
Can BPA Analogs Affect Cellular and Biochemical Responses in the Microalga Phaeodactylum tricornutum Bohlin?
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalga Exposure
2.2. Growth Curve and Growth Rate
2.3. Biochemical Assays
2.4. Statistical Analysis
3. Results
4. Discussion
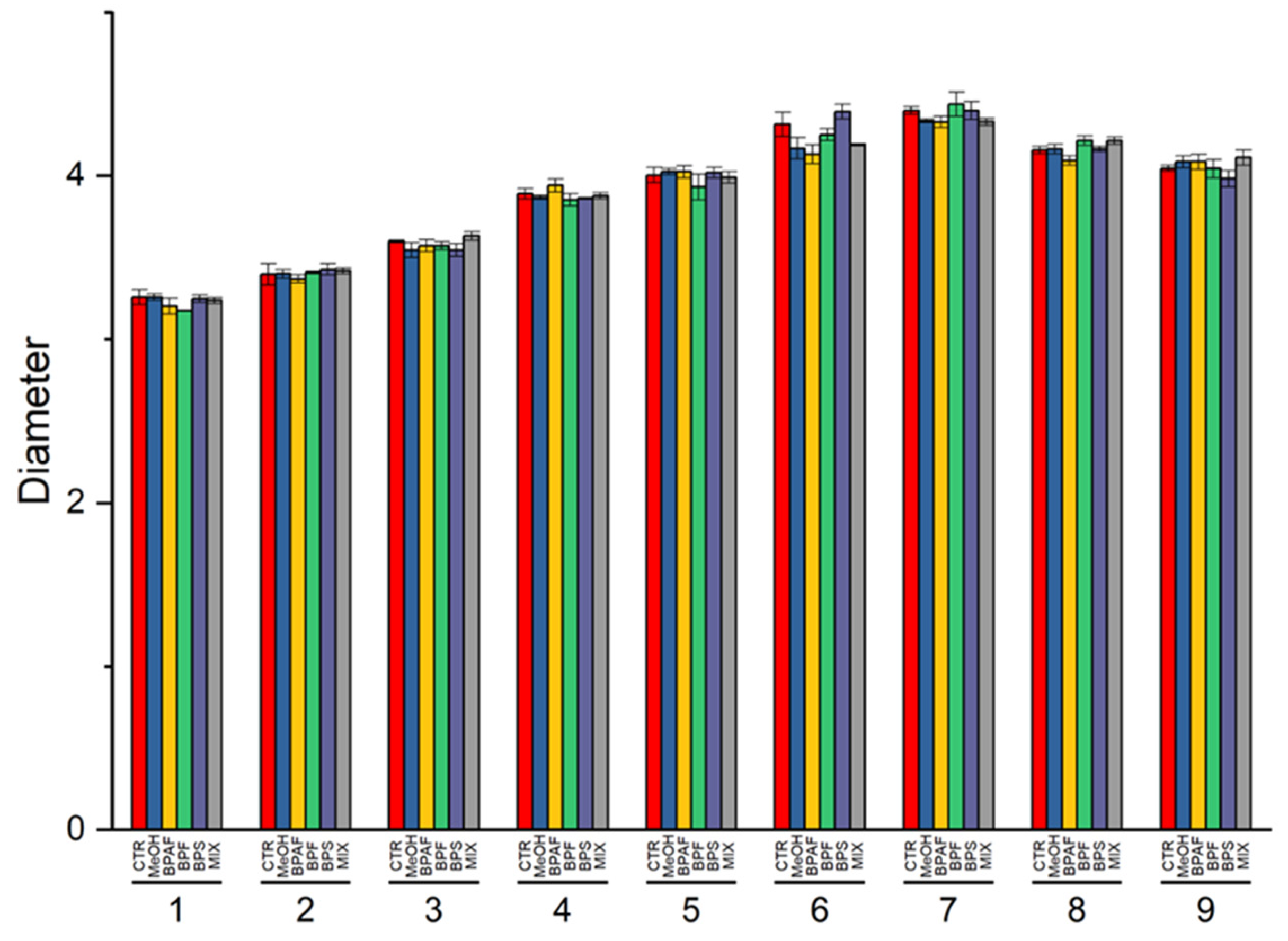
4.1. Growth and Cell Size
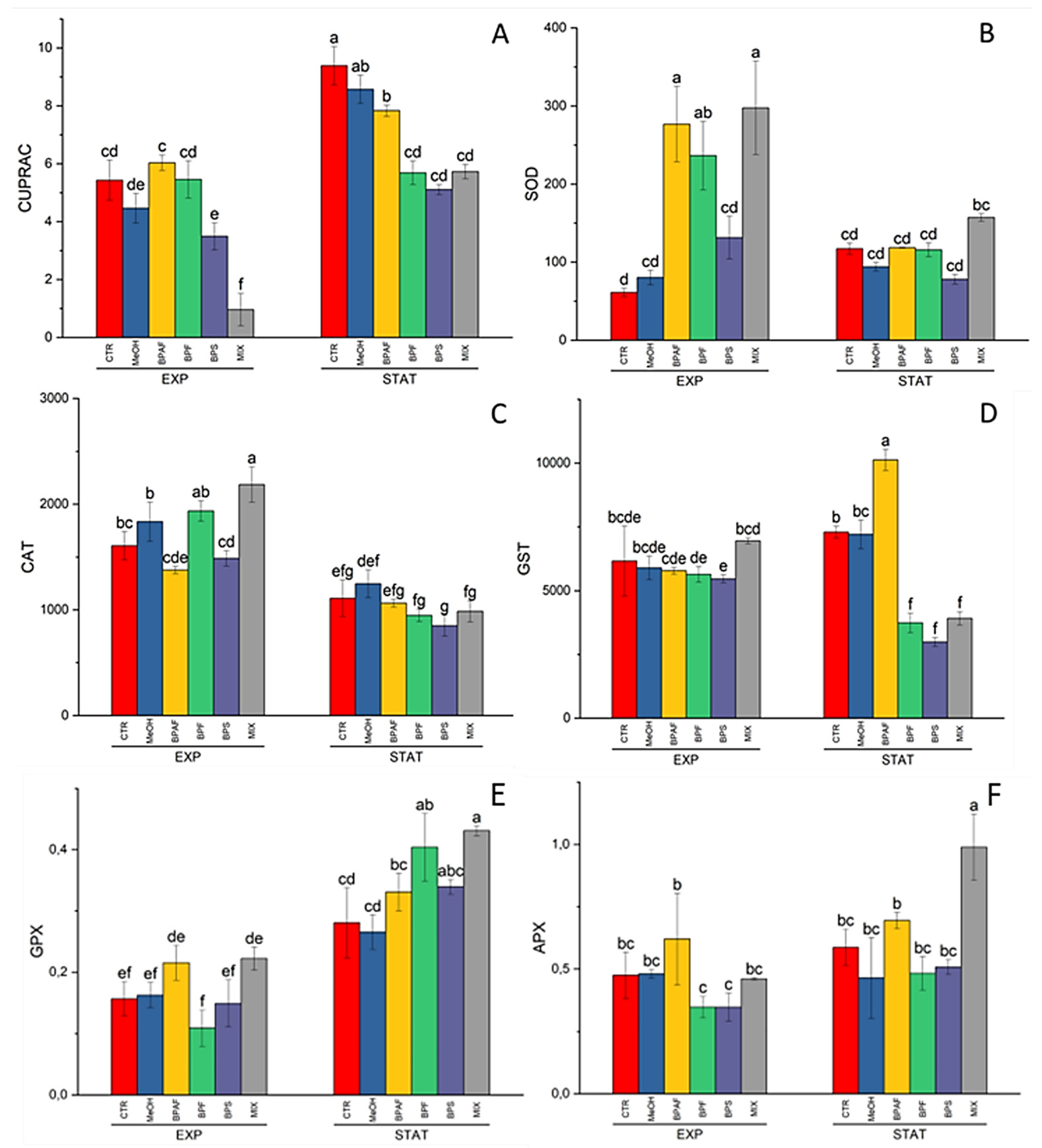
4.2. Biomarker Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Matuszczak, E.; Komarowska, M.D.; Debek, W.; Hermanowicz, A. The Impact of Bisphenol A on Fertility, Reproductive System, and Development: A Review of the Literature. Int. J. Endocrinol. 2019, 2019, 4068717. [Google Scholar] [CrossRef] [PubMed]
- Gassman, N.R. Induction of oxidative stress by bisphenol A and its pleiotropic effects. Environ. Mol. Mutagen. 2017, 58, 60–71. [Google Scholar] [CrossRef] [PubMed]
- EU. Commission regulation 2016/2235 of 12 December 2016 amending annex XVII to regulation (EC) No 1907/2006 of the European parliament and of the council concerning the registration, evaluation, authorisation and restriction of chemicals (REACH) as regards bisphenol A. Off. J. Eur. Union 2016, L337, 3–5. [Google Scholar]
- Chen, D.; Kannan, K.; Tan, H.; Zheng, Z.; Feng, Y.L.; Wu, Y.; Widelka, M. Bisphenol analogues other than BPA: Environmental occurrence, human exposure, and toxicity—A review. Environ. Sci. Technol. 2016, 50, 5438–5453. [Google Scholar] [CrossRef] [PubMed]
- Usman, A.; Ikhlas, S.; Ahmad, M. Occurrence, toxicity and endocrine disrupting potential of Bisphenol-B and Bisphenol-F: A mini-review. Toxicol. Lett. 2019, 312, 222–227. [Google Scholar] [CrossRef]
- EFSA; FitzGerald, R.; Van Loveren, H.; Civitella, C.; Castoldi, A.F.; Giovanni Bernasconi, G. Assessment of New Information on Bisphenol S (BPS) Submitted in Response to the Decision under REACH Regulation (EC) No 1907/2006; EFSA Supporting Publication; European Food Safety Authority (EFSA): Parma, Italy, 2020; p. 39. [Google Scholar]
- ECHA. Inclusion of BPA as a Substance of Very High Concern (Reason for Inclusion: Endocrine Disrupting Properties—Article 57f Environment) in the Candidate List for Eventual Inclusion in Annex XIV; Decision of the European Chemicals Agency; European Chemicals Agency: Helsinki, Finland, 2018. [Google Scholar]
- Hu, Y.; Zhu, Q.; Yan, X.; Liao, C.; Jiang, G. Occurrence, fate and risk assessment of BPA and its substituents in wastewater treatment plant: A review. Environ. Res. 2019, 178, 108732. [Google Scholar] [CrossRef]
- ECHA; Bisphenol, A.F. Substance Infocard. Available online: https://echa.europa.eu/substance-information/-/substanceinfo/100.014.579 (accessed on 15 July 2023).
- Frankowski, R.; Zgoła-Grześkowiak, A.; Smułek, W.; Grześkowiak, T. Removal of bisphenol A and its potential substitutes by biodegradation. Appl. Biochem. Biotechnol. 2020, 191, 1100–1110. [Google Scholar] [CrossRef]
- Commission of the European Communities. Technical Guidance Document in Support of Commission Directive 93/67/EEC on Risk Assessment for New Notified Substances and Commission Regulation (EC) No 1488/94 on Risk Assessment; Part II; Environmental Risk Assessment; Office for Existing Substances; Office for Official Publications of the European Communities: Luxembourg, 1996. [Google Scholar]
- Li, J.; Wang, Y.; Li, N.; He, Y.; Xiao, H.; Fang, D.; Chen, C. Toxic effects of bisphenol A and bisphenol S on Chlorella pyrenoidosa under single and combined action. Int. J. Environ. Res. Public Health 2022, 19, 4245. [Google Scholar] [CrossRef]
- Yadav, N.; Ahn, H.J.; Kurade, M.B.; Ahn, Y.; Park, Y.K.; Khan, M.A.; Salama, E.S.; Li, X.; Jeon, B.H. Fate of five bisphenol derivatives in Chlamydomonas mexicana: Toxicity, removal, biotransformation and microalgal metabolism. J. Hazard. Mater. 2023, 454, 131504. [Google Scholar] [CrossRef]
- Tišler, T.; Krel, A.; Gerželj, U.; Erjavec, B.; Dolenc, M.S.; Pintar, A. Hazard identification and risk characterization of bisphenols A, F and AF to aquatic organisms. Environ. Pollut. 2016, 212, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Li, W.; Yang, M.; Yang, B.; Li, J. Toxicity and biotransformation of bisphenol S in freshwater green alga Chlorella vulgaris. Sci. Total Environ. 2020, 747, 141144. [Google Scholar] [CrossRef] [PubMed]
- Czarny-Krzymińska, K.; Krawczyk, B.; Szczukocki, D. Toxicity of bisphenol A and its structural congeners to microalgae Chlorella vulgaris and Desmodesmus armatus. J. Appl. Phycol. 2022, 34, 1397–1410. [Google Scholar] [CrossRef]
- Elersek, T.; Notersberg, T.; Kovačič, A.; Heath, E.; Filipič, M. The effects of bisphenol A, F and their mixture on algal and cyanobacterial growth: From additivity to antagonism. Environ. Sci. Pollut. Res. 2021, 28, 3445–3454. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, T.; Yang, X.; Liu, H. Insight into The Aquatic Toxicity and Ecological Risk of Bisphenol B, And Comparison with That of Bisphenol A. Res. Artic. 2021. 22 November 2021, available at Research Square. [Google Scholar] [CrossRef]
- Fabrello, J.; Matozzo, V. Bisphenol analogs in aquatic environments and their effects on marine species—A review. J. Mar. Sci. Eng. 2022, 10, 1271. [Google Scholar] [CrossRef]
- EU. European Union Updated Risk Assessment Report. Environment Addendum of April 2008 (To be Read in Conjunction with Published EU RAR of BPA, 2003) 4,4′-ISOPROPYLIDENEDIPHENOL (Bisphenol-A) Part 1 Environment; European Commission; Joint Research Centre; Institute for Health and Consumer Protection: Luxembourg, 2008. [Google Scholar]
- Wang, Q.; Chen, M.; Shan, G.; Chen, P.; Cui, S.; Yi, S.; Zhu, L. Bioaccumulation and biomagnification of emerging bisphenol analogues in aquatic organisms from Taihu Lake, China. Sci. Total Environ. 2017, 598, 814–820. [Google Scholar] [CrossRef]
- Huang, C.; Wu, L.H.; Liu, G.Q.; Shi, L.; Guo, Y. Occurrence and ecological risk assessment of eight endocrine-disrupting chemicals in urban river water and sediments of South China. Arch. Environ. Contam. Toxicol. 2018, 75, 224–235. [Google Scholar] [CrossRef]
- Yamazaki, E.; Yamashita, N.; Taniyasu, S.; Lam, J.; Lam, P.K.; Moon, H.B.; Jeong, Y.; Kannan, P.; Achyuthan, H.; Munuswamy, N.; et al. Bisphenol A and other bisphenol analogues including BPS and BPF in surface water samples from Japan, China, Korea and India. Ecotoxicol. Environ. Saf. 2015, 122, 565–572. [Google Scholar] [CrossRef]
- Zhao, X.; Qiu, W.; Zheng, Y.; Xiong, J.; Gao, C.; Hu, S. Occurrence, distribution, bioaccumulation, and ecological risk of bisphenol analogues, parabens and their metabolites in the Pearl River Estuary, South China. Ecotoxicol. Environ. Saf. 2019, 180, 43–52. [Google Scholar] [CrossRef]
- Guillard, R.R. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals: Proceedings—1st Conference on Culture of Marine Invertebrate Animals Greenport; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Fon Sing, S. Standard methods for measuring growth of algae and their composition. In Algae for Biofuels and Energy; Borowitzka, M., Moheimani, N., Eds.; Developments in Applied Phycology; Springer: Dordrecht, The Netherlands, 2013; Volume 5, pp. 265–284. [Google Scholar]
- Bradford, M.M. A Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Crapo, J.D.; McCord, J.M.; Fridovich, I. Preparation and assay of superioxide dismutases. Methods Enzymol. 1978, 53, 382–393. [Google Scholar]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-Dithiobis(2-Nitrobenzoic Acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S Transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Janknegt, P.J.; De Graaff, C.M.; Van de Poll, W.H.; Visser, R.J.; Rijstenbil, J.W.; Buma, A.G. Short-term antioxidative responses of 15 microalgae exposed to excessive irradiance including ultraviolet radiation. Eur. J. Phycol. 2009, 44, 525–539. [Google Scholar] [CrossRef]
- Mecocci, P.; Fanó, G.; Fulle, S.; MacGarvey, U.; Shinobu, L.; Polidori, M.C.; Cherubini, A.; Vecchiet, J.; Senin, U.; Beal, M.F. Age-dependent increases in oxidative damage to DNA, lipids, and proteins in human skeletal muscle. Free Radic. Biol. Med. 1999, 26, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Damiens, G.; Gnassia-Barelli, M.; Loquès, F.; Roméo, M.; Salbert, V. Integrated biomarker response index as a useful tool for environmental assessment evaluated using transplanted mussels. Chemosphere 2007, 66, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Azizullah, A.; Khan, S.; Gao, G.; Gao, K. The interplay between bisphenol A and algae–A review. J. King Saud Univ. Sci. 2022, 34, 102050. [Google Scholar] [CrossRef]
- M’Rabet, C.; Pringault, O.; Zmerli-Triki, H.; Gharbia, H.B.; Couet, D.; Yahia, O.K.D. Impact of two plastic-derived chemicals, the Bisphenol A and the di-2-ethylhexyl phthalate, exposure on the marine toxic dinoflagellate Alexandrium pacificum. Mar. Pollut. Bull. 2018, 126, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Naveira, C.; Rodrigues, N.; Santos, F.S.; Santos, L.N.; Neves, R.A. Acute toxicity of Bisphenol A (BPA) to tropical marine and estuarine species from different trophic groups. Environ. Pollut. 2021, 268, 115911. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Hou, S.; Huang, Y.; Chen, H.; Sun, Z.; Chen, D. Exposure to bisphenol analogues interrupts growth, proliferation, and fatty acid compositions of protozoa Tetrahymena thermophila. J. Hazard. Mater. 2020, 395, 122643. [Google Scholar] [CrossRef]
- Duan, L.; Chen, Q.; Duan, S. Transcriptional analysis of Chlorella pyrenoidosa exposed to bisphenol A. Int. J. Environ. Res. Public Health 2019, 16, 1374. [Google Scholar] [CrossRef]
- Zhang, W.; Xiong, B.; Sun, W.F.; An, S.; Lin, K.F.; Guo, M.J.; Cui, X.H. Acute and chronic toxic effects of bisphenol a on Chlorella pyrenoidosa and Scenedesmus obliquus. Environ. Toxicol. 2014, 29, 714–722. [Google Scholar] [CrossRef]
- Li, R.; Chen, G.Z.; Tam, N.F.Y.; Luan, T.G.; Shin, P.K.; Cheung, S.G.; Liu, Y. Toxicity of bisphenol A and its bioaccumulation and removal by a marine microalga Stephanodiscus hantzschii. Ecotoxicol. Environ. Saf. 2009, 72, 321–328. [Google Scholar] [CrossRef]
- Sendra, M.; Moreno-Garrido, I.; Blasco, J. Single and multispecies microalgae toxicological tests assessing the impact of several BPA analogues used by industry. Environ. Pollut. 2023, 333, 122073. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, W.; Huang, X.; Ding, T. Comparative study on the toxicity and removal of bisphenol S in two typical freshwater algae. Environ. Sci. Pollut. Res. 2021, 28, 36861–36869. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, Y.; Chen, G.; Tam, N.F.; Shin, P.K.; Cheung, S.G.; Luan, T. Physiological responses of the alga Cyclotella caspia to bisphenol A exposure. Bot. Mar. 2008, 51, 360–369. [Google Scholar] [CrossRef]
- Yang, M.; Fan, Z.; Xie, Y.; Fang, L.; Wang, X.; Yuan, Y.; Li, R. Transcriptome analysis of the effect of bisphenol A exposure on the growth, photosynthetic activity and risk of microcystin-LR release by Microcystis aeruginosa. J. Hazard. Mater. 2020, 397, 122746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ding, T.; Luo, X.; Li, J. Toxic effect of fluorene-9-bisphenol to green algae Chlorella vulgaris and its metabolic fate. Ecotoxicol. Environ. Saf. 2021, 216, 112158. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guan, Y.; Gao, Q.; Tam, N.F.Y.; Zhu, W. Cellular responses, biodegradation and bioaccumulation of endocrine disrupting chemicals in marine diatom Navicula incerta. Chemosphere 2010, 80, 592–599. [Google Scholar] [CrossRef]
- Ben Ouada, S.; Ben Ali, R.; Leboulanger, C.; Zaghden, H.; Choura, S.; Ben Ouada, H.; Sayadi, S. Effect and removal of bisphenol A by two extremophilic microalgal strains (Chlorophyta). J. Appl. Phycol. 2018, 30, 1765–1776. [Google Scholar] [CrossRef]
- Jiefeng, L.; Xufa, M.; Yu, L.; Shenjuan, Z.; Boli, Z.; Jie, H. Effects of bisphenol A on growth, photosynthesis pigment and antioxidant system of Spirodela polyrrhiza. Asian J. Ecotoxicol. 2019, 6, 144–151. [Google Scholar]
- Xiang, R.; Shi, J.; Yu, Y.; Zhang, H.; Dong, C.; Yang, Y.; Wu, Z. The effect of bisphenol A on growth, morphology, lipid peroxidation, antioxidant enzyme activity, and PS II in Cylindrospermopsis raciborskii and Scenedesmus quadricauda. Arch. Environ. Contam. Toxicol. 2018, 74, 515–526. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Species | Concentration | Effects on Growth | Time | Ref. |
|---|---|---|---|---|---|
| BPA | Chlorella pyrenoidosa | 2, 5, 8, 11, 15 mg/L | 2 mg/L (1, 2, 3 d): increased 2 mg/L (4, 5, 6 d): inhibition 5, 8, 11, 15 mg/L: inhibition | 6 days | [13] |
| BPS | C. pyrenoidosa | 5, 10, 15, 20, 40 mg/L | 5 mg/L (1, 2, 3, 4 d): inhibition 5 mg/L (5, 6 d): increased 10, 15, 20, 40 mg/L: inhibition | 6 days | [13] |
| BPA + BPS | C. pyrenoidosa | 0.05 P, 0.1 P, 0.2 P, 0.3 P, 0.4 P, 0.5 P | 0.05 P (1, 5, 6 d): increased 0.1 P (5, 6 d): inhibition 0.2, 0.3, 0.4, 0.5 P: inhibition | 6 days | [13] |
| BPA | Tetraselmis sp. | 2.34, 4.69, 9.38, 18.75, 37.5, 75, 150, 300 mg/L | Inhibition | 24, 48, 72, 96 h | [42] |
| BPA, BPAF, BPB, BPE | Tetrahymena thermophila | 2.6–13.0 μM | BPA (13 μM): inhibition BPAF (13 μM): inhibition BPB (2.6–13 μM): inhibition BPE (2.6–13 μM): inhibition | 30 h | [43] |
| BPA | C. pyrenoidosa | 0.1, 1, 10 mg/L | 0.1 mg/L (2–5 d): increased 1 mg/ (4–5 d): inhibition 10 mg/L (1–5 d): inhibition | 5 days | [44] |
| BPA | C. pyrenoidosa | 1, 5, 10, 25, 50 mg/L | 1, 5 mg/L (96 h): inhibition 10 mg/L (72, 96, 144 h): inhibition 25 mg/L (48, 72, 96 h): inhibition 50 mg/L (24, 48, 72, 96 h): inhibition | 144 h | [45] |
| BPA | Scenedesmus obliquus | 1, 5, 10, 25, 50 mg/L | 10 mg/L (24, 96 h): inhibition 25 mg/L (24, 48, 72, 96 h): inhibition 50 mg/L (24, 48, 72, 96 h): inhibition | 144 h | [45] |
| BPA | C. pyrenoidosa | 1, 10, 50 mg/L | 1, 50 mg/L (30 d): increased | 30 d | [45] |
| BPA | Stephanodiscus hantzschii | 0.01, 0.1, 1, 3, 5, 7, 9 mg/L | ≤1 mg/L: no effects 3 mg/L (4 d): inhibition 3 mg/L (8, 12, 16 d): increased 5, 7, 9 mg/L: inhibition | 16 d | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabrello, J.; Ciscato, M.; Moschin, E.; Vecchia, F.D.; Moro, I.; Matozzo, V. Can BPA Analogs Affect Cellular and Biochemical Responses in the Microalga Phaeodactylum tricornutum Bohlin? J. Xenobiot. 2023, 13, 479-491. https://doi.org/10.3390/jox13030030
Fabrello J, Ciscato M, Moschin E, Vecchia FD, Moro I, Matozzo V. Can BPA Analogs Affect Cellular and Biochemical Responses in the Microalga Phaeodactylum tricornutum Bohlin? Journal of Xenobiotics. 2023; 13(3):479-491. https://doi.org/10.3390/jox13030030
Chicago/Turabian StyleFabrello, Jacopo, Maria Ciscato, Emanuela Moschin, Francesca Dalla Vecchia, Isabella Moro, and Valerio Matozzo. 2023. "Can BPA Analogs Affect Cellular and Biochemical Responses in the Microalga Phaeodactylum tricornutum Bohlin?" Journal of Xenobiotics 13, no. 3: 479-491. https://doi.org/10.3390/jox13030030