
Menthalactone from Mentha piperita L., a Monocot-Selective Bioherbicide

 ,
,  , and
, and 
Abstract
1. Introduction
2. Experimental Section
2.1. General Experimental Procedures
2.2. Chemicals
2.2.1. Menthalactone (1)
2.2.2. δ-Dodecalactone (2)
2.2.3. γ-Nonalactone (3)
2.2.4. δ-Tetradecalactone (4)
2.2.5. δ-Undecalactone (5)
2.2.6. γ-Octalactone (6)
2.2.7. γ-Valerolactone (7)
2.2.8. δ-Hexalactone (8)
2.2.9. γ-Undecalactone (9)
2.2.10. D-(-)-Pantolactone (10)
2.2.11. γ-Hexalactone (11)
2.3. Phytotoxicity Bioassays
2.3.1. The Pre-Emergence Selection Test of Extracts and Pure Compounds
2.3.2. A Dose Response Bioassay of Lemna paucicostata
2.3.3. A Dose Response Bioassay of A. stolonifera
2.3.4. The Pre-Emergence Test with Selected Weed Species
2.3.5. The Pre-Emergence and Post-Emergence Bioassay with Selected Weed Species in Soil
2.4. Postemergence Application against Sorghum halepense L. Pers., Lolium multiflorum L., and E. crus-galli
2.4.1. Seeds and Growth of Plants
2.4.2. Postemergence Activity Assay, Calculations, and Statistics
3. Results
3.1. Bioassay-Guided Identification of Phytotoxic Components—Pre-Emergence Bioassay
3.2. Phytotoxicity of Selected Lactones
3.3. Dose–Response Analysis of Menthalactone against A. stolonifera and L. pausicostata
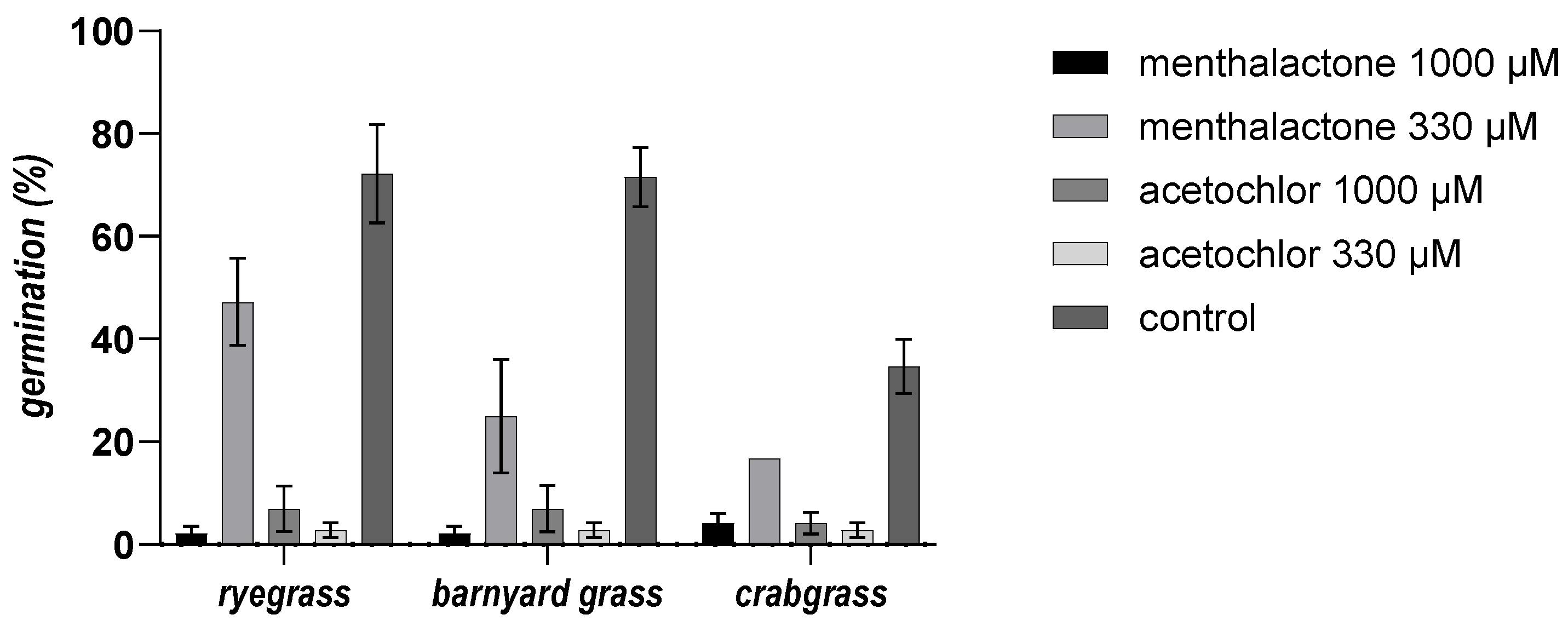
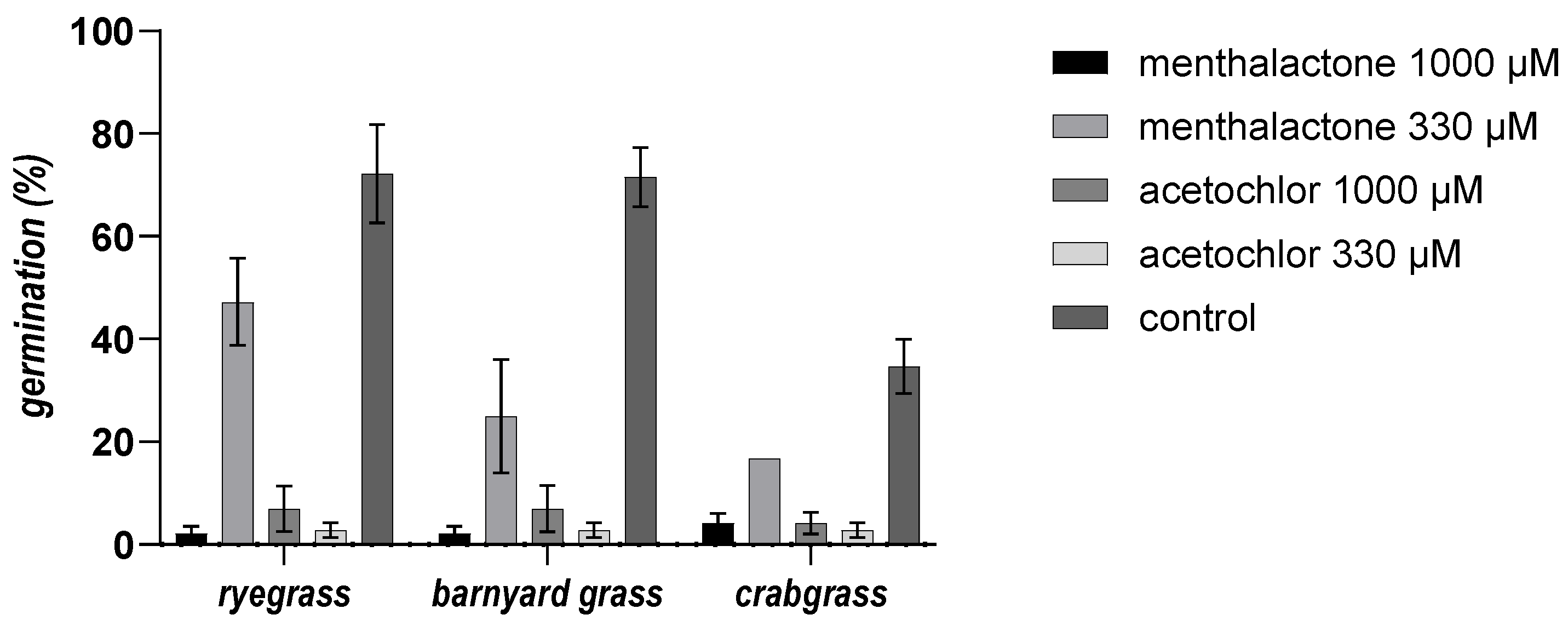
3.4. Phytotoxicity Bioassay In Vitro against Selected Weeds
3.5. Pre-Emergence Bioassay in Soil
3.6. Post-Emergence Application against Sorghum halepense, Lolium multiflorum, and E. crus-galli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stefanski, F.S.; Camargo, A.F.; Scapini, T.; Bonatto, C.; Venturin, B.; Weirich, S.N.; Ulkovski, C.; Carezia, C.; Ulrich, A.; Michelon, W.; et al. Potential use of biological herbicides in a circular economy context: A sustainable approach. Front. Sustain. Food Syst. 2020, 4, 521102. [Google Scholar] [CrossRef]
- Macbryde, B. White Paper: Perspective on Creeping Bentgrass, Agrostis stolonifera L.; USDA/APHIS/BRS: Riverdale, MD, USA, 2006. [Google Scholar]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef]
- Shigeto, A.; Wada, A.; Kumazawa, K. Identification of the novel odor active compounds “p-menthane lactones” responsible for the characteristic aroma of fresh peppermint leaf. Biosci. Biotechnol. Biochem. 2020, 84, 421–427. [Google Scholar] [CrossRef]
- Tamura, H.; Appel, M.; Richling, E.; Schreier, P. Authenticity assessment of γ-and δ-decalactone from Prunus fruits by gas chromatography combustion/pyrolysis isotope ratio mass spectrometry (GC-C/P-IRMS). J. Agric. Food Chem. 2005, 53, 5397–5401. [Google Scholar] [CrossRef] [PubMed]
- Sartori, S.K.; Diaz, M.A.N.; Diaz-Munoz, G. Lactones: Classification, synthesis, biological activities, and industrial applications. Tetrahedron 2021, 84, 132001. [Google Scholar] [CrossRef]
- Dhyani, P.; Sati, P.; Sharma, E.; Attri, D.C.; Bahukhandi, A.; Tynybekov, B.; Szopa, A.; Sharifi-Rad, J.; Calina, D.; Suleria, H.A.R.; et al. Sesquiterpenoid lactones as potential anti-cancer agents: An update on molecular mechanisms and recent studies. Cancer Cell Int. 2022, 22, 305. [Google Scholar] [CrossRef] [PubMed]
- Wedge, D.E.; Galindo, J.C.G.; Macıas, F.A. Fungicidal activity of natural and synthetic sesquiterpene lactone analogs. Phytochemistry 2000, 53, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Bridgford, J.L.; Xie, S.C.; Cobbold, S.A.; Pasaje, C.F.A.; Herrmann, S.; Yang, T.; Gillett, D.L.; Dick, L.R.; Ralph, S.A.; Dogovski, C.; et al. Artemisinin kills malaria parasites by damaging proteins and inhibiting the proteasome. Nat. Commun. 2018, 9, 3801. [Google Scholar] [CrossRef] [PubMed]
- Scavo, A.; Mauromicale, G. Crop allelopathy for sustainable weed management in agroecosystems: Knowing the present with a view to the future. Agronomy 2021, 11, 2104. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Flath, R.A.; Guentert, M.; Jennings, W. Nectarine volatiles: Vacuum steam distillation versus headspace sampling. J. Agric. Food Chem. 1988, 36, 553–560. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Flath, R.A.; Mon, T.R.; Teranishi, R.; Guentert, M. Volatile constituents of apricot (Prunus armeniaca). J. Agric. Food Chem. 1990, 38, 471–477. [Google Scholar] [CrossRef]
- Horvat, R.J.; Chapman, G.W., Jr.; Robertson, J.A.; Meredith, F.I.; Scorza, R.; Callahan, A.M.; Morgens, P. Comparison of the volatile compounds from several commercial peach cultivars. J. Agric. Food Chem. 1990, 38, 234–237. [Google Scholar] [CrossRef]
- Narain, N.; Hsieh, T.C.Y.; Johnson, C.E. Dynamic headspace concentration and gas chromatography of volatile flavor components in peach. J. Food Sci. 1990, 55, 1303–1307. [Google Scholar] [CrossRef]
- Do, J.Y. Isolation, Identification, and Comparison of the Volatiles of Peach (Prunus persica L., Cultivar, Gleason Early Elberta) Fruit as Related to Harvest Maturity and Artificial Ripening. In All Graduate Theses and Dissertations; Utah State University: Logan, UT, USA, 1968; p. 5104. [Google Scholar]
- Jennings, W.G.; Sevenants, M.R. Volatile Esters of Bartlett Pear. III. J. Food Sci. 1964, 29, 158–163. [Google Scholar] [CrossRef]
- Michel, A.; Johnson, R.D.; Duke, S.O.; Scheffler, B.E. Dose-response relationships between herbicides with different modes of action and growth of Lemna paucicostata: An improved ecotoxicological method. Environ. Toxicol. Chem. Int. J. 2004, 23, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Romagni, J.G.; Duke, S.O. Investigating the mode of action of natural phytotoxins. J. Chem. Ecol. 2000, 26, 2079–2094. [Google Scholar] [CrossRef]
- Krähmer, H.; Walter, H.; Jeschke, P.; Haaf, K.; Baur, P.; Evans, R. What makes a molecule a pre-or a post-herbicide–how valuable are physicochemical parameters for their design? Pest Manag. Sci. 2021, 77, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Plačková, L.; Kulkarni, M.G.; Doležal, K.; Van Staden, J. Role of smoke stimulatory and inhibitory biomolecules in phytochrome-regulated seed germination of Lactuca sativa. Plant Physiol. 2019, 181, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Kim, E.; Choi, S.; Kim, D.; Hong, W.; Choi, J.; Choi, H.; Kim, J.; Sable, G.A.; Markkandan, K.; et al. A Raf-like kinase is required for smoke-induced seed dormancy in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2020636118. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, Y.; Moustakas, H.; Api, A.M.; Smith, B.; Williams, G.; Greim, H.; Eisenbrand, G.; Dekant, W. Assessment of the genotoxic potential of mintlactone. Food Chem. Toxicol. 2022, 159, 112659. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| # | Ranking | |||
|---|---|---|---|---|
| Name | Structure | A. stolonifera | L. sativa | |
| 1 | Menthalactone | 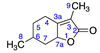 | 5 | 2 |
| 2 | δ-Dodecalactone | 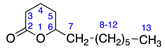 | 2 | 0 |
| 3 | γ-Nonalactone | 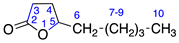 | 0 | 0 |
| 4 | δ-Tetradecalactone | 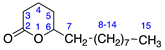 | 0 | 0 |
| 5 | δ-Undecalactone |  | 3 | 1 |
| 6 | γ-Octalactone |  | 3 | 0 |
| 7 | γ-Valerolactone | 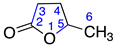 | 1 | 0 |
| 8 | δ-Hexalactone | 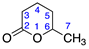 | 2 | 0 |
| 9 | γ-Undecalactone | 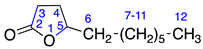 | 2 | 0 |
| 10 | D-(-)-Pantolactone |  | 1 | 0 |
| 11 | γ-Hexalactone |  | 1 | 0 |
| Name | Menthalactone (µM) | Acifluorfen (µM) |
|---|---|---|
| A.stolonifera | 4.9 ± 1.2 | 4.0 ± 1 |
| L.pausicostata | 293.4 ± 70.6 | 0.2 ±0.04 |
| Johnsongrass (S. halepense) * | Rye grass (L. multiflorum) * | Barnyard Grass (E. crus-galli) * | |
|---|---|---|---|
| water | 89.2 ± 11.28 a | 90.1 ± 23.83 a | 53.4 ± 12.03 a |
| 2% glyphosate | −94.9 ± 2.15 b | −51.7 ± 0.99 b | −93.7 ± 2.70 b |
| 1% menthalactone | 78.8 ± 8.19 a | 105.5 ± 4.35 a | 38.8 ± 2.47 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltani, A.; Ospanov, M.; Ibrahim, Z.M.A.; Bajsa-Hirschel, J.; Cantrell, C.L.; Cizdziel, J.V.; Khan, I.A.; Ibrahim, M.A. Menthalactone from Mentha piperita L., a Monocot-Selective Bioherbicide. Int. J. Plant Biol. 2024, 15, 293-303. https://doi.org/10.3390/ijpb15020025
Soltani A, Ospanov M, Ibrahim ZMA, Bajsa-Hirschel J, Cantrell CL, Cizdziel JV, Khan IA, Ibrahim MA. Menthalactone from Mentha piperita L., a Monocot-Selective Bioherbicide. International Journal of Plant Biology. 2024; 15(2):293-303. https://doi.org/10.3390/ijpb15020025
Chicago/Turabian StyleSoltani, Adam, Meirambek Ospanov, Zeyad M. A. Ibrahim, Joanna Bajsa-Hirschel, Charles L. Cantrell, James V. Cizdziel, Ikhlas A. Khan, and Mohamed A. Ibrahim. 2024. "Menthalactone from Mentha piperita L., a Monocot-Selective Bioherbicide" International Journal of Plant Biology 15, no. 2: 293-303. https://doi.org/10.3390/ijpb15020025
APA StyleSoltani, A., Ospanov, M., Ibrahim, Z. M. A., Bajsa-Hirschel, J., Cantrell, C. L., Cizdziel, J. V., Khan, I. A., & Ibrahim, M. A. (2024). Menthalactone from Mentha piperita L., a Monocot-Selective Bioherbicide. International Journal of Plant Biology, 15(2), 293-303. https://doi.org/10.3390/ijpb15020025