
Micro and Nanoplastic Contamination and Its Effects on Freshwater Mussels Caged in an Urban Area



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mussel Caging Experiments
2.2. Size Exclusion Chromatography of Polystyrene Nanoplastics in Tissues
2.3. Digestion via KOH and Determination of Microplastics via Flow Cytometry
2.4. Measurements of Amyloids
2.5. Determination of Protein Aggregation
2.6. Determination of Aldehydes
2.7. Estimation of Lipid Peroxidation, Total Lipids, and Thiols
2.8. Estimation of Metallothionein Content
2.9. Determination of Glutathione S-Transferase and Alcool Dehydrogenase Activity
2.10. Data Analysis
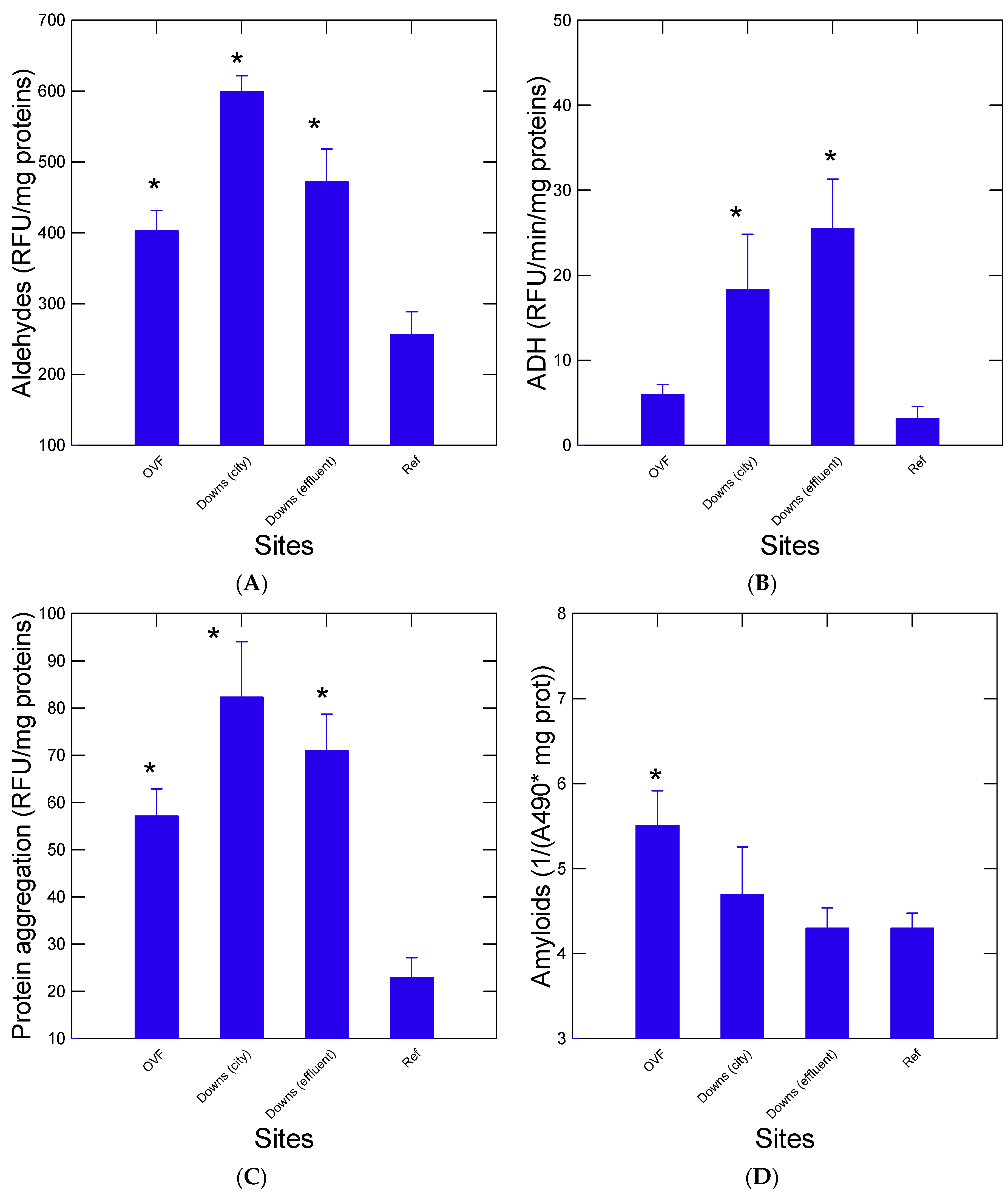
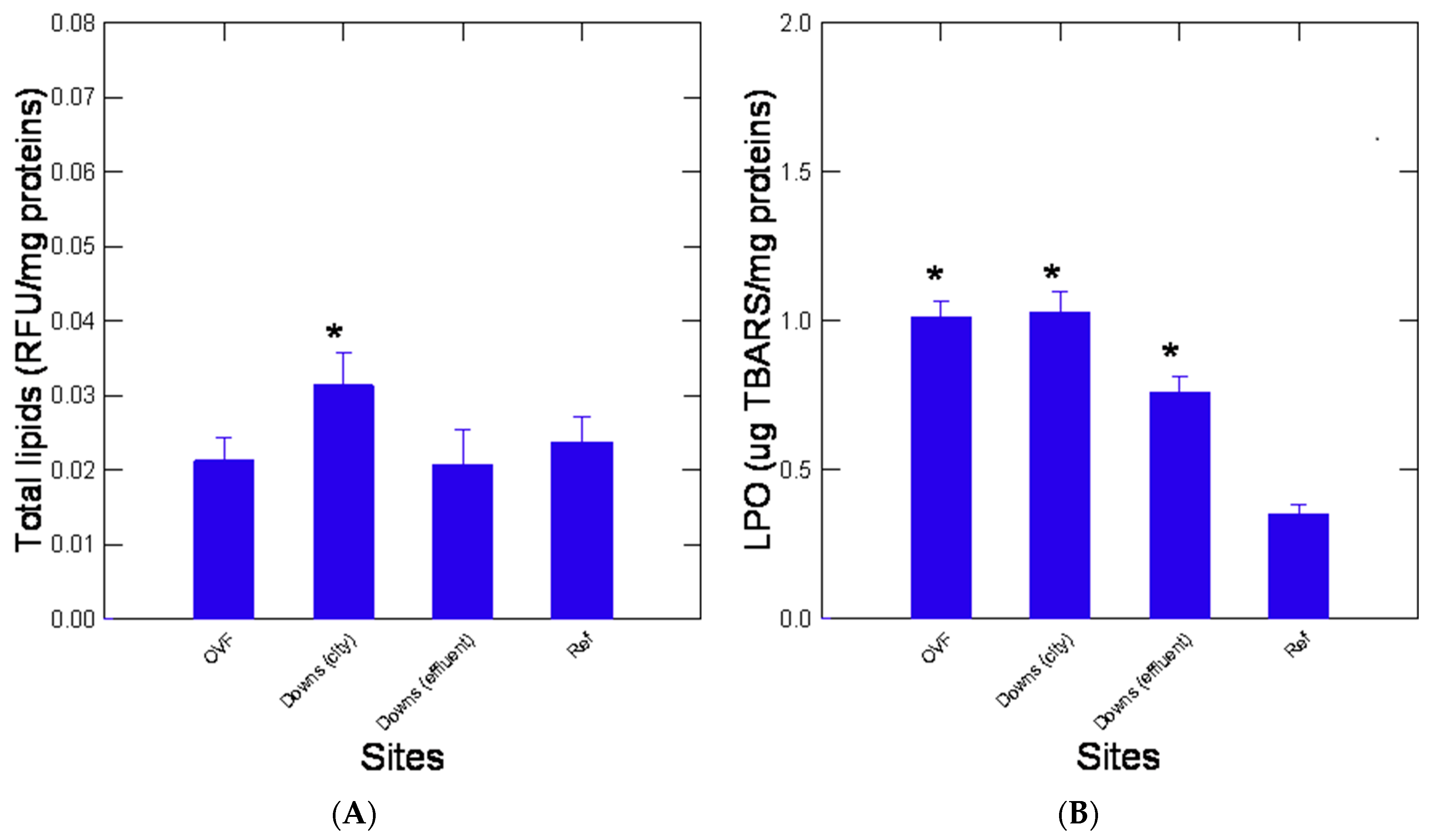
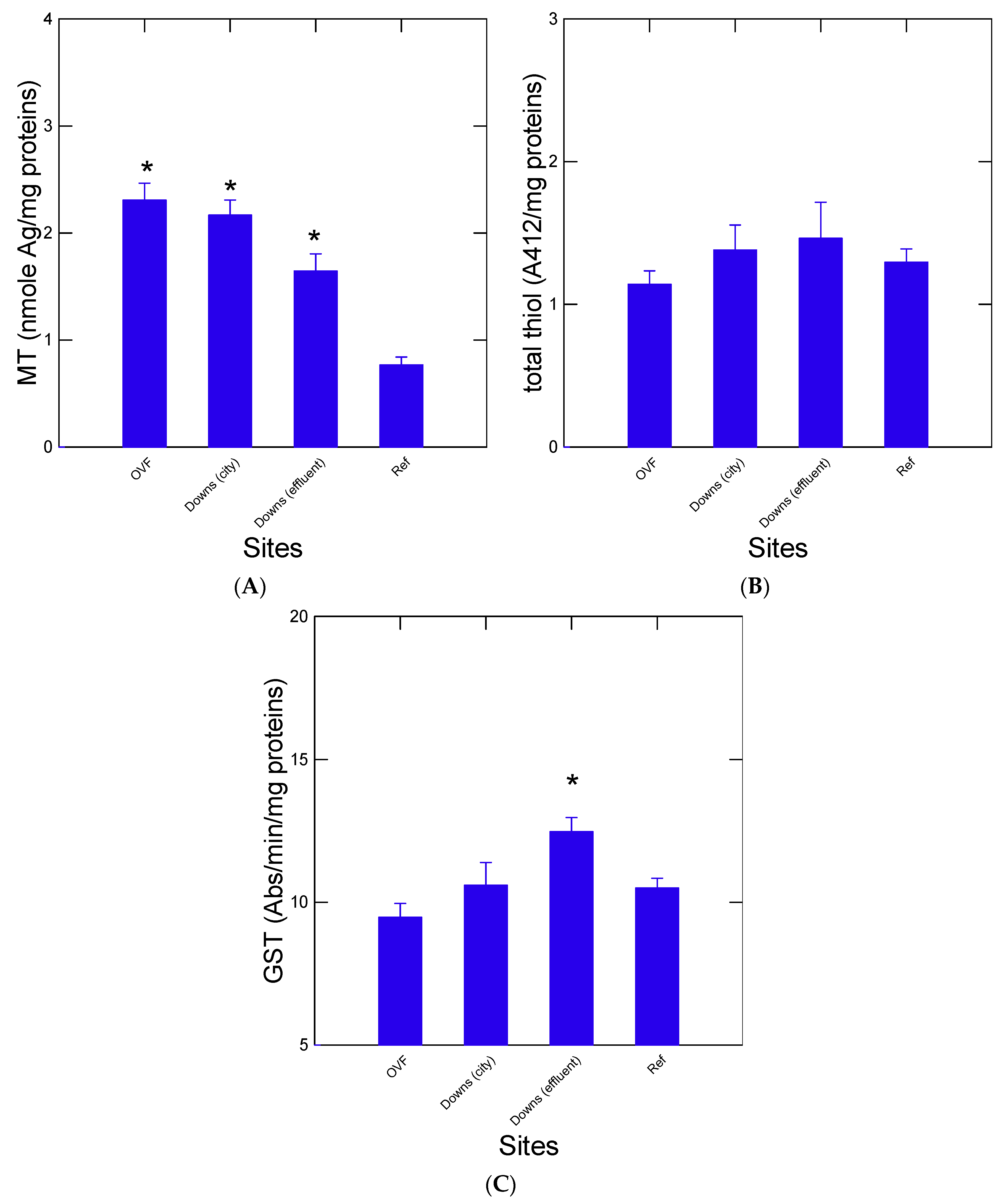
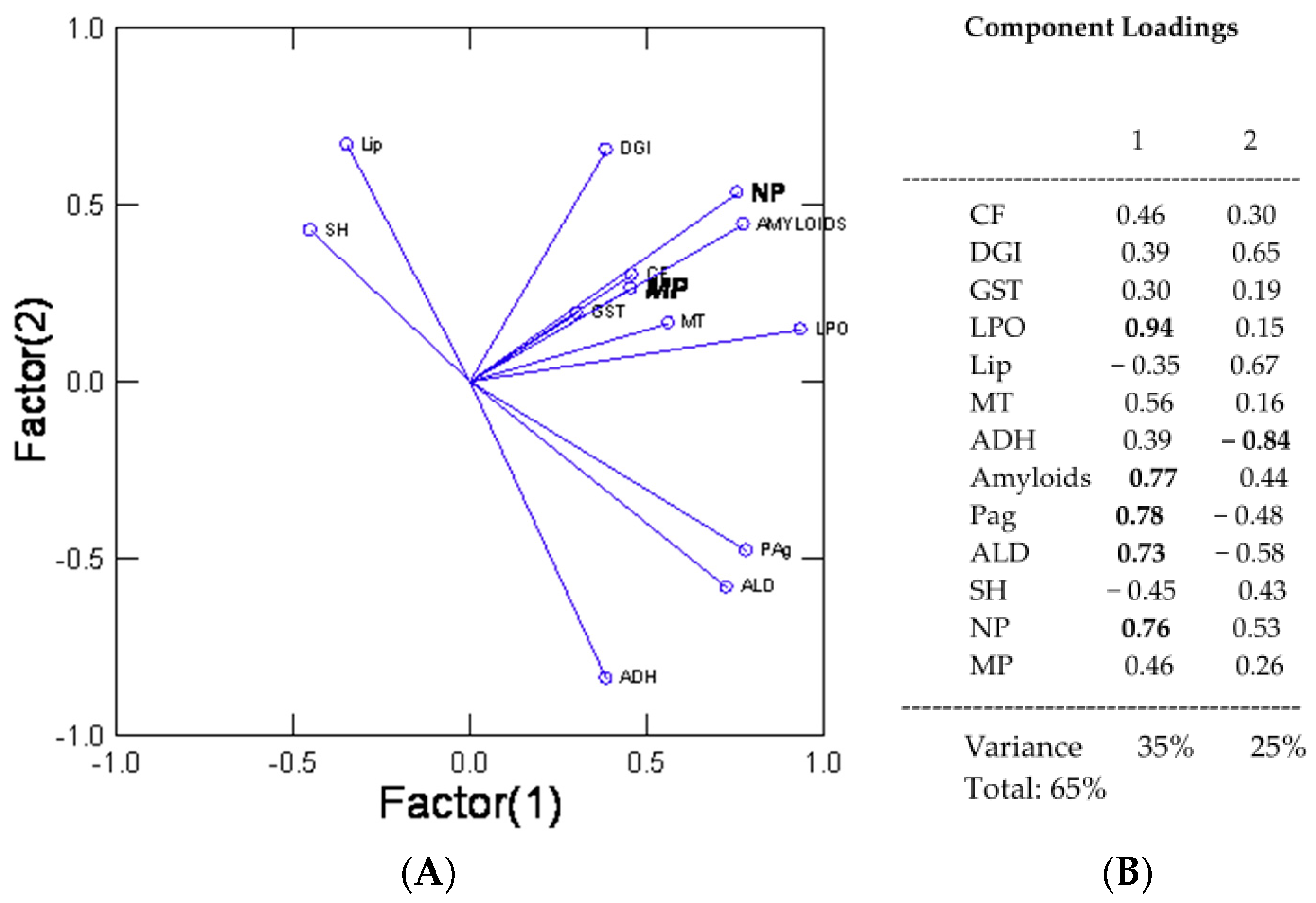
3. Results
Nanoplastics and Microplastics in Mussels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plastic Europe 2021. Plastics—The facts 2022. PlasticEurope 2022, 1, 1–64. [Google Scholar]
- Capricho, J.C.; Prasad, K.; Hameed, N.; Nikzad, M.; Salim, N. Upcycling Polystyrene. Polymers 2022, 14, 5010. [Google Scholar] [CrossRef] [PubMed]
- Roubeau Dumont, E.; Gao, X.; Zheng, J.; Macairan, J.; Hernandez, L.M.; Baesu, A.; Bayen, S.; Robinson, S.A.; Ghoshal, S.; Tufenkji, N. Unraveling the toxicity of tire wear contamination in three freshwater species: From chemical mixture to nanoparticles. J. Hazard. Mater. 2023, 453, 131402. [Google Scholar] [CrossRef]
- Wagner, S.; Hüffer, T.; Klöckner, P.; Wehrhahn, M.; Hofmann, T.; Reemtsma, T. Tire wear particles in the aquatic environment—A review on generation, analysis, occurrence, fate and effects. Water Res. 2018, 139, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Andrady, A.L.; Neal, M.A. Applications and societal benefits of plastics. Philos. Trans. R. Soc. Lond B Biol. Sci. 2009, 364, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Fu, D.; Qi, H.; Lan, C.Q.; Yu, H.; Ge, C. Micro- and nano-plastics in marine environment: Source, distribution and threats—A review. Sci. Total Environ. 2020, 698, 134254. [Google Scholar] [CrossRef] [PubMed]
- Gagné, F. Isolation and quantification of polystyrene nanoplastics in tissues by low pressure size exclusion chromatography. J. Xenobiot. 2022, 12, 109–121. [Google Scholar] [CrossRef]
- Halle, T.; Jeanneau, L.; Martignac, M.; Jardé, E.; Pedrono, B.; Brach, L.; Gigault, J. Nanoplastic in the North Atlantic Subtropical Gyre. Environ. Sci. Technol. 2017, 51, 13689–13697. [Google Scholar] [CrossRef]
- Rowenczyk, L.; Cai, H.; Nguyen, B.; Sirois, M.; Côté-Laurin, M.C.; Toupoint, N.; Ismail, A.; Tufenkji, N.J. From freshwaters to bivalves: Microplastic distribution along the Saint-Lawrence river-to-sea continuum. Hazard. Mater. 2022, 435, 128977. [Google Scholar] [CrossRef]
- Gonçalves, J.M.; Bebianno, M.J. Nanoplastics impact on marine biota: A review. Environ. Pollut. 2021, 273, 116426. [Google Scholar] [CrossRef]
- Wang, S.; Zhong, Z.; Li, Z.; Wang, X.; Gu, H.; Huang, W.; Fang, J.K.-H.; Shi, H.; Hu, M.; Wang, Y. Physiological effects of plastic particles on mussels are mediated by food presence. J. Hazard. Mater. 2021, 404, 124136. [Google Scholar] [CrossRef]
- Guerlet, E.; Vasseur, P.; Giambérini, L. Spatial and temporal variations of biological responses to environmental pollution in the freshwater zebra mussel. Ecotoxicol. Environ. Saf. 2010, 73, 1170–1181. [Google Scholar] [CrossRef]
- Giaccone, G.; Orsi, L.; Cupidi, C.; Tagliavini, F. Lipofuscin hypothesis of Alzheimer’s disease. Dement. Geriatr. Cogn. Dis. Extra 2011, 1, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, S.; Huang, X.; Chen, Y.; Cheng, J.; Zhan, A. Protein-mediated bioadhesion in marine organisms: A review. Mar. Environ. Res. 2021, 170, 105409. [Google Scholar] [CrossRef] [PubMed]
- André, C.; Duy, S.V.; Sauvé, S.; Gagné, F. Comparative toxicity of urban wastewater and rainfall overflow in caged freshwater mussel Elliptio complanata. Front. Physiol. 2023, 14, 1233659. [Google Scholar] [CrossRef]
- Gagné, F. Detection of polystyrene nanoplastics in biological tissues with a fluorescent molecular rotor probe. J. Xenobiot. 2019, 9, 8147–8149. [Google Scholar] [CrossRef] [PubMed]
- Niki, Y.; Matsumoto, H.; Otani, T.; Suda, Y.; Yatabe, T.; Yoshimine, F.; Kondo, M.; Toyama, Y. Flow cytometric technique for the detection of phagocytosed wear particles in patients with total joint arthroplasty. Biomaterials 2003, 24, 3715–3724. [Google Scholar] [CrossRef] [PubMed]
- Piras, M.; Schubert, M.; Zucker-Franklin, D.; Rimon, A.; Franklin, E.C. The characterization of soluble amyloid prepared in water. J. Clin. Investig. 1968, 47, 924–933. [Google Scholar]
- Girych, M.; Gorbenko, G.; Maliyov, I.; Trusova, V.; Mizuguchi, C.; Saito, H.; Kinnunen, P. Combined thioflavin T–Congo red fluorescence assay for amyloid fibril detection. Methods Appl. Fluor. 2016, 4, 034010. [Google Scholar] [CrossRef]
- Cabaleiro-Lago, C.; Quinlan-Pluck, F.; Lynch, I.; Dawson, K.A.; Linse, S. Dual effect of amino modified polystyrene nanoparticles on amyloid β protein fibrillation. ACS Chem. Neurosci. 2010, 1, 279–287. [Google Scholar] [CrossRef]
- Auclair, J.; Quinn, B.; Peyrot, C.; Wilkinson, K.J.; Gagné, F. Detection, biophysical effects, and toxicity of polystyrene nanoparticles to the cnidarian Hydra attenuata. Environ. Sci. Pollut. Res. 2020, 27, 11772–11781. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Wang, S.; Mao, X.; Zhao, X.; Wei, D. An easy and efficient fluorescent method for detecting aldehydes and its application in biotransformation. J. Fluor. 2020, 21, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Wills, E.D. Evaluation of lipid peroxidation in lipids and biological membranes. In Biochemical Toxicology: A Practical Approach; Snell, K., Mullock, B., Eds.; Oxford University Press: Oxford, UK, 1987; pp. 127–152. [Google Scholar]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile red: A selective fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef]
- Butterworth, P.H.W.; Baum, H.; Porter, J.W. A modification of the Ellman procedure for the estimation of protein sulfhydryl groups. Arch. Biochem. Biophys. 1967, 118, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.M.; Cherian, M.G. Quantification of metallothioneins by a silver-saturation method. Toxicol. Appl. Pharm. 1986, 82, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Gagné, F. Metal metabolism and detoxication. In Biochemical Ecotoxicology: Principle and Methods; Elsevier Inc.: New York, NY, USA, 2014; pp. 75–100. [Google Scholar]
- Gowland, B.T.G.; McIntosh, A.D.; Davies, I.M.; Moffat, C.F.; Webster, L. Implications from a field study regarding the relationship between polycyclic aromatic hydrocarbons and glutathione S-transferase activity in mussels. Marine Environ. Res. 2002, 54, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, Y.; Zhang, P.; Mei, R.; Ji, Y.; Zhao, X.; Zhang, Z.; Ma, J.; Chen, L. Quantitative assessment of in vivo distribution of nanoplastics in bivalve Ruditapes philippinarum using reliable SERS tag-labeled nanoplastic models. Nanoscale 2022, 14, 7807–7816. [Google Scholar] [CrossRef]
- Xu, X.; Jian, Y.; Xue, Y.; Hou, Q.; Wang, L. Microplastics in the wastewater treatment plants (WWTPs): Occurrence and removal. Chemosphere 2019, 235, 1089–1096. [Google Scholar] [CrossRef]
- Merzel, R.L.; Purser, L.; Soucy, T.L.; Olszewski, M.; Colón-Bernal, I.; Duhaime, M.; Elgin, A.K.; Holl, M.M.B. Uptake and retention of nanoplastics in Quagga mussels. Glob. Chall. 2019, 4, 1800104. [Google Scholar] [CrossRef]
- Xu, Y.; Ou, Q.; Wang, X.; Hou, F.; Li, P.; van der Hoek, J.P.; Liu, G. Assessing the mass concentration of microplastics and nanoplastics in wastewater treatment plants by pyrolysis gas chromatography-mass Spectrometry. Environ. Sci. Technol. 2023, 57, 3114–3123. [Google Scholar] [CrossRef]
- Wang, S.; Hu, M.; Zheng, J.; Huang, W.; Shang, Y.; Fang, J.K.H.; Shi, H.; Wang, Y. Ingestion of nano/micro plastic particles by the mussel Mytilus coruscus is size dependent. Chemosphere 2021, 263, 127957. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Cai, Y.; Ma, C.; Han, L.; Yang, Z. Combined toxicity of micro/nano scale polystyrene plastics and ciprofloxacin to Corbicula fluminea in freshwater sediments. Sci. Total Environ. 2021, 789, 147887. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M. Understanding the remobilization of copper, zinc, cadmium and lead due to ageing through sequential extraction and isotopic exchangeability. Environ. Monit. Assess. 2016, 188, 381. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hentschel, B.T.; The, S.J. Long-term sorption of metals is similar among plastic types: Implications for plastic debris in aquatic environments. PLoS ONE 2014, 9, e85433. [Google Scholar] [CrossRef]
- Zhang, Q.; He, Y.; Cheng, R.; Li, Q.; Qian, Z.; Lin, X. Recent advances in toxicological research and potential health impact of microplastics and nanoplastics in vivo. Environ. Sci. Pollut. Res. Int. 2022, 29, 40415–40448. [Google Scholar] [CrossRef] [PubMed]
- Farcy, E.; Gagné, F.; Martel, L.; Fortier, M.; Trépanier, S.; Brousseau, P.; Fournier, M. Short-term physiological effects of a xenobiotic mixture on the freshwater mussel Elliptio complanata exposed to municipal effluents. Environ. Res. 2011, 111, 1096–1106. [Google Scholar] [CrossRef]
- Gagné, F.; André, C.; Turgeon, S.; Ménard, N. Evidence of polystyrene nanoplastic contamination and potential impacts in Mya arenaria clams in the Saint-Lawrence estuary (Canada). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 266, 109563. [Google Scholar] [CrossRef]
- Duan, J.; Li, Y.; Gao, J.; Cao, R.; Shang, E.; Zhang, W. ROS-mediated photoaging pathways of nano- and micro-plastic particles under UV irradiation. Water Res. 2022, 216, 118320. [Google Scholar] [CrossRef]
- Woo, S.-H.; Jang, H.; Mun, S.-H.; Lim, Y.; Lee, S. Effect of treadwear grade on the generation of tire PM emissions in laboratory and real-world driving conditions. Sci. Total Environ. 2022, 838, 156548. [Google Scholar] [CrossRef]
- Pocker, Y.; Page, J.D. Zinc-activated alcohols in ternary complexes of liver alcohol dehydrogenase. J. Biol. Chem. 1990, 265, 22101–22108. [Google Scholar] [CrossRef]
- Read, E.; Milford, J.; Zhu, J.; Wu, L.; Bilodeau, M.; Yang, G. The interaction of disulfiram and H2S metabolism in inhibition of aldehyde dehydrogenase activity and liver cancer cell growth. Toxicol. Appl. Pharmacol. 2021, 426, 115642. [Google Scholar] [CrossRef] [PubMed]
- Hellstomn-Lindahl, E.; Oskarsson, A. Response of rat hepatocyte cultures to cadmium chloride and cadmium diethyldithiocarbamate. Toxicology 1989, 56, 9–21. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gagné, F.; Roubeau-Dumont, E.; André, C.; Auclair, J. Micro and Nanoplastic Contamination and Its Effects on Freshwater Mussels Caged in an Urban Area. J. Xenobiot. 2023, 13, 761-774. https://doi.org/10.3390/jox13040048
Gagné F, Roubeau-Dumont E, André C, Auclair J. Micro and Nanoplastic Contamination and Its Effects on Freshwater Mussels Caged in an Urban Area. Journal of Xenobiotics. 2023; 13(4):761-774. https://doi.org/10.3390/jox13040048
Chicago/Turabian StyleGagné, François, Eva Roubeau-Dumont, Chantale André, and Joëlle Auclair. 2023. "Micro and Nanoplastic Contamination and Its Effects on Freshwater Mussels Caged in an Urban Area" Journal of Xenobiotics 13, no. 4: 761-774. https://doi.org/10.3390/jox13040048
APA StyleGagné, F., Roubeau-Dumont, E., André, C., & Auclair, J. (2023). Micro and Nanoplastic Contamination and Its Effects on Freshwater Mussels Caged in an Urban Area. Journal of Xenobiotics, 13(4), 761-774. https://doi.org/10.3390/jox13040048