3.1. Average SNP and SBP Systems
Results in this section were obtained using the averages for all farms and years presented in
Table 1.
3.1.1. Evaluation of the Parameters Used
The C content in the aboveground biomass usually ranges between 0.30 and 0.50, and the N content in the aboveground biomass between 0.02 and 0.15, according to pasture chemical analysis [
22] depending on the type and species in the pasture. The parameters we chose are within this range. Regarding the parameter
, literature values for pastures can range between 0.5 and 9. For grasslands in the region of Alentejo, which are classified according to the Moderate Resolution Imaging Spectroradiometer (MODIS) [
40] as “temperate arid shrubland”, the
is 1.063 [
28,
41]. We used 0.98, which besides being consistent with this estimate was obtained from an optimization process using the same input data used in this paper. Finally,
is highly dependent on the feed ingredients and whether the supplemental feed is concentrate or roughage. The most common value is about 20, but for the most common concentrate ingredients the C:N ranges between 15 and 30 (e.g., 15 for olive husk and 29 for maize grain). According to Matschullat et al. [
42], the
in agricultural and grazing areas in the Alentejo region is higher than 18. All parameters used in this paper are within these ranges.
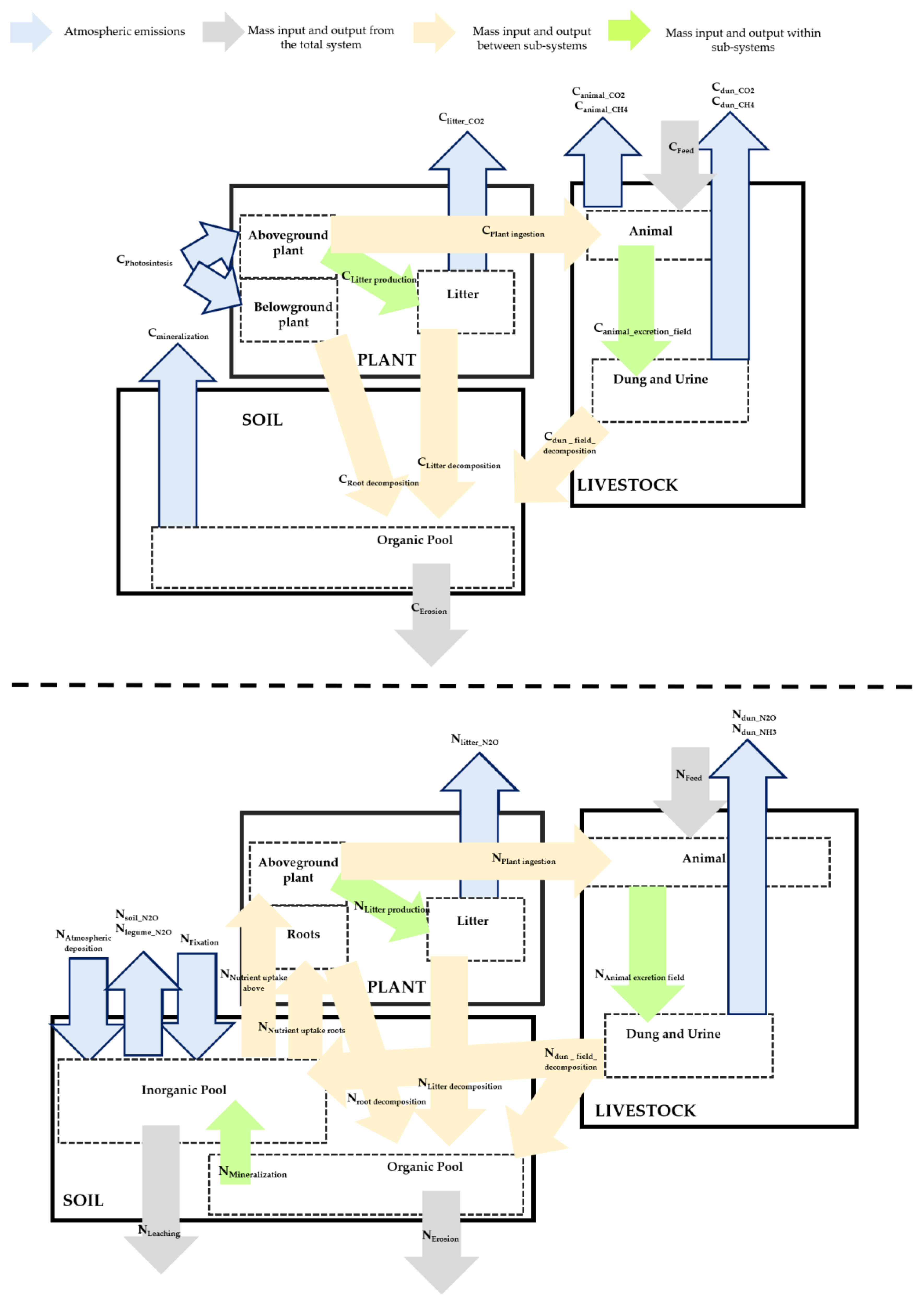
3.1.2. Carbon and Nitrogen Flows
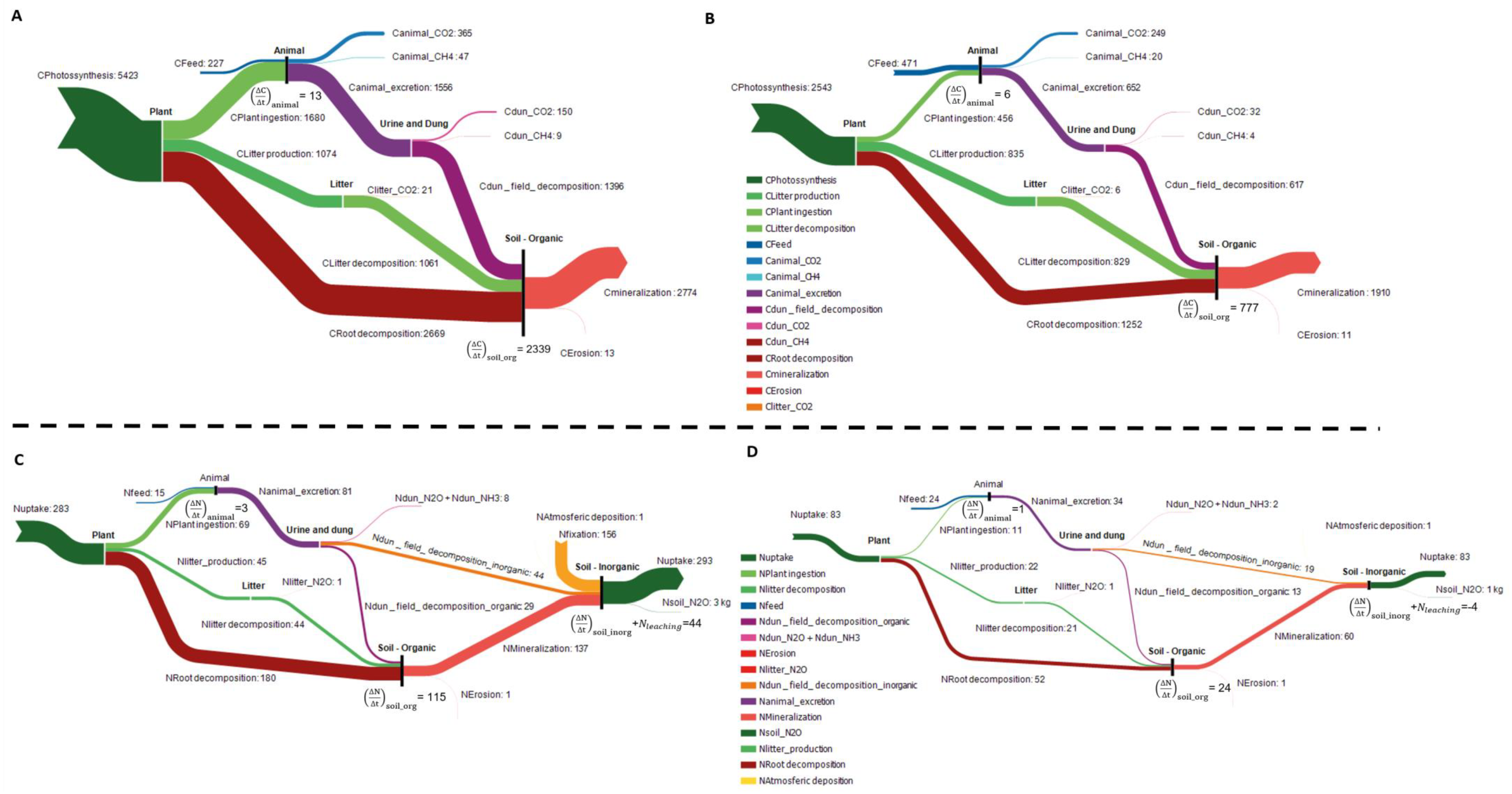
Figure 2 shows that the amount of C and N from the pasture biomass ingested by the animals was, for C, more than triple in SBP compared to SNP (1680 and 456 kgC/ha.yr) and, for N, more than 6 times higher in SBP (69 and 11 kg N/ha.yr). The combined flows of the plant and litter pools were also higher in SBP when compared to SNP. Root decomposition is the main outflow from the plant sub-system–2669 (SBP) and 2543 kgC/ha.yr (SNP) and 180 (SBP) and 51 (SNP) kg N/ha.yr respectively. Litter decomposition was particularly high for SNP in C with a contribution of 829 kgC/ha.yr, while nitrogen was less than half of what was decomposed in the SBP. N
2O (in kg N/ha.yr) emissions represented around 0.2% of the respective N inflow for the plant balance in both pasture systems, thus playing a small part in the mass balance calculation.
Inputs into the animal pool include not only the pasture ingested during grazing, but also the supplemental feed intake, which is higher in the SNP than in the SBP, both in carbon (302 and 471 kg C/ha.yr) and nitrogen (15 and 24 kg N/ha.yr), despite the fact that stocking rates are much lower in SNP. The biggest contributor in terms of mass outflow is the animal excretion through urine and dung, which represents 70–78% of the intake in carbon in SBP and SNP and 96% of the intake in nitrogen. CO2 emissions through respiration are 18% and 27% of the intake in SBP and SNP, while enteric fermentation contributes 2% (in C) in both pasture systems. The accumulation in the animal body was 13 and 6 kg C/ha and 3 and 1 kg N/ha in SBP and SNP respectively.
Emissions of CO2 resulting from the dung and urine excreted represented 11% and 5% of the excreted material in the field in SNP and SBP respectively, and CH4 was 1% for both pasture systems. N2O emissions were also higher in SBP (11%) compared to SNP (6%). However, the main flow was urine and dung that enters the soil organic and inorganic pool, both in C and N.
The soil organic C pool accumulated a total of 2,339 and 777 kg C/ha in SBP and SNP respectively. The main inflow in both systems was C from root decomposition (52% and 46% of the total inflow in SBP and SNP). The same occurred in the organic N pool, with an accumulation of 115 and 24 kg N/ha in SBP and SNP respectively and the root decomposition contribution as high as 71% and 60% in the same systems. The main outflows were the mineralization of SOM in both pasture systems for C (originating CO2 emissions, 2774 and 1910 kg C/ha.yr) and N, and NH4+ that is ready for plant uptake or nitrification (137 and 60 kg N/ha.yr).
In the inorganic N pool there is a major difference between the two systems. SBP gain from a high contribution from legume fixation (159 kg N/ha.yr) and 341 kg N/ha.yr from SOM mineralization, besides the inorganic part of dung and urine that enters the soil. SNP systems only have 80 kg N/ha.yr at their disposal. The outflows of this pool are again for both systems totally dominated by the plant uptake, since N2O emissions from the soil and the legume plants, if existing, represent only 1–2 kg kg N/ha.yr.
It is also visible in
Figure 2 that the largest C and N flows are related with the plant (C input from photosynthesis, N intake from soil, root decomposition into the soil, etc.) or with the soil organic sub-system (accumulation and mineralization). Measured field data was used to estimate all these flows. Flows that were estimated from secondary sources without influence from the data in
Table 1 are relatively small in terms of C and N – although they can be relevant for the results of the GHG balance explained next.
3.1.3. Greenhouse Gases Balance
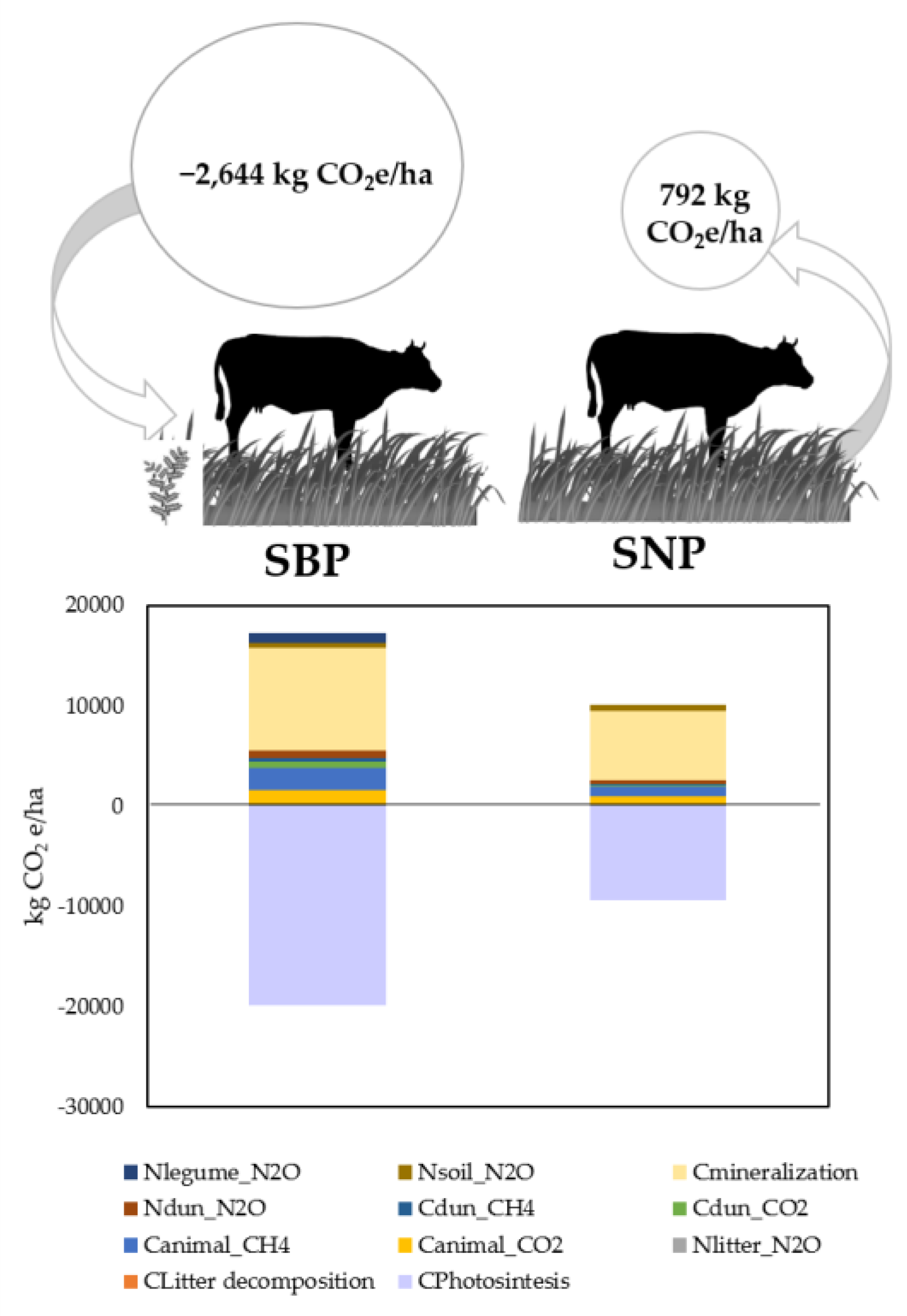
The results for the GHG balance in
Figure 3, show that while the SNP system is a net emitter, with 792 kg CO
2e/ha, the SBP system captured 2644 kg CO
2e/ha. The major contributions for the emissions in both systems are soil mineralization (59% and 69% of the total emissions respectively) and both animal respiration and enteric fermentation representing up to 7–12% in SBP and 9% in SNP. The rest of the emissions individually account for less than 7% of total emissions.
The different result achieved in the two systems is a direct consequence of more than the double SOC accumulation in SBP (8578 kg CO2e/ha.yr) compared to SNP (2850 kg CO2e/ha.yr), while SOM mineralization emissions from SBP represented only 1.5 times the emissions of SNP.
Considering only non-CO
2 emissions, we obtained results shown in
Table 3. The most relevant flow is CH
4 emissions from enteric fermentation in
Table 5, which alone accounts for 43% of all emissions in SBP and 45% in SNP. Emissions per hectare are higher in SBP due to the fact that the stocking rate is more than double than in SNP. However, emissions per LU and live weight of the animal are almost equal (e.g., 17.0 vs. 16.3 kg CO
2e/kg LW).
3.2. Verification of Average Results
The amount of N ingested by the animals according to the N balance is equal for both systems at 0.249 kg N intake/LU.day. This was due to the fact that we assumed that the animals grew by the same amount in both systems. N is connected with important diet parameters such as protein, and as such it should be the same for all systems. Regarding consumption from feed alone, results of the application of the balance revealed that the difference in the feed required in both systems, when converted to DM, is approximately 1962 kg DM/LU.yr. When applied to the SR for SNP, this would result in an extra feed consumption of 769 kg DM/yr, which is equivalent to a 1% supplemental feed intake per day, and therefore within the plausible range for these pastures.
Regarding the closure of the organic C soil balance, if the 13% mineralization rate had been used it would imply that was 2196 and 1788 kg C/ha.yr for SBP and SNP, rather than the 2774 and 1910 kgC/ha.yr obtained from the balance. This represents a deviation that is 11% and 4% of the total C flowing into this pool, respectively, which was probably due to the errors involved not only in the determination of the other variables (parameters used, etc.) but also in the independent estimation of the mineralization rate itself.
As for the inorganic N soil pool, the balance of in and outflows results in a net accumulation plus leaching of inorganic N of 44 kg N/ha.yr for SBP (while the maximum potential was 30 kg N/ha.yr [
38]) and a negative flow of-4 kg N/ha.yr for the SNP (while the expectation was zero). This represents deviations that are 4% and 5% of the total N that flows into this pool, which can also be explained by the error associated with the estimation of the flows.
Regarding the results for the GHG balance, Eldesouky et al. [
43] studied a similar cattle production system in the Spanish
dehesa. In that paper, the
(0.36 LU/ha) is similar to the
in SNP in this paper (0.39 LU/ha). Eldesouky et al. [
43] obtained on-farm emissions of 16.06 kg CO
2e/ha (converting from IPCC AR4 [
44] to IPCC AR5 [
45] with carbon feedback) for an extensive beef/calf cattle farm production system (with calves sold at weaning), which is almost the same obtained here for the non-CO
2 emissions in SNP.
This procedure provides indication that our estimates of those individual flows are aligned with prior results of other studies. We were unable to find additional studies to verify the rest of the flows. However, BalSim imposes that all sub-systems must be balanced, meaning that variables move in tandem in response to changes in parameters. This makes it likely that mistakes in other variables would have affected the variables verified here.
3.3. Carbon and Nitrogen Flows in Different Scenarios
The changes made and results obtained from the scenarios modelled are shown in
Table 6. Regarding question S1, annual SOM increments in SBP increased, compared to the average case presented in 3.1, to 0.55 percentage points per year by doubling yield and stocking rate with no more feed supplementation. In SNP SOM increments decreased (in comparison to results in
Section 3.1 with average yield and stocking rate) to 0.075 percentage points per year. Because of the extra C sequestration in SBP, the GHG balance became more negative. Nevertheless, the non-CO
2 emissions increased to 9685 and 3545 kg CO2e/ha.yr in SBP and SNP, respectively. As the stocking rate is higher, the output in terms of meat produced is also higher and therefore, these emissions, when converted into the amount emitted per kg LW, actually decrease (16.1 and 14.0 kg CO2e/kg LW). The effects of a good agronomic year would therefore be the overall reduction of emissions, regardless of whether or not C sequestration was taken into account.
As for the second question, there are multiple combinations of yield and stocking rate that produce the doubling of average annual SOM increases. One example would be to change yield to 7760 and 3690 kg DM/ha.yr in SBP and SNP respectively, and stocking rates to 1.5 and 0.68 LU/ha.yr. In this case the GHG balance for SBP would become even more negative than in S1, and non-CO2 emissions would decrease for SBP (15.5 kg CO2e/kg LW) and remain the same as in S1 for SNP. However, as the fraction of litter would have to remain high in order to provide high C inputs to soil, this could only be achieved through increased feed supplementation. Ultimately this highlights the trade-off between using plant C (and N) for animal intake or for C input to soils. If the stocking rate is sufficiently large, C input to soil may be insufficient; for C inputs to remain high and enable large SOM increases, either yields also rise or feed supplementation is required.
Finally, the last question S3 was about the consequences of halving the yield and sustaining SOM increases. In this case, we had to make the litter fraction equal to zero to ensure that there was no significant increase of feed supplementation. This seriously compromised the amount of C entering the soil. Consequently, at best the stocking rate could be 0.79 for SBP and 0.39 for SNP. However, the organic C balance does not fully close under these conditions, as it would require a level of SOM mineralization that is 10 times lower than expected. It is therefore very unlikely that half the yield would enable similar SOM increments in any plausible scenario.
3.4. Annual Variation in Different Farms
As shown in
Table 7, in 2001–2002 the productivity of the SBP farms was lower compared to the next year, which had an influence on the C and N intake. However, the year 2002–2003 also had higher stocking rate and therefore there was an approximated adjustment of stocking rates to the yield that meant that, despite large variation in the fraction of litter, the feed supplementation was, on average, relatively stable. Also, the higher stocking rates observed in these intermediate years also translated into higher CO
2 and CH
4 emissions from respiration and enteric fermentation. In the last two years (2003–2004 and 2004–2005) there were higher CO
2 emissions from soil mineralization as a consequence of a high difference between the input of organic carbon in soil—translated by the root, litter and dung and urine contributions and the accumulation of organic carbon in soil. The last year was a drought year, and consequently the stocking rates decreased particularly in SNP – but non-CO
2 emissions per LU nevertheless increased.
In the first and last years there was also accumulation of inorganic nitrogen, while in the middle years, inorganic nitrogen accumulation was slightly negative, suggesting that during this period, all nitrogen was used efficiently in the pasture system. In the first year, which was the year when the pasture was sown, plants seemed to have grown more above than belowground, as shown by the low
in
Table 6.
3.5. Limitations and Further Developments of the Model
BalSim was developed specifically to deal with situations of limited data availability. SBP have been and continue to be the subject of multiple research studies, but it is difficult to simultaneously collect all data required to use full process-based models for C and N in all sub-systems considered here. The modelling approach in this paper enabled us to use a limited amount of field data complemented by literature data. We only had four variables/flows measured, and the rest had to be inferred. Through the restrictions imposed by the mass balance formulation, we obtained valid results that we also showed to be useful for joint agronomic and environmental management of the pastures. The limited amount of field data used was therefore not a weakness of the study, but rather the main reason why we developed BalSim.
Nevertheless, future applications would benefit from a more comprehensive set of field measurements, particularly when they are critical in calculations. The model requires a limited number of parameters, but some of them (such as
) are time-consuming, labor intensive and expensive to measure on the field (field travels, pasture cutting and laboratorial analysis) [
46,
47,
48]. Alternative methods to obtain such data have been proposed elsewhere, e.g., using remote sensing imaging for pasture productivity [
49,
50], and also to obtain indirect measures of SOM concentrations and increments [
51,
52]. Indirect methods have their own limitations and challenges. One of the most relevant problems for application in SBP an SNP area is tree cover. SBP and SNP in “Montado” regions are agri-forestry systems characterized by relatively high tree cover, which makes data collection more difficult, as trees also influence productivities and SOM content and increments [
53,
54]. Our work also showed that knowing the C and N contents of plant tissue and the C:N ratios of soils are critical, as they are determinant for most sub-systems and influence directly the estimation of the larger flows. We used averages and/or ad-hoc adjustments here, but in future applications, if any data additionally to
Table 1 can be collected, these are the parameters that should be given priority.
As the goal of the present paper was to present the model and demonstrate its application potential, and in order to make the model manageable in terms of data requirements, we needed several simplifications that should be reassessed in future. For example, BalSim as applied here has no process basis for the relationships between variables. It is a simple mass balance model that ensures C and N are fully distributed among the most important sub-systems, but it does not provide a mechanistic relationship between the variables. This was a deliberate choice made to test the simplest possible approach to modelling these pasture systems and ensure that BalSim requires the minimum possible information to produce meaningful results. As a future development, the correlation between variables should be explicitly modelled, without disregarding the fact that BalSim should strive to keep field data requirements to a bare minimum.
The issue of independence of data sources used for the parameters introduces particularly high uncertainty in the estimation of atmospheric emissions. This calculation relies mostly on independent emissions factors that are unaffected by the measured data. For example, we used IPCC [
28] Tier 1 emission factors to calculate N
2O emissions from urine and faces during grazing. Several Tier 3 models have been proposed, as for example for New Zealand [
55,
56,
57]. These particular models calibrated at the national level were not suited for this study, as they were developed for sites and animal breeds significantly different from the ones in this paper. Direct applications of Tier 3 models without any recalibration could provide substantially worse estimates of emissions that are even more uncertain than the simple use of emission factors. Calibrations of Tier 3 models for Portugal will be welcome future additions. Emissions from animal excreta can then be modelled as a function of the nitrogen content of the feed and the climate region.
We also used a default value from IPCC (1996) [
31] to estimate the N
2O emissions from legumes. However, the more recent IPCC (2006) report [
28] does not consider this emission due to the lack of evidence that legumes contribute significantly to increasing soil N
2O emissions [
58]. Here too, future versions of BalSim should consider whether N
2O emissions increase due to legumes in SBP in Portugal. One potential approach is to model emissions depending on the site conditions (e.g., soil characteristics and precipitation).
Because of our modelling approach, particularly the assumption that parameters are constant, we believe that within a 1-axis simple-complex spectrum of models, this first version of BalSim would be closer to the simpler models. However, in the foreseeable future BalSim can easily move closer to the middle, namely by modelling the most important flows, rather than assuming them constant, and introducing co-dependencies between parameters. This can be achieved by coupling specific models for particular parameters to BalSim, namely simpler process-based models. For example, the RothC [
33,
59] model could potentially be coupled to estimate soil organic carbon changes and soil mineralization depending explicitly on pasture productivity and excretion during grazing as C inputs to soil. Despite possible improvements achieved by introducing process-based models, the data required should not increase considerably, otherwise the advantage of this model (particularly its simplicity) could be quickly lost.
Regarding the variables estimated here, soil respiration, used to assess the closure of the balance, was particularly important as it was the main outflow from the soil C pool. Soil respiration was estimated using the average SOM concentration and respiration factor obtained from SBP modelling [
7]. SOM accumulation is a fast process in SBP, and it is also probably linked to annual climate conditions. This suggests that follow-up work should take into account a temporal dimension in the analysis. Soil bulk density, which is necessary to convert SOM concentration (kg SOM/kg soil) into the amount of C stored in each unit area (t C/ha), was assumed to be similar in all systems and locations. Consequently, there was a significant error not just in the amount of C lost from respiration but also in the target or reference value that we assumed for balancing the model. This should be fixed in future versions of this model.
Another limitation was that we manually calibrated parameters such as
in order to ensure that the ratio between inorganic and organic N from animal sources was 60–40%. In the future this procedure can be automated using a machine learning (optimization process) where all the parameters vary independently within a plausible range. Different sets of parameters would lead to a different closure of the balances, and the optimum solution of the problem would be the set that leads to a higher balance closure upon multiple iterations in a Monte Carlo procedure. Nevertheless, the solutions of optimization processes are highly dependent on how many parameters are indeterminate. If too many parameters are left free to vary independently, the optimization process could lead to multiple optima [
21]. This procedure would, however, enable an estimation of uncertainty and confidence intervals for all variables estimated—which, using the method in this paper, we were unable to do. Uncertainty is a key aspect to improve in future versions of this work. The source data in
Table 1 and in S1 display considerable spatial and temporal variability. The reliance of literature data for the parameters also introduces additional uncertainty. Calculating uncertainty for these results would require more than simply assigning confidence intervals to each parameter, as their correlations require them to vary in tandem. Additionally, the mass balance approach mitigates some of the potential uncertainty in final results, as flows are interconnected even though the parameters may have been obtained from independent sources. Each sub-system must be balanced simultaneously, which reduces the potential variability. The Monte Carlo approach, taking into account the interdependency of the parameters and variables, is the most appropriate procedure to calculate uncertainties in future versions of the model.
3.6. Future Model Applications
The mass balance approach developed here was used to estimative GHG for two pasture systems where different management decisions were applied. However, this model was developed to be fully applicable to other farming systems where mass balances for carbon and nitrogen are important. BalSim may be used to estimate the GHG balance but also to estimate any C or N flow that is significant in a specific farming system as in the case of N leaching, which is difficult to estimate otherwise. This approach is therefore an intermediate response between broad estimates of C and N flows and very detailed calculations often dependent on too many and locally specific parameters. The model is, however, limited by the number of unknowns that can be estimated through the mass balance, since within each sub-system there is only the possibility to have one unknown, while the other flows must be estimated using measured data or resorting to the literature as we did in this work.
This modelling approach could also be applied for annual or permanent crops if the model was sufficiently adapted. Those applications could include the modelling of C and N flows in annual and permanent lands where different management practices are in use with potential to maximize the CO2 sequestration or minimize nitrogen leaching. In that case, the animal sub-systems would no longer apply. Primary production would instead be exported from the farm as a product.
One of the main advantages of BalSim is the simplicity of the processes it depicts, which makes it a feasible option to calculate C and N balances at higher levels, e.g., at the regional level such as the European Union, or even at a global scale. The applications at those scales are difficult for process-based models due to their complexity of processes and the data requirements. For example, the Biome-BGC model [
60] requires data such as transpiration of soil water through leaf stomata. Keeping the model simple and using a limited number of parameters and data required is one of the advantages of this model when compared with others.
We applied BalSim here using field data for yield, stocking rate and SOM accumulation in SBP and SNP. The model could potentially be generalized for wider regions using other data sets, namely for aboveground productivity (such as available net primary production data sets [
61,
62]) and stocking rates (such as the Livestock Geo-Wiki data set [
63]). Despite of the utility of these datasets, there could potentially be data consistency problems due to different assumptions and methodological choices in their construction. Specific systems such as SBP would be very difficult to assess using those data that fail to grasp the specificity of the system (such as higher productivity and stocking rates) and only catch general trends and “generic” systems (e.g., land use data [
61] used in the net primary production data [
61,
62] only considers one “grazing” class).
BalSim can also be expanded in cases where animals are housed for part of the year. In our case, we assumed full time grazing. If cattle had spent a fraction of the year indoors, the C and N excreted during housing would need to be subtracted from the terms, as they would be exported from the field and never reach the soil. Then, that amount of C and N would undergo more transformation, and a new pool (the “manure” pool) would have to be considered. That pool has its own emissions from storage and treatment of manures, but it could potentially also be recirculated into the field if the manure was then used as organic fertilizer in the same pasture plots (also with associated emissions).








{kind=link}
{kind=link}
{kind=link}