Silica Nanoparticles Disclose a Detailed Neurodegeneration Profile throughout the Life Span of a Model Organism

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
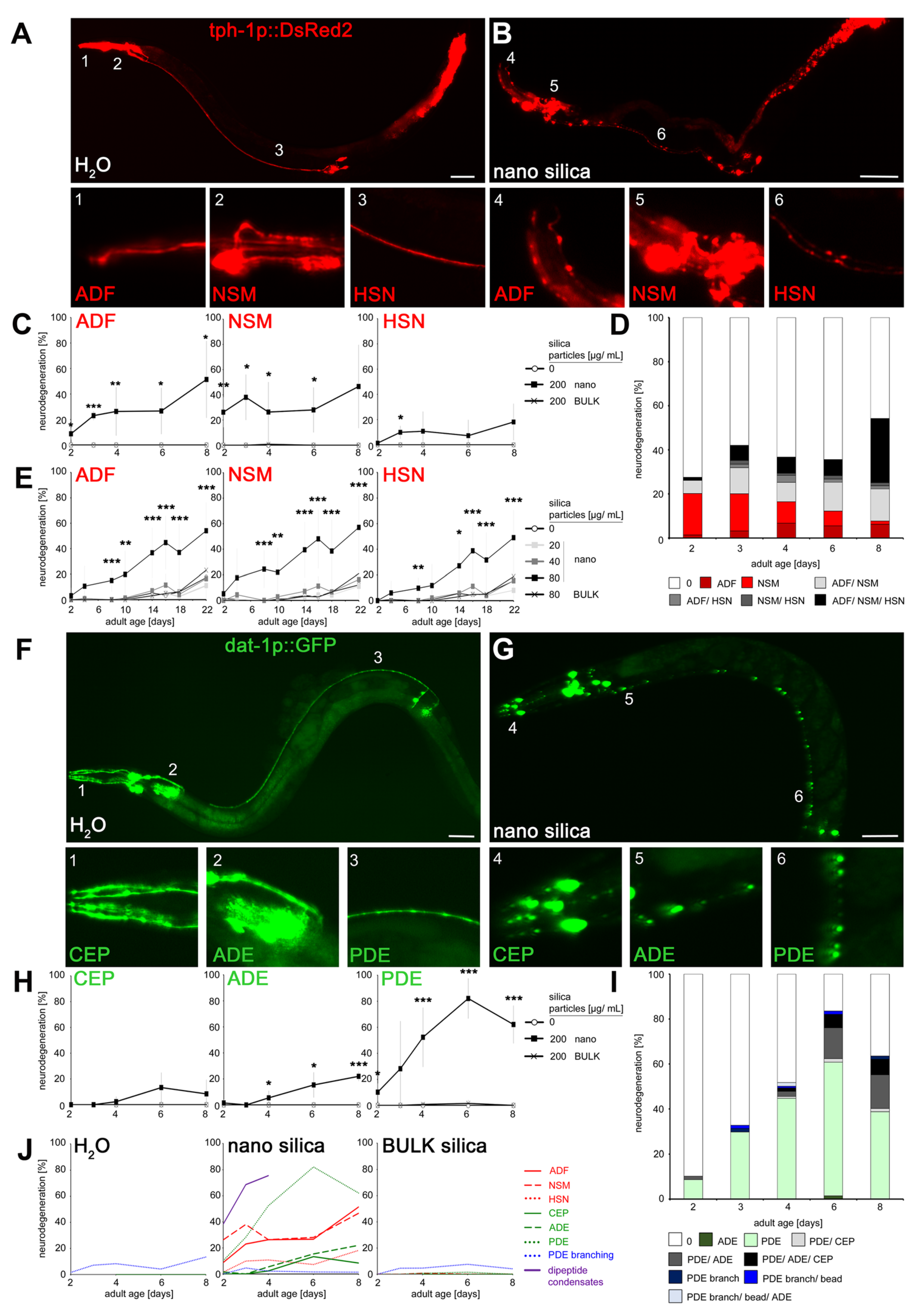
2.1. Chronologic Sequence of Nano Silica-Induced Degeneration in Serotonergic and Dopaminergic Neurons
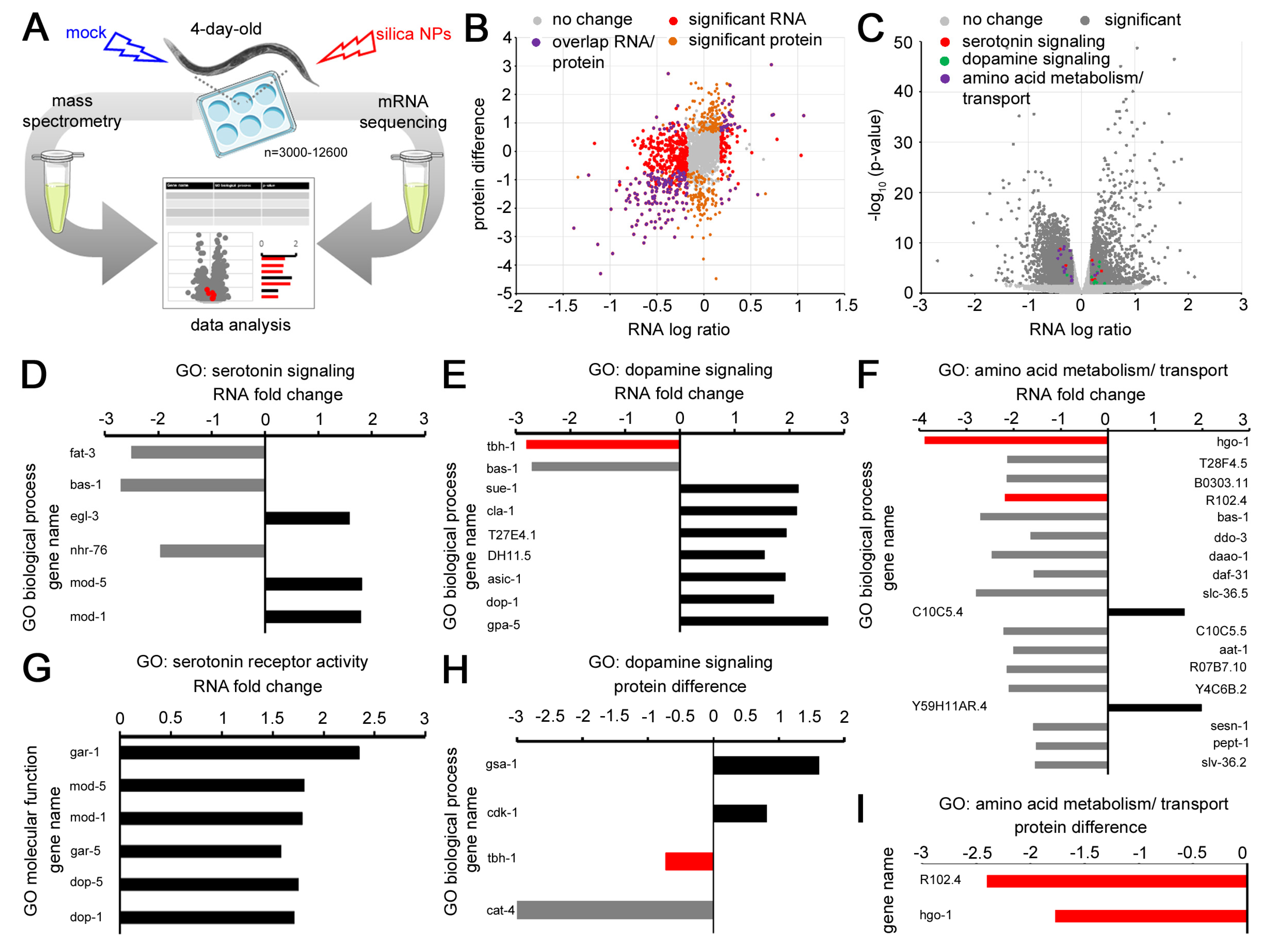
2.2. Gene Expression in 4-Day-Old Worms Exposed to Nano Silica
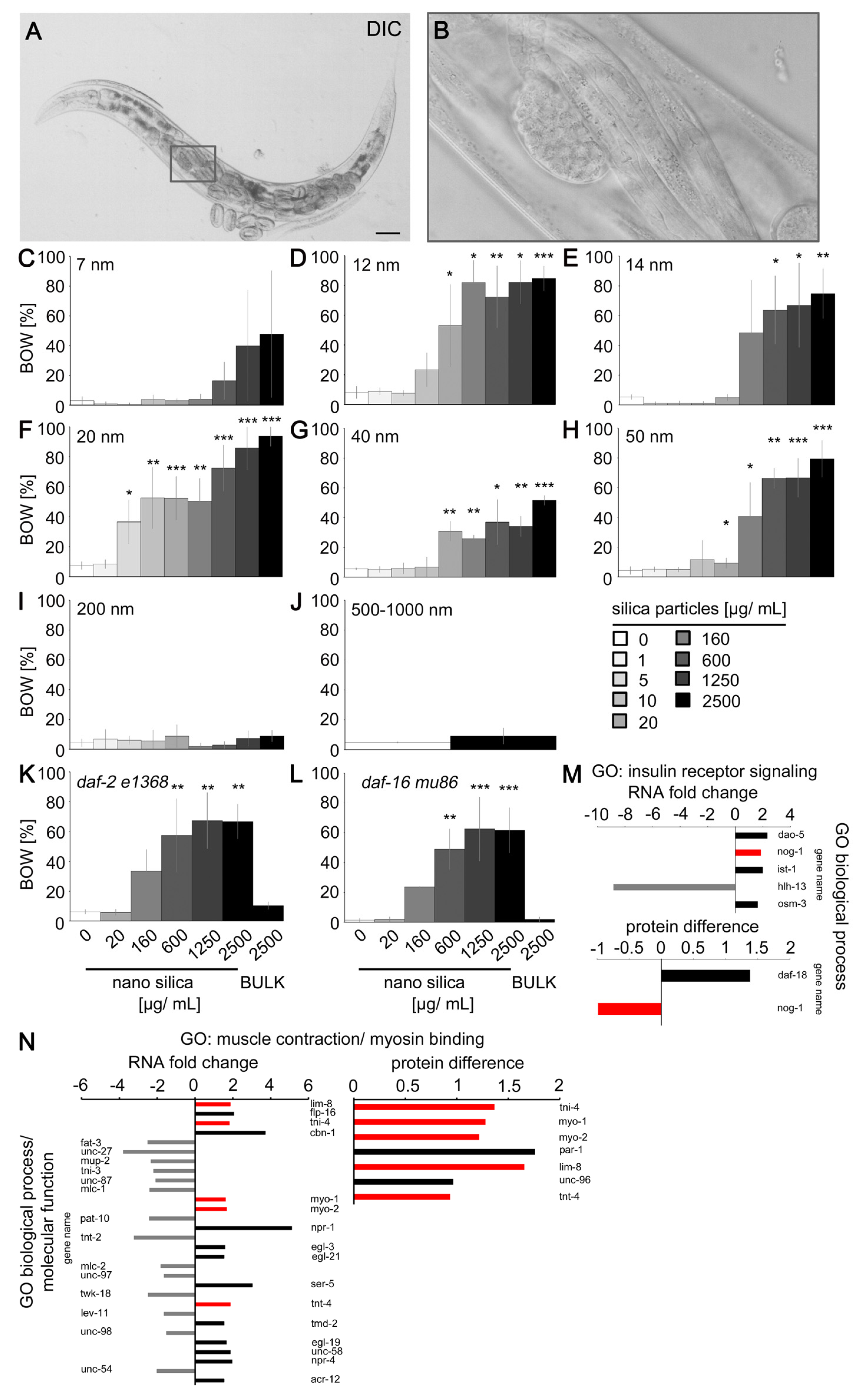
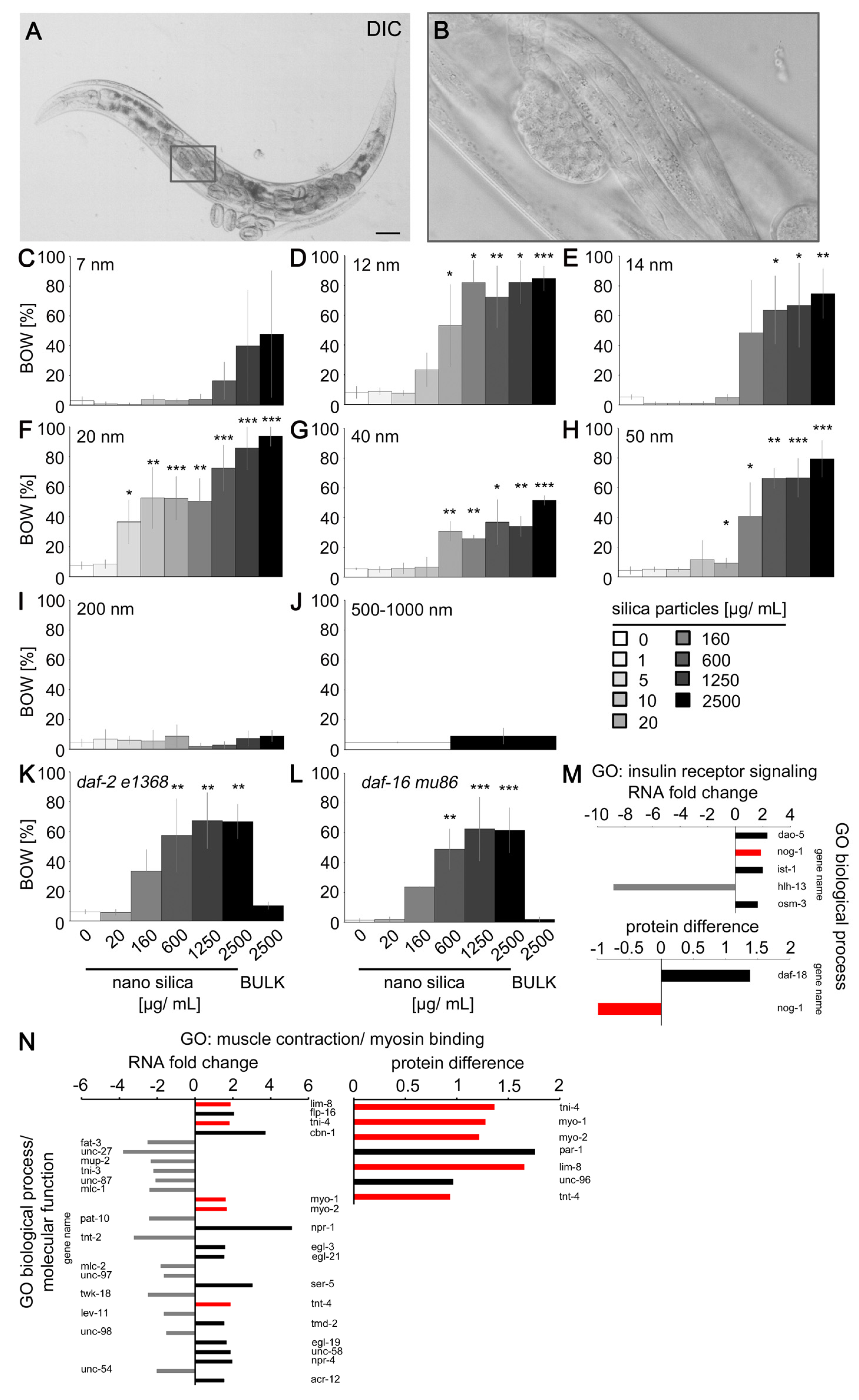
2.3. Silica Nanomaterials Induced the Behavior Defect Internal Hatch
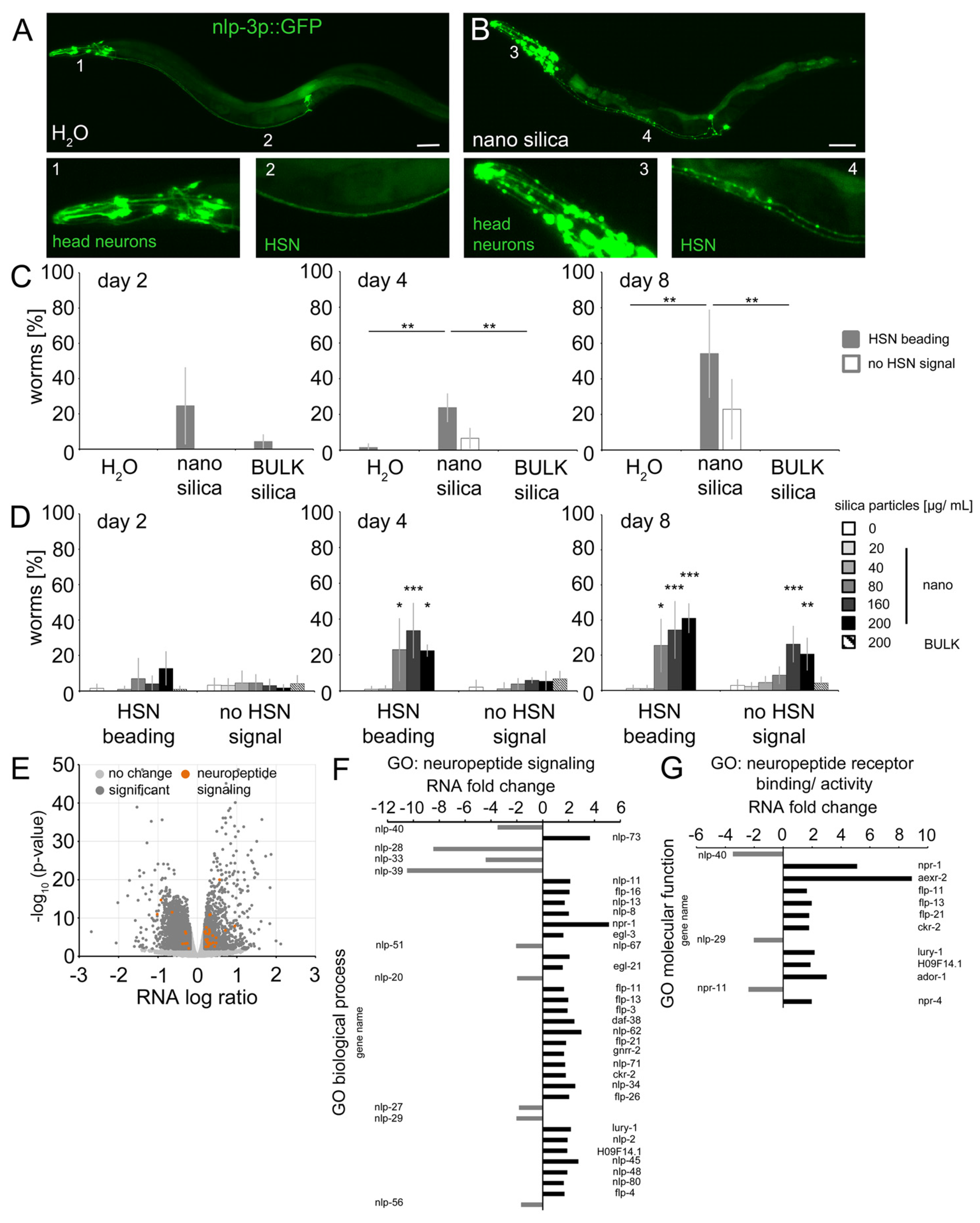
2.4. Nano Silica Induced Relocation of Neuropeptide nlp-3 and Axonal Beading in the Command Neuron HSN
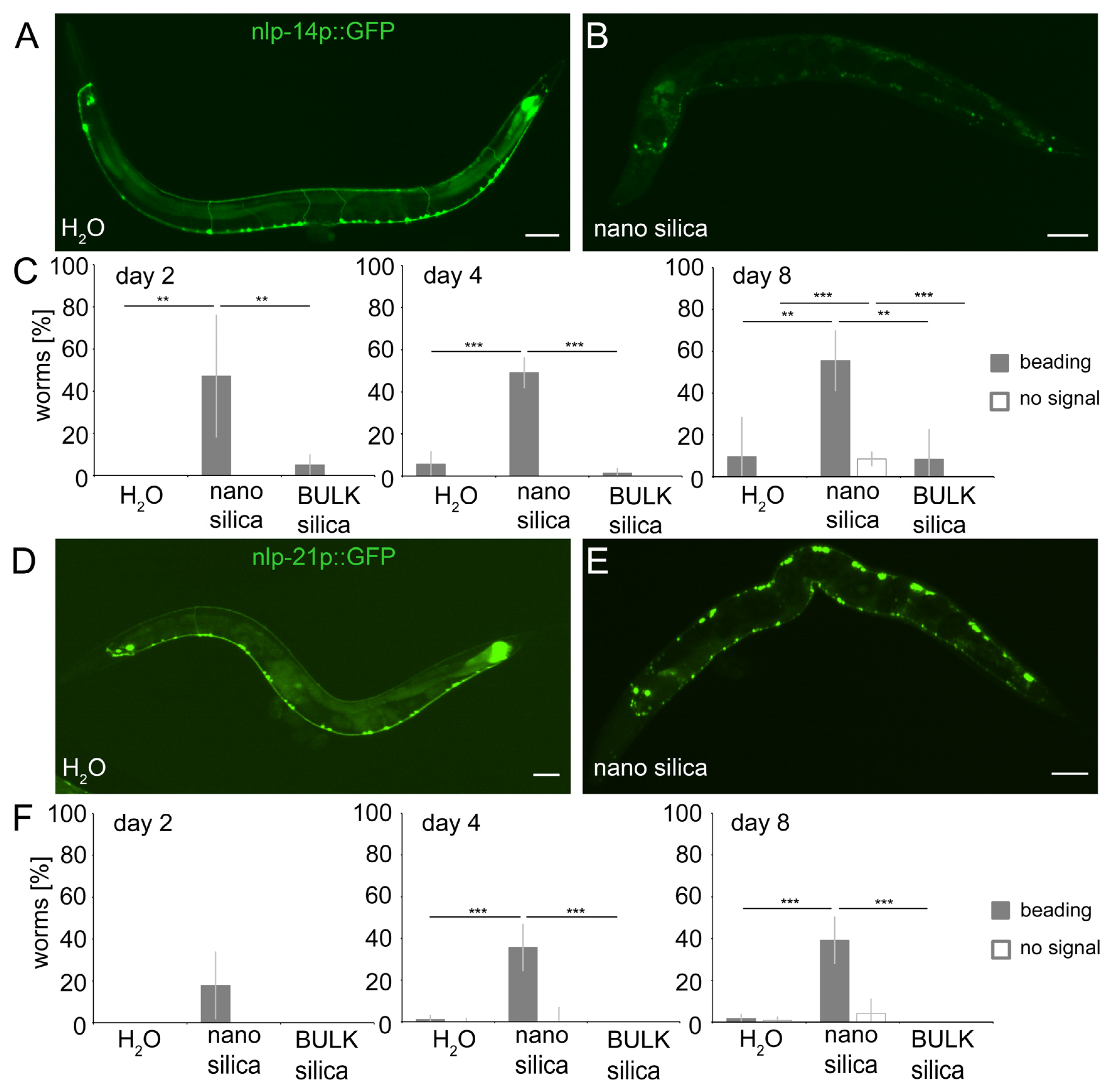
2.5. Nano Silica Induced Relocation of Neuropeptides nlp-14 and nlp-21 as Well as Neural Beading
3. Conclusions
4. Materials and Methods
4.1. Particles
4.2. Caenorhabditis Elegans Strains
4.3. Particle Exposures in Liquid Media
4.4. Microscopy and Quantification of Neurodegeneration
4.5. Microscopy and Quantification of Peptide Condensates
4.6. Internal Hatch
4.7. C. elegans Sample Preparation for Transcriptomic and Proteomic Analyses
4.8. Worm Lysis, Protein Determination and PAA Gel Quality Control
4.9. Protein Sample Preparation for Mass Spectrometric Analysis
4.10. Mass Spectrometric Analysis of the C. elegans Proteome
4.11. RNA Isolation
4.12. RNA Seq Analyses
4.13. Transcriptome and Proteome Analyses
4.14. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Lancet Neurology. Parkinson’s disease needs an urgent public health response. Lancet Neurol. 2022, 21, 759. [Google Scholar] [CrossRef]
- Wilker, E.H.; Osman, M.; Weisskopf, M.G. Ambient air pollution and clinical dementia: Systematic review and meta-analysis. BMJ 2023, 381, e071620. [Google Scholar] [CrossRef]
- Sandner, G.; König, A.; Wallner, M.; Weghuber, J. Alternative model organisms for toxicological fingerprinting of relevant parameters in food and nutrition. Crit. Rev. Food Sci. Nutr. 2022, 62, 5965–5982. [Google Scholar] [CrossRef]
- Zheng, F.; Chen, C.; Aschner, M. Neurotoxicity Evaluation of Nanomaterials Using C. elegans: Survival, Locomotion Behaviors, and Oxidative Stress. Curr. Protoc. 2022, 2, e496. [Google Scholar] [CrossRef]
- Treinin, M.; Jin, Y. Cholinergic transmission in C. elegans: Functions, diversity, and maturation of ACh-activated ion channels. J. Neurochem. 2021, 158, 1274–1291. [Google Scholar] [CrossRef]
- Sawin, E.R.; Ranganathan, R.; Horvitz, H.R. C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 2000, 26, 619–631. [Google Scholar] [CrossRef]
- Waller, A.V. Experiments on the section of the glossopharyngeal and hypoglossal nerves of the frog, and observations of the alterations produced thereby in the structure of their primitive fibres. Philos. Trans. R Soc. 1850, 140, 423–429. [Google Scholar]
- De Vos, K.J.; Grierson, A.J.; Ackerley, S.; Miller, C.C. Role of axonal transport in neurodegenerative diseases. Annu. Rev. Neurosci. 2008, 31, 151–173. [Google Scholar] [CrossRef]
- Merlini, E.; Coleman, M.P.; Loreto, A. Mitochondrial dysfunction as a trigger of programmed axon death. Trends Neurosci. 2022, 45, 53–63. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.F.; Riehn, R.; Ryu, W.S.; Khanarian, N.; Tung, C.K.; Tank, D.; Austin, R.H. In vivo and scanning electron microscopy imaging of up-converting nanophosphors in Caenorhabditis elegans. Nano Lett. 2006, 6, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Scharf, A.; Piechulek, A.; von Mikecz, A. Effect of nanoparticles on the biochemical and behavioral aging phenotype of the nematode Caenorhabditis elegans. ACS Nano 2013, 7, 10695–10703. [Google Scholar] [CrossRef]
- Piechulek, A.; Berwanger, L.C.; von Mikecz, A. Silica nanoparticles disrupt OPT-2/PEP-2-dependent trafficking of nutrient peptides in the intestinal epithelium. Nanotoxicology 2019, 13, 1133–1148. [Google Scholar] [CrossRef]
- Scharf, A.; Gührs, K.H.; von Mikecz, A. Anti-amyloid compounds protect from silica nanoparticle-induced neurotoxicity in the nematode C. elegans. Nanotoxicology 2016, 10, 426–435. [Google Scholar] [CrossRef]
- Taylor, S.R.; Santpere, G.; Weinreb, A.; Barrett, A.; Reilly, M.B.; Xu, C.; Varol, E.; Oikonomou, P.; Glenwinkel, L.; McWhirter, R.; et al. Molecular topography of an entire nervous system. Cell 2021, 184, 4329–4347.e23. [Google Scholar] [CrossRef] [PubMed]
- Scharf, A.; Limke, A.; Guehrs, K.H.; von Mikecz, A. Pollutants corrupt resilience pathways of aging in the nematode C. elegans. iScience 2022, 25, 105027. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Schaeffer, W.J. The synaptic pathology of alpha-synuclein aggregation in dementia with Lewy bodies, Parkinson’s disease and Parkinson’s disease dementia. Acta Neuropathol. 2010, 120, 131–143. [Google Scholar] [CrossRef]
- Fong, S.; Teo, E.; Ng, L.F.; Chen, C.B.; Lakshmanan, L.N.; Tsoi, S.Y.; Moore, P.K.; Inoue, T.; Halliwell, B.; Gruber, J. Energy crisis precedes global metabolic failure in a novel Caenorhabditis elegans Alzheimer Disease model. Sci. Rep. 2016, 6, 33781. [Google Scholar] [CrossRef]
- Alkema, M.J.; Hunter-Ensor, M.; Ringstad, N.; Horvitz, H.R. Tyramine Functions independently of octopamine in the Caenorhabditis elegans nervous system. Neuron 2005, 46, 247–260. [Google Scholar] [CrossRef]
- Zhao, Y.; Hua, X.; Rui, Q.; Wang, D. Exposure to multi-walled carbon nanotubes causes suppression in octopamine signal associated with transgenerational toxicity induction in C. elegans. Chemosphere 2023, 318, 137986. [Google Scholar] [CrossRef]
- Loer, C.M.; Calvo, A.C.; Watschinger, K.; Werner-Felmayer, G.; O’Rourke, D.; Stroud, D.; Tong, A.; Gotenstein, J.R.; Chisholm, A.D.; Hodgkin, J.; et al. Cuticle integrity and biogenic amine synthesis in Caenorhabditis elegans require the cofactor tetrahydrobiopterin (BH4). Genetics 2015, 200, 237–253. [Google Scholar] [CrossRef]
- Tank, E.M.; Rodgers, K.E.; Kenyon, C. Spontaneous age-related neurite branching in Caenorhabditis elegans. J. Neurosci. 2011, 31, 9279–9288. [Google Scholar] [CrossRef] [PubMed]
- Okkema, P.G.; Harrison, S.W.; Plunger, V.; Aryana, A.; Fire, A. Sequence requirements for myosin gene expression and regulation in Caenorhabditis elegans. Genetics 1993, 135, 385–404. [Google Scholar] [CrossRef] [PubMed]
- Gieseler, K.; Qadota, H.; Benian, G.M. Development, structure, and maintenance of C. elegans body wall muscle. WormBook 2017, 2017, 1–59. [Google Scholar] [CrossRef] [PubMed]
- Benian, G.M.; Choo, H.J. From worms to fish to mice. eLife 2020, 9, e57481. [Google Scholar] [CrossRef]
- Wang, Y.; Nowack, B. Dynamic probabilistic material flow analysis of nano-SiO2, nano iron oxides, nano-CeO2, nano-Al2O3, and quantum dots in seven European regions. Environ. Pollut. 2018, 235, 589–601. [Google Scholar] [CrossRef]
- von Mikecz, A. Exposome, Molecular Pathways and One Health: The Invertebrate Caenorhabditis elegans. Int. J. Mol. Sci. 2022, 23, 9084. [Google Scholar] [CrossRef]
- Nathoo, A.N.; Moeller, R.A.; Westlund, B.A.; Hart, A.C. Identification of neuropeptide-like protein gene families in Caenorhabditis elegans and other species. Proc. Natl. Acad. Sci. USA 2001, 98, 14000–14005. [Google Scholar] [CrossRef]
- Li, C.; Kim, K. Neuropeptides. WormBook 2008, 25, 1–36. [Google Scholar] [CrossRef]
- Piechulek, A.; von Mikecz, A. Life span-resolved nanotoxicology enables identification of age-associated neuromuscular vulnerabilities in the nematode Caenorhabditis elegans. Environ. Pollut. 2018, 233, 1095–1103. [Google Scholar] [CrossRef]
- Brewer, J.C.; Olson, A.C.; Collins, K.M.; Koelle, M.R. Serotonin and neuropeptides are both released by the HSN command neuron to initiate Caenorhabditis elegans egg laying. PLoS Genet. 2019, 15, e1007896. [Google Scholar] [CrossRef]
- Zhao, B.; Schafer, W.R. Neuropeptide signaling: From the gut. Curr. Biol. 2013, 23, R481–R483. [Google Scholar] [CrossRef] [PubMed]
- Lin-Moore, A.T.; Oyeyemi, M.J.; Hammarlund, M. rab-27 acts in an intestinal pathway to inhibit axon regeneration in C. elegans. PLoS Genet. 2021, 17, e1009877. [Google Scholar] [CrossRef]
- Matty, M.A.; Lau, H.E.; Haley, J.A.; Singh, A.; Chakraborty, A.; Kono, K.; Reddy, K.C.; Hansen, M.; Chalasani, S.H. Intestine-to-neuronal signaling alters risk-taking behaviors in food-deprived Caenorhabditis elegans. PLoS Genet. 2022, 18, e1010178. [Google Scholar] [CrossRef] [PubMed]
- Nagai, J.; Yu, X.; Papouin, T.; Cheong, E.; Freeman, M.R.; Monk, K.R.; Hastings, M.H.; Haydon, P.G.; Rowitch, D.; Shaham, S.; et al. Behaviorally consequential astrocytic regulation of neural circuits. Neuron 2021, 109, 576–596. [Google Scholar] [CrossRef] [PubMed]
- von Mikecz, A. Elegant Nematodes Improve Our Understanding of Human Neuronal Diseases, the Role of Pollutants and Strategies of Resilience. Environ. Sci. Technol. 2023, 57, 16755–16763. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Tanis, J.E.; Moresco, J.J.; Lindquist, R.A.; Koelle, M.R. Regulation of serotonin biosynthesis by the G proteins Galphao and Galphaq controls serotonin signaling in Caenorhabditis elegans. Genetics 2008, 178, 157–169. [Google Scholar] [CrossRef]
- Nass, R.; Hall, D.H.; Miller, D.M., III; Blakely, R.D. Neurotoxin-induced degeneration of dopamine neurons in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 3264–3269. [Google Scholar] [CrossRef]
- Petrascheck, M.; Ye, X.; Buck, L.B. An antidepressant that extends lifespan in adult Caenorhabditis elegans. Nature 2007, 450, 553–556. [Google Scholar] [CrossRef]
- Brenig, K.; Grube, L.; Schwarzländer, M.; Köhrer, K.; Stühler, K.; Poschmann, G. The Proteomic Landscape of Cysteine Oxidation That Underpins Retinoic Acid-Induced Neuronal Differentiation. J. Proteome Res. 2020, 19, 1923–1940. [Google Scholar] [CrossRef] [PubMed]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Thomas, P.D. PANTHER in 2013: Modeling the evolution of gene function, and other gene attributes, in the context of phylogenetic trees. Nucleic Acid Res. 2013, 41, D377–D386. [Google Scholar] [CrossRef]
- Harris, T.W.; Arnaboldi, V.; Cain, S.; Chan, J.; Chen, W.J.; Cho, J.; Davis, P.; Gao, S.; Grove, C.A.; Kishore, R.; et al. WormBase: A modern Model Organism Information Resource. Nucleic Acids Res. 2020, 48, D762–D767. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limke, A.; Poschmann, G.; Stühler, K.; Petzsch, P.; Wachtmeister, T.; von Mikecz, A. Silica Nanoparticles Disclose a Detailed Neurodegeneration Profile throughout the Life Span of a Model Organism. J. Xenobiot. 2024, 14, 135-153. https://doi.org/10.3390/jox14010008
Limke A, Poschmann G, Stühler K, Petzsch P, Wachtmeister T, von Mikecz A. Silica Nanoparticles Disclose a Detailed Neurodegeneration Profile throughout the Life Span of a Model Organism. Journal of Xenobiotics. 2024; 14(1):135-153. https://doi.org/10.3390/jox14010008
Chicago/Turabian StyleLimke, Annette, Gereon Poschmann, Kai Stühler, Patrick Petzsch, Thorsten Wachtmeister, and Anna von Mikecz. 2024. "Silica Nanoparticles Disclose a Detailed Neurodegeneration Profile throughout the Life Span of a Model Organism" Journal of Xenobiotics 14, no. 1: 135-153. https://doi.org/10.3390/jox14010008