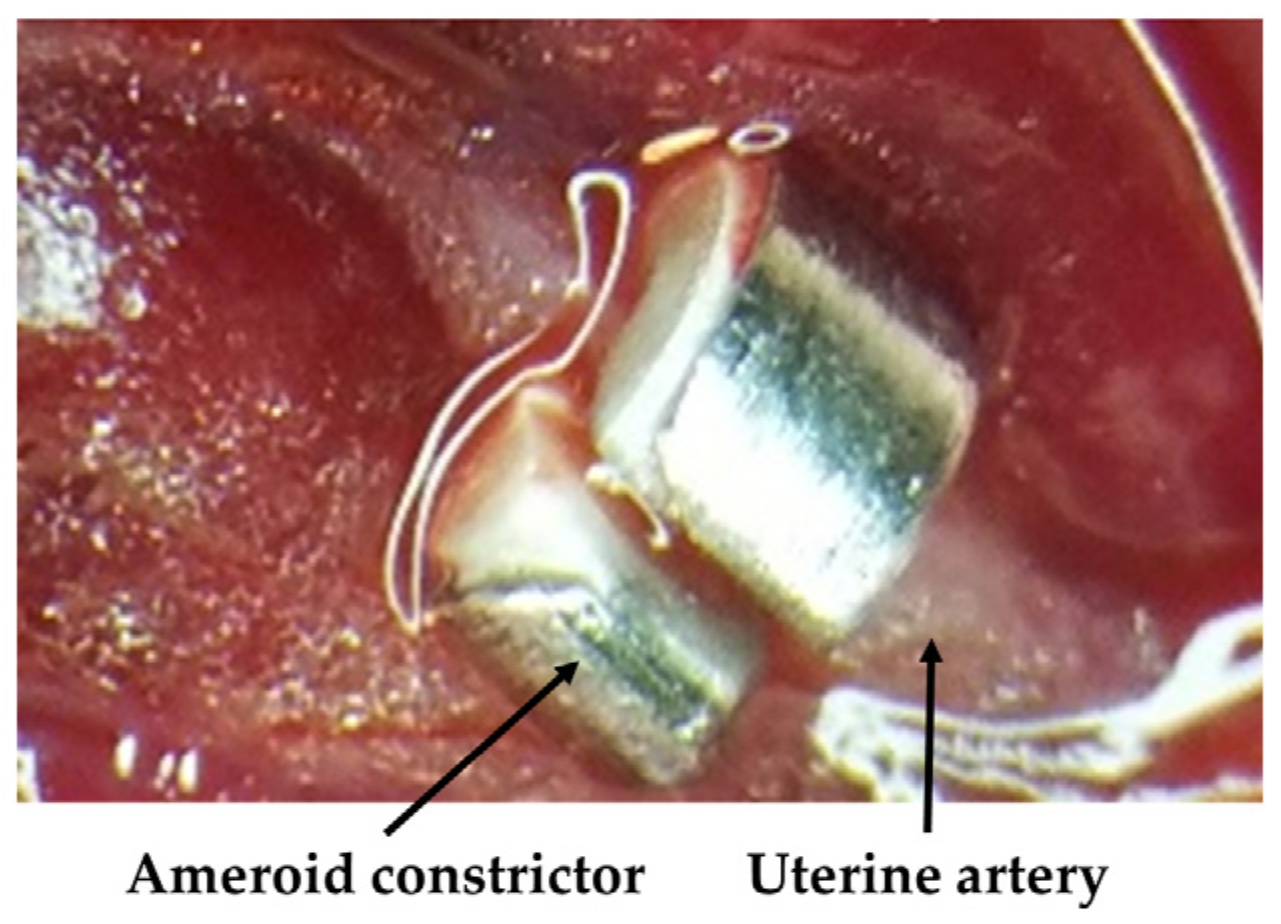
2.1. Animals and Experimental Design
2.2. Measurement of Glucose Metabolic Indices
2.3. Real-Time Quantitative Reverse Transcription–Polymerase Chain Reaction (RT-PCR)
2.4. Western Blot Assay
2.5. Immunohistochemistry for Adipophilin
2.6. Statistical Analysis
3. Results
3.1. Body Weight
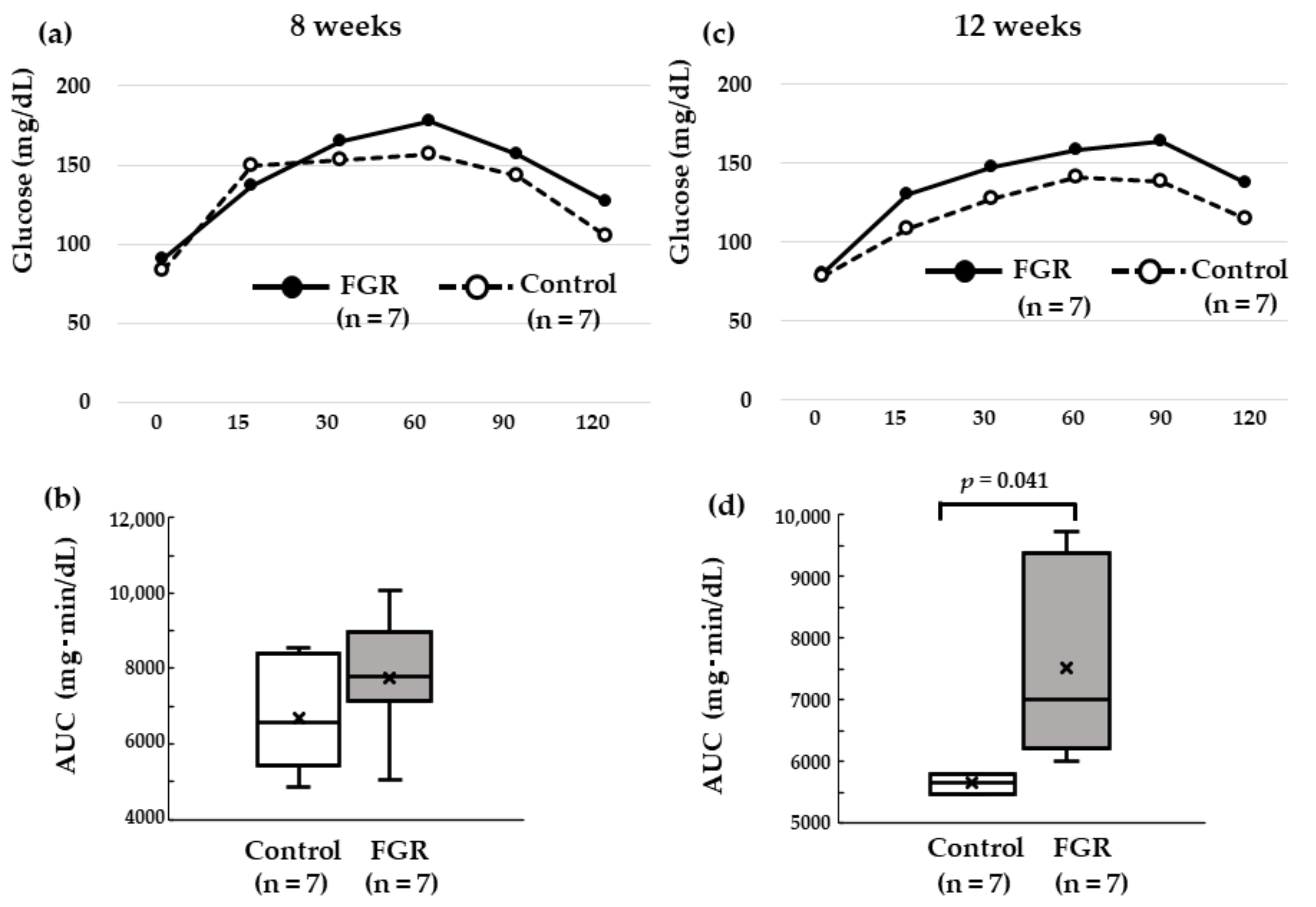
3.2. OGTT
3.3. RT-PCR of Insulin Signaling in Skeletal Muscles
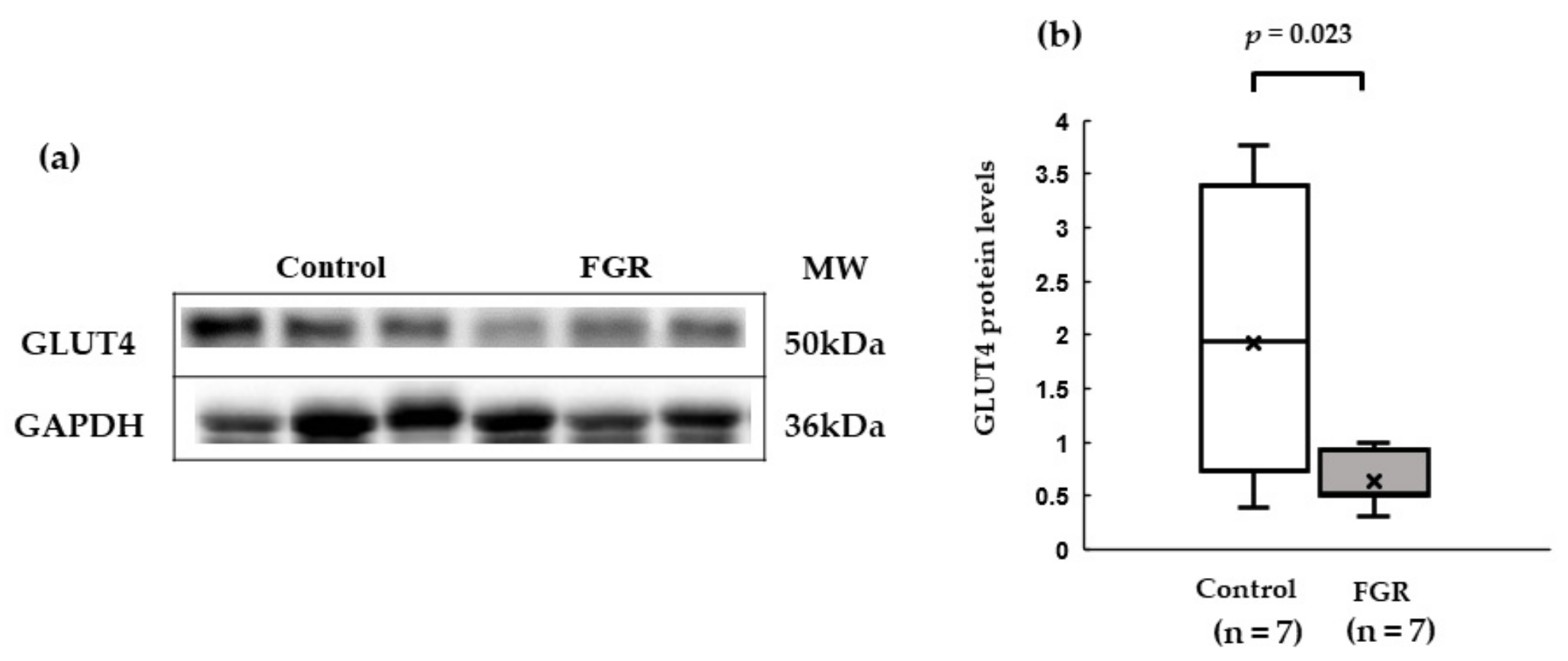
3.4. Western Blot Analysis of Insulin Signaling in Skeletal Muscles
3.5. Association between GLUT4 Expression and iAUC at 12 Weeks of Age
3.6. Intracellular Lipid DROPLET accumulation in the Liver and Skeletal Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J.P.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in england and wales. Lancet 1986, 327, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Szostak-Węgierek, D.; Szamotulska, K. Fetal development and risk of cardiovascular diseases and diabetes type 2 in adult life. Med. Wieku Rozw. 2011, 15, 203–215. [Google Scholar]
- Mierzynski, R.; Dluski, D.; Darmochwal-Kolarz, D.; Poniedziałek-Czajkowska, E.; Leszczynska-Gorzelak, B.; Kimber-Trojnar, Z.; Wankowicz, A.; Oleszczuk, J. Intra-uterine Growth Retardation as a Risk Factor of Postnatal Metabolic Disorders. Curr. Pharm. Biotechnol. 2016, 17, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, Y.; Ota, E.; Yoneoka, D.; Mori, R.; Takeda, S. Japanese secular trends in birthweight and the prevalence of low birthweight infants during the last three decades: A population-based study. Sci. Rep. 2016, 6, 31396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, E.A.; Longo, L.D.; Goyal, R. Antenatal maternal hypoxia: Criterion for fetal growth restriction in rodents. Front. Physiol. 2015, 6, 176. [Google Scholar] [CrossRef] [Green Version]
- Wixey, J.A.; Chand, K.K.; Colditz, P.B.; Bjorkman, S.T. Review: Neuroinflammation in intrauterine growth restriction. Placenta 2017, 54, 117–124. [Google Scholar] [CrossRef] [Green Version]
- DeFronzo, R.A.; Jacot, E.; Jequier, E.; Maeder, E.; Wahren, J.; Felber, J.P. The Effect of Insulin on the Disposal of Intravenous Glucose: Results from Indirect Calorimetry and Hepatic and Femoral Venous Catheterization. Diabetes 1981, 30, 1000–1007. [Google Scholar] [CrossRef]
- Zurlo, F.; Nemeth, P.M.; Choksi, R.M.; Sesodia, S.; Ravussin, E. Whole-body energy metabolism and skeletal muscle biochemical characteristics. Metabolism 1994, 43, 481–486. [Google Scholar] [CrossRef]
- Bridges, D.; Saltiel, A.R. Phosphoinositides: Key modulators of energy metabolism. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Welsh, G.; Hers, I.; Berwick, D.; Dell, G.; Wherlock, M.; Birkin, R.; Leney, S.; Tavaré, J. Role of protein kinase B in insulin-regulated glucose uptake. Biochem. Soc. Trans. 2005, 33, 346–349. [Google Scholar] [CrossRef]
- Someya, Y.; Tamura, Y.; Suzuki, R.; Kaga, H.; Kadowaki, S.; Sugimoto, D.; Kakehi, S.; Funayama, T.; Furukawa, Y.; Takeno, K.; et al. Characteristics of Glucose Metabolism in Underweight Japanese Women. J. Endocr. Soc. 2018, 2, 279–289. [Google Scholar] [CrossRef]
- Năstase, L.; Cretoiu, D.; Stoicescu, S.M. Skeletal Muscle Damage in Intrauterine Growth Restriction. Muscle Atrophy 2018, 93–106. [Google Scholar] [CrossRef]
- Harder, T.; Rodekamp, E.; Schellong, K.; Dudenhausen, J.W.; Plagemann, A. Birth Weight and Subsequent Risk of Type 2 Diabetes: A Meta-Analysis. Am. J. Epidemiol. 2007, 165, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Oya, J.; Nakagami, T.; Kurita, M.; Yamamoto, Y.; Hasegawa, Y.; Tanaka, Y.; Endo, Y.; Uchigata, Y. Association of birthweight with diabetes and insulin sensitivity or secretion in the Japanese general population. J. Diabetes Investig. 2015, 6, 430–435. [Google Scholar] [CrossRef]
- Ruff, C.A.; Faulkner, S.D.; Rumajogee, P.; Beldick, S.; Foltz, W.; Corrigan, J.; Basilious, A.; Jiang, S.; Thiyagalingam, S.; Yager, J.Y.; et al. The extent of intrauterine growth restriction determines the severity of cerebral injury and neurobehavioural deficits in rodents. PLoS ONE 2017, 12, e0184653. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, N.; Shoji, H.; Suganuma, H.; Ohkawa, N.; Kantake, M.; Murano, Y.; Sakuraya, K.; Shimizu, T. Effect of insulin-like growth factor-I during the early postnatal period in intrauterine growth-restricted rats. Pediatr. Int. 2016, 58, 353–358. [Google Scholar] [CrossRef]
- Simmons, R.A.; Templeton, L.J.; Gertz, S.J. Intrauterine Growth Retardation Leads to the Development of Type 2 Diabetes in the Rat. Diabetes 2001, 50, 2279–2286. [Google Scholar] [CrossRef] [Green Version]
- Selak, M.A.; Storey, B.T.; Peterside, I.; Simmons, R.A. Impaired oxidative phosphorylation in skeletal muscle of intrauterine growth-retarded rats. Am. J. Physiol. Metab. 2003, 285, E130–E137. [Google Scholar] [CrossRef] [Green Version]
- Kitase, Y.; Sato, Y.; Arai, S.; Onoda, A.; Ueda, K.; Go, S.; Mimatsu, H.; Jabary, M.; Suzuki, T.; Ito, M.; et al. Establishment of a Novel Fetal Growth Restriction Model and Development of a Stem-Cell Therapy Using Umbilical Cord-Derived Mesenchymal Stromal Cells. Front. Cell. Neurosci. 2020, 14, 212. [Google Scholar] [CrossRef]
- Salmaso, N.; Jablonska, B.; Scafidi, J.; Vaccarino, F.M.; Gallo, V. Neurobiology of premature brain injury. Nat. Neurosci. 2014, 17, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Gatford, K.; Simmons, R.; De Blasio, M.; Robinson, J.; Owens, J. Review: Placental Programming of Postnatal Diabetes and Impaired Insulin Action after IUGR. Placenta 2010, 31, S60–S65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatulian, S.A. Structural Dynamics of Insulin Receptor and Transmembrane Signaling. Biochemistry 2015, 54, 5523–5532. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, R.A.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2017, 19, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Lillioja, S.; Young, A.A.; Culter, C.L.; Ivy, J.L.; Abbott, W.G.; Zawadzki, J.K.; Yki-Järvinen, H.; Christin, L.; Secomb, T.W.; Bogardus, C. Skeletal muscle capillary density and fiber type are possible determinants of in vivo insulin resistance in man. J. Clin. Investig. 1987, 80, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Nagatomo, F.; Mori, A.; Ohira, Y.; Ishihara, A. Cell Size and Oxidative Enzyme Activity of Rat Biceps Brachii and Triceps Brachii Muscles. J. Physiol. Sci. 2007, 57, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, A.; Itoh, K.; Itoh, M.; Hirofuji, C. Effect of Hypobaric Hypoxia on Rat Soleus Muscle Fibers and Their Innervating Motoneurons: A Review. Jpn. J. Physiol. 2000, 50, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, E.J.; Bourey, R.E.; Rodnick, K.J.; Koranyi, L.; Permutt, M.A.; Holloszy, J.O. Glucose transporter protein content and glucose transport capacity in rat skeletal muscles. Am. J. Physiol. Metab. 1990, 259, E593–E598. [Google Scholar] [CrossRef]
- Daugaard, J.R.; Nielsen, J.N.; Kristiansen, S.; Andersen, J.L.; Hargreaves, M.; Richter, E.A. Fiber type-specific expression of GLUT4 in human skeletal muscle: Influence of exercise training. Diabetes 2000, 49, 1092–1095. [Google Scholar] [CrossRef] [Green Version]
- Gaster, M.; Poulsen, P.; Handberg, A.; Schrøder, H.D.; Beck-Nielsen, H. Direct evidence of fiber type-dependent GLUT-4 expression in human skeletal muscle. Am. J. Physiol. Metab. 2000, 278, E910–E916. [Google Scholar] [CrossRef] [Green Version]
- Esteves, J.V.; Enguita, F.J.; Machado, U.F. MicroRNAs-Mediated Regulation of Skeletal Muscle GLUT4 Expression and Translocation in Insulin Resistance. J. Diabetes Res. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, J.; Wei, H.; Zhang, H.; Guan, Y.; Wang, X.; Tong, X. Reduction of the PI3K/Akt related signaling activities in skeletal muscle tissues involves insulin resistance in intrauterine growth restriction rats with catch-up growth. PLoS ONE 2019, 14, e0216665. [Google Scholar] [CrossRef] [Green Version]
- Thamotharan, M.; Shin, B.-C.; Suddirikku, D.T.; Thamotharan, S.; Garg, M.; Devaskar, S.U. GLUT4 expression and subcellular localization in the intrauterine growth-restricted adult rat female offspring. Am. J. Physiol. Metab. 2005, 288, E935–E947. [Google Scholar] [CrossRef]
- Wang, J.; Cao, M.; Yang, M.; Lin, Y.; Che, L.; Fang, Z.; Xu, S.; Feng, B.; Li, J.; Wu, D. Intra-uterine undernutrition amplifies age-associated glucose intolerance in pigs via altered DNA methylation at muscle GLUT4 promoter. Br. J. Nutr. 2016, 116, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.K.; Wang, D.; Goldstrohm, D.A.; Brown, L.D.; Rozance, P.J.; Limesand, S.W.; Wesolowski, S.R. Tissue-specific responses that constrain glucose oxidation and increase lactate production with the severity of hypoxemia in fetal sheep. Am. J. Physiol. Metab. 2022, 322, E181–E196. [Google Scholar] [CrossRef]
- Ozanne, S.E.; Jensen, C.B.; Tingey, K.J.; Storgaard, H.; Madsbad, S.; Vaag, A.A. Low birthweight is associated with specific changes in muscle insulin-signalling protein expression. Diabetologia 2005, 48, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Camm, E.J.; Martin-Gronert, M.S.; Wright, N.L.; Hansell, J.A.; Ozanne, S.E.; Giussani, D.A. Prenatal hypoxia independent of undernutrition promotes molecular markers of insulin resistance in adult offspring. FASEB J. 2010, 25, 420–427. [Google Scholar] [CrossRef]
- Martins, V.F.; Tahvilian, S.; Kang, J.H.; Svensson, K.; Hetrick, B.; Chick, W.S.; Schenk, S.; McCurdy, C.E. Calorie Restriction-Induced Increase in Skeletal Muscle Insulin Sensitivity Is Not Prevented by Overexpression of the p55α Subunit of Phosphoinositide 3-Kinase. Front. Physiol. 2018, 9, 789. [Google Scholar] [CrossRef]
- Garofalo, R.S.; Orena, S.J.; Rafidi, K.; Torchia, A.J.; Stock, J.L.; Hildebrandt, A.L.; Coskran, T.; Black, S.C.; Brees, D.J.; Wicks, J.R.; et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKBβ. J. Clin. Investig. 2003, 112, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Thorn, S.R.; Regnault, T.; Brown, L.D.; Rozance, P.J.; Keng, J.; Roper, M.; Wilkening, R.B.; Hay, W.W.; Friedman, J.E. Intrauterine Growth Restriction Increases Fetal Hepatic Gluconeogenic Capacity and Reduces Messenger Ribonucleic Acid Translation Initiation and Nutrient Sensing in Fetal Liver and Skeletal Muscle. Endocrinology 2009, 150, 3021–3030. [Google Scholar] [CrossRef]
- Esteves, J.V.; Yonamine, C.Y.; Pinto-Junior, D.C.; Gerlinger-Romero, F.; Enguita, F.J.; Machado, U.F. Diabetes Modulates MicroRNAs 29b-3p, 29c-3p, 199a-5p and 532-3p Expression in Muscle: Possible Role in GLUT4 and HK2 Repression. Front. Endocrinol. 2018, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Giorgino, F.; de Robertis, O.; Laviola, L.; Montrone, C.; Perrini, S.; McCowen, K.C.; Smith, R.J. The sentrin-conjugating enzyme mUbc9 interacts with GLUT4 and GLUT1 glucose transporters and regulates transporter levels in skeletal muscle cells. Proc. Natl. Acad. Sci. 2000, 97, 1125–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleasby, M.E.; Jamieson, P.M.; Atherton, P.J. Insulin resistance and sarcopenia: Mechanistic links between common co-morbidities. J. Endocrinol. 2016, 229, R67–R81. [Google Scholar] [CrossRef] [PubMed]
- Conte, M.; Vasuri, F.; Trisolino, G.; Bellavista, E.; Santoro, A.; Degiovanni, A.; Martucci, E.; D’Errico-Grigioni, A.; Caporossi, D.; Capri, M.; et al. Increased Plin2 Expression in Human Skeletal Muscle Is Associated with Sarcopenia and Muscle Weakness. PLoS ONE 2013, 8, e73709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meex, R.C.; Blaak, E.E.; Van Loon, L.J. Lipotoxicity plays a key role in the development of both insulin resistance and muscle atrophy in patients with type 2 diabetes. Obes. Rev. 2019, 20, 1205–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, E.S.; Campbell, M.S.; Lang, B.Y.; Bierer, R.; Wang, Y.; Bagley, H.N.; Joss-Moore, L.A. Intrauterine Growth Restriction Increases TNFαand Activates the Unfolded Protein Response in Male Rat Pups. J. Obes. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L.J. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Jellyman, J.K.; Han, G.; Beall, M.; Lane, R.H.; Ross, M.G. Maternal obesity and high-fat diet program offspring metabolic syndrome. Am. J. Obstet. Gynecol. 2014, 211, 237.e1–237.e13. [Google Scholar] [CrossRef] [Green Version]
- Yajnik, C.S.; Fall, C.H.D.; Coyaji, K.J.; Hirve, S.S.; Rao, S.; Barker, D.J.P.; Joglekar, C.; Kellingray, S. Neonatal anthropometry: The thin–fat Indian baby. The Pune Maternal Nutrition Study. Int. J. Obes. 2003, 27, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Lane, R.H.; Kelley, D.E.; Ritov, V.H.; Tsirka, A.E.; Gruetzmacher, E.M. Altered Expression and Function of Mitochondrial β-Oxidation Enzymes in Juvenile Intrauterine-Growth-Retarded Rat Skeletal Muscle. Pediatr. Res. 2001, 50, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, D.; Yao, Y.; Yu, B.; Mao, X.; He, J.; Huang, Z.; Zheng, P. Intrauterine Growth Retardation Increases the Susceptibility of Pigs to High-Fat Diet-Induced Mitochondrial Dysfunction in Skeletal Muscle. PLoS ONE 2012, 7, e34835. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 7) | FGR (n = 7) | |
|---|---|---|
| Birth | 6.5 ± 0.2 g | 5.8 ± 0.6 g * |
| 4 weeks | 96.0 ± 13.3 g | 84.4 ± 12.6 g |
| 8 weeks | 315.9 ± 9.8 g | 346.3 ± 38.9 g |
| 12 weeks | 436.1 ± 16.1 g | 430.6 ± 51.2 g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokita, K.; Shoji, H.; Arai, Y.; Awata, K.; Santosa, I.; Murano, Y.; Shimizu, T. Skeletal Muscle Insulin Resistance in a Novel Fetal Growth Restriction Model. Pediatr. Rep. 2023, 15, 45-54. https://doi.org/10.3390/pediatric15010006
Tokita K, Shoji H, Arai Y, Awata K, Santosa I, Murano Y, Shimizu T. Skeletal Muscle Insulin Resistance in a Novel Fetal Growth Restriction Model. Pediatric Reports. 2023; 15(1):45-54. https://doi.org/10.3390/pediatric15010006
Chicago/Turabian StyleTokita, Kazuhide, Hiromichi Shoji, Yoshiteru Arai, Kentaro Awata, Irena Santosa, Yayoi Murano, and Toshiaki Shimizu. 2023. "Skeletal Muscle Insulin Resistance in a Novel Fetal Growth Restriction Model" Pediatric Reports 15, no. 1: 45-54. https://doi.org/10.3390/pediatric15010006
APA StyleTokita, K., Shoji, H., Arai, Y., Awata, K., Santosa, I., Murano, Y., & Shimizu, T. (2023). Skeletal Muscle Insulin Resistance in a Novel Fetal Growth Restriction Model. Pediatric Reports, 15(1), 45-54. https://doi.org/10.3390/pediatric15010006