Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site, Species and Cultivars Selection, and Drought Experiment
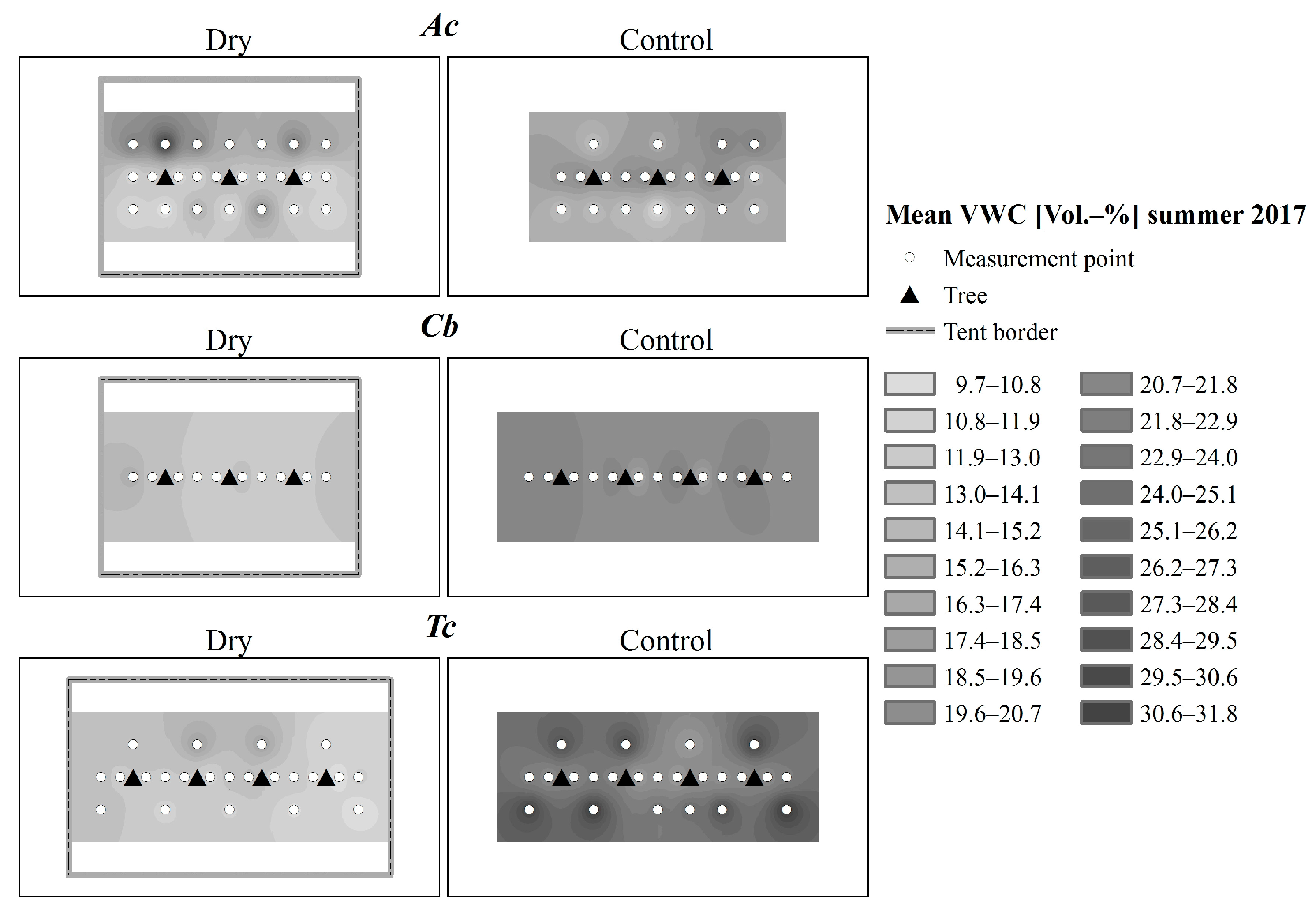
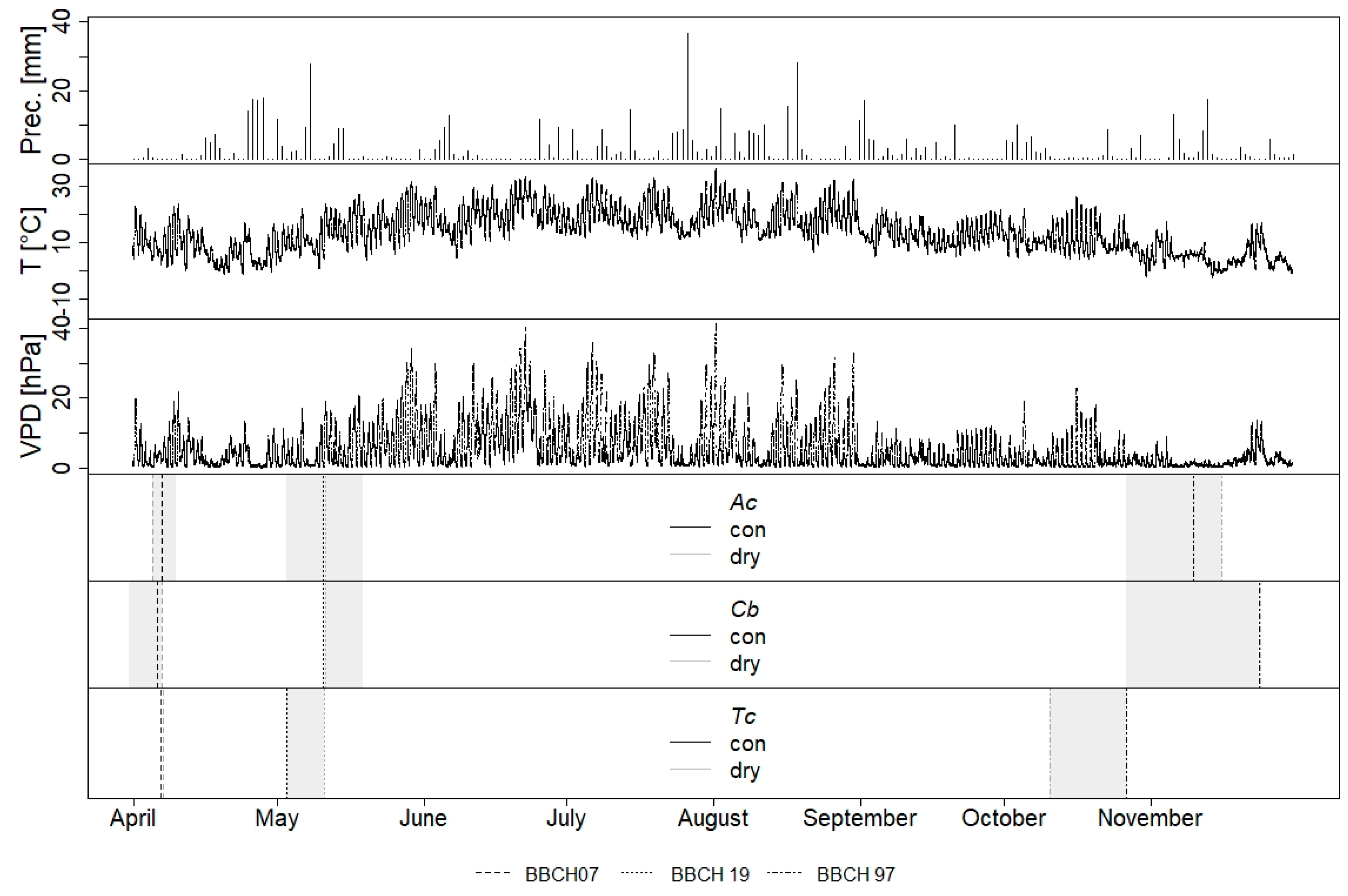
2.2. Climate and Soil Moisture
2.3. Spring and Autumn Phenology
2.4. Optical Measurements of Crown and Root System Architecture
2.5. Stem Growth Development, Excavation, and Dry Weight
2.6. Statistical Analysis
3. Results
3.1. Environmental Conditions
3.2. Leaf Phenology
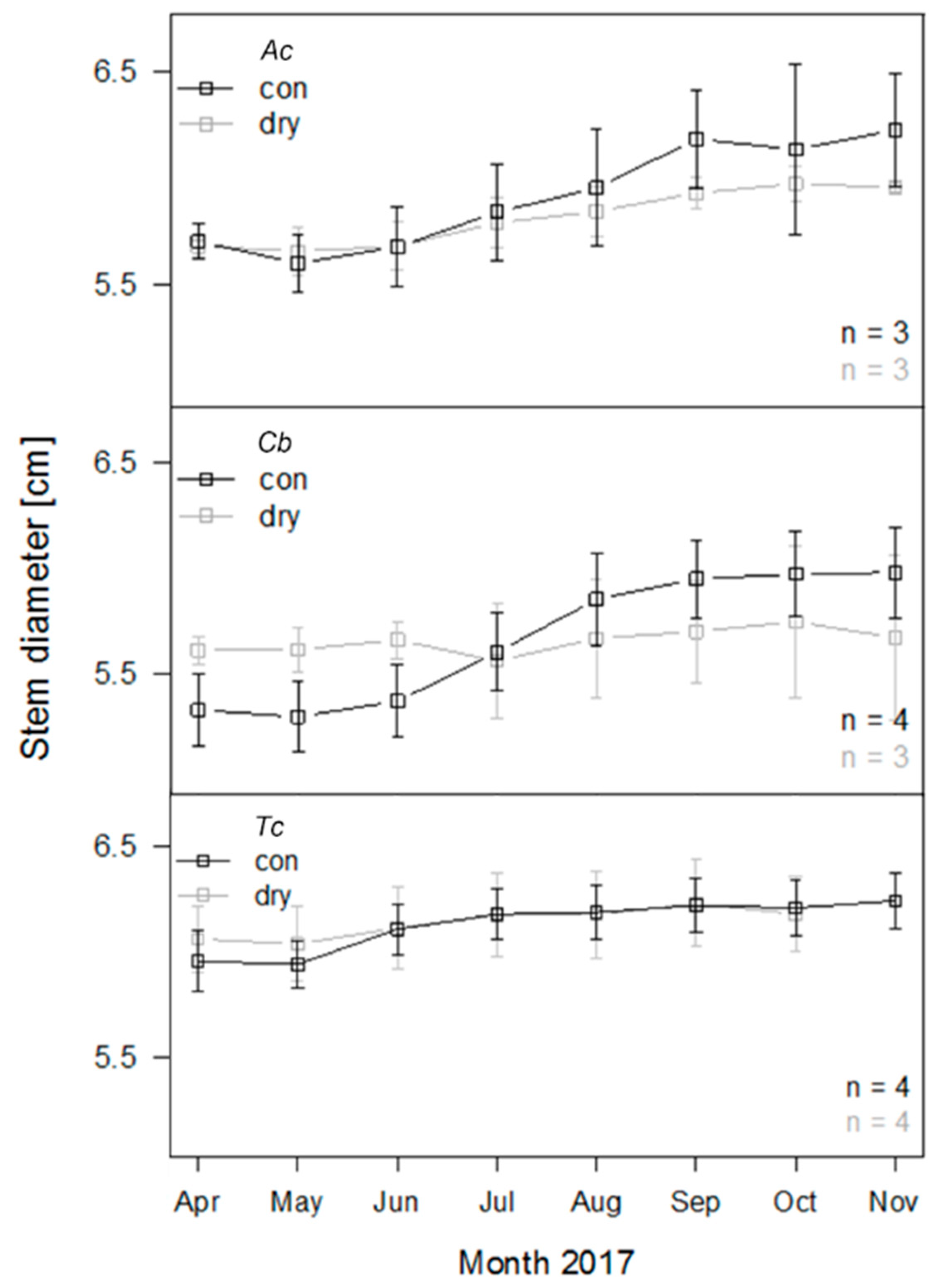
3.3. Stem Growth
3.4. Biomass Partitioning and Allocation
3.5. Species/Cultivar-Specific Adaptation and Changes of Tree Shapes with Drought
4. Discussion
4.1. Drought Resistance and Within-Plant Carbon Partitioning and Allocation
4.2. The Role of the Root System in Sustaining Drought
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Roloff, A.R. Stadt-und Straßenbäume der Zukunft—Welche Arten sind geeignet? Forstwiss Beitr Tharandt Beih 2013, 14, 173–187. [Google Scholar]
- Oleson, K.W.; Bonan, G.B.; Feddema, J. Effects of white roofs on urban temperature in a global climate model. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Sailor, D.J. A review of methods for estimating anthropogenic heat and moisture emissions in the urban environment. Int. J. Climatol. 2011, 31, 189–199. [Google Scholar] [CrossRef]
- Stone, B.; Vargo, J.; Habeeb, D. Managing climate change in cities: Will climate action plans work? Landsc. Urban Plan. 2012, 107, 263–271. [Google Scholar] [CrossRef]
- Moser, A.; Rahman, M.A.; Pretzsch, H.; Pauleit, S.; Rötzer, T. Inter- and intraannual growth patterns of urban small-leaved lime (Tilia cordata mill.) at two public squares with contrasting microclimatic conditions. Int. J. Biometeorol. 2017, 61, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Schütze, G.; Perkins, D.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; et al. Climate change accelerates growth of urban trees in metropolises worldwide. Sci. Rep. 2017, 7, 15403. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Microclimatic differences and their influence on transpirational cooling of Tilia cordata in two contrasting street canyons in Munich, Germany. Agric. For. Meteorol. 2017, 232, 443–456. [Google Scholar] [CrossRef]
- Nowak, D.J.; Crane, D.E.; Stevens, J.C. Air pollution removal by urban trees and shrubs in the United States. Urban For. Urban Green. 2006, 4, 115–123. [Google Scholar] [CrossRef]
- Armson, D. The Effect of Trees and Grass on the Thermal and Hydrological Performance of an Urban Area. Ph.D. Thesis, University of Manchester, Manchester, UK, 2012. [Google Scholar]
- Nowak, D.J.; Greenfield, E.J.; Hoehn, R.E.; Lagpoint, E. Carbon storage and sequestration by trees in urban and community areas of the United States. Environ. Pollut. 2013, 178, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Within canopy temperature differences and cooling ability of Tilia cordata trees grown in urban conditions. Build. Environ. 2017, 114, 118–128. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Stratópoulos, L.M.F.; Duthweiler, S.; Häberle, K.-H.; Pauleit, S. Effect of native habitat on the cooling ability of six nursery-grown tree species and cultivars for future roadside plantings. Urban For. Urban Green. 2018, 30, 37–45. [Google Scholar] [CrossRef]
- Stratópoulos, L.M.F.; Zhang, C.; Duthweiler, S.; Häberle, K.-H.; Rötzer, T.; Xu, C.; Pauleit, S. Tree species from two contrasting habitats for use in harsh urban environments respond differently to extreme drought. Int. J. Biometeorol. 2019, 63, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Field, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Stratopoulos, L.M.F.; Rötzer, T.; Xu, C.; Pretzsch, H. Development of fine-root biomass in response to drought stress of two contrasting urban tree species. under review.
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root:shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Kunz, J.; Räder, A.; Bauhus, J. Effects of drought and rewetting on growth and gas exchange of minor European broadleaved tree species. Forests 2016, 7, 239. [Google Scholar] [CrossRef]
- Sjöman, H.; Nielsen, A.B.; Pauleit, S.; Olsson, M. Habitat Studies Identifying Potential Trees for Urban Paved Environments: A Case Study from Qinling Mt., China. Arboric. Urban For. 2010, 36, 261–271. [Google Scholar]
- Sjöman, H.; Gunnarsson, A.; Pauleit, S.; von Bothmer, R. Selection Approach of Urban Trees for Inner-city Environments: Learning from Nature. Arboric. Urban For. 2012, 38, 194–204. [Google Scholar]
- Gillner, S.; Vogt, J.; Tharang, A.; Dettmann, S.; Roloff, A. Role of street trees in mitigating effects of heat and drought at highly sealed urban sites. Landsc. Urban Plan. 2015, 143, 33–42. [Google Scholar] [CrossRef]
- Dahlhausen, J.; Uhl, E.; Heym, M.; Biber, P.; Ventura, M.; Panzacchi, P.; Tonon, G.; Horváth, T.; Pretzsch, H. Stand density sensitive biomass functions for young oak trees at four different European sites. Trees 2017, 31, 1811–1826. [Google Scholar] [CrossRef]
- Rötzer, T.; Biber, P.; Moser, A.; Schäfer, C.; Pretzsch, H. Stem and root diameter growth of European beech and Norway spruce under extreme drought. For. Ecol. Manag. 2017, 406, 184–195. [Google Scholar] [CrossRef]
- DWD. 2018. Available online: https://www.dwd.de (accessed on 15 March 2018).
- BGR. Bodenkundliche Kartieranleitung; E. Schweizerbart’sche: Hannover, Germany, 2005. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and water logging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Zhang, C.; Stratopoulos, L.M.F.; Pretzsch, H.; Rotzer, T. How Do Tilia Cordata Greenspire Trees Cope with Drought Stress Regarding Their Biomass Allocation and Ecosystem Services? Forests 2019, 10, 676. [Google Scholar] [CrossRef]
- Biologische Bundesanstalt für Land- und Forstwirtschaft. Available online: http://www.agrometeo.ch/sites/default/files/u10/bbch-skala_deutsch.pdf (accessed on 18 August 2018).
- Miller, J.B. A formula for average foliage density. Aust. J. Bot. 1967, 15, 141–144. [Google Scholar] [CrossRef]
- Welles, J.M.; Norman, J.M. Instrument for indirect measurement of canopy architecture. Agron. J. 1991, 83, 818–825. [Google Scholar] [CrossRef]
- Hotaling, N.A.; Bharti, K.; Kriel, H.; Simon, C.G., Jr. DiameterJ: A validated open source nanofiber diameter measurement tool. Biomaterials 2015, 61, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014.
- ArcGIS, version 10.2.1; Esri: Redlands, CA, USA, 2014.
- Kiermeier, P. Die Lebensbereiche der Gehölze eingeteilt nach dem Kennziffernsystem; Grün ist Leben: Pinneberg, Germany, 1995. [Google Scholar]
- Roloff, A.R.; Bärtels, A. Flora der Gehölze—Bestimmung, Eigenschaften Verwendung; Ulmer Verlag: Stuttgart, Germany, 2008. [Google Scholar]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [Green Version]
- Reubens, B.; Poesen, J.; Danjon, F. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Gewin, V. Food: An underground revolution. Nat. News 2010, 466, 552–553. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.L.; Clément, C. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Reichwein, S. Baumwurzeln unter Verkehrsflächen. Untersuchungen zu Schäden an Verkehrsflächen durch Baumwurzeln und Ansätze zur Schadensbehebung und Schadensvermeidung. Ph.D. Thesis, University of Hannover, Hannover, Germany, 2002. [Google Scholar]
- Pretzsch, H.; Uhl, E.; Biber, P.; Schütze, G.; Coates, K.D. Change of allometry between coarse root and shoot of Lodgepole pine (Pinus contorta DOUGL. ex. LOUD) along a stress gradient in the sub-boreal forest zone of British Columbia. Scand. J. For. Res. 2012, 27, 532–544. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter [cm ± sd] | |||||||
|---|---|---|---|---|---|---|---|
| Code | Control n | Dry | Control | Dry | Shade Tolerance 1 | Drought Tolerance 1 | |
| Acer campestre | Ac | 3 | 3 | 5.60 ± 0.25 | 5.66 ± 0.09 | 3.18 ± 0.14 | 2.93 ± 0.32 |
| Carpinus betulus ‘Fastigiata’ | Cb | 4 | 3 | 5.30 ± 0.14 | 5.61 ± 0.08 | 3.97 ± 0.12 | 2.66 ± 0.16 |
| Tilia cordata ‘Greenspire’ | Tc | 4 | 4 | 5.90 ± 0.10 | 6.00 ± 0.16 | 4.18 ± 0.16 | 2.75 ± 0.15 |
| Dry Weight (g) | Group | n | Mean | ±SD | Min | Media | Max | Rtc | Drought Effect Species/Cultivar Effect | |
|---|---|---|---|---|---|---|---|---|---|---|
| Branch | Ac | con | 3 | 1121.33 | 177.46 | 920 | 1189 | 1255 | 0.70 | P < 0.001 *** P < 0.001 *** |
| dry | 3 | 785.00 | 89.50 | 696 | 784 | 875 | ||||
| Cb | con | 4 | 593.25 | 8.54 | 588 | 590 | 606 | 0.61 | ||
| dry | 3 | 365.33 | 60.34 | 309 | 358 | 429 | ||||
| Tc | con | 4 | 445.75 | 31.02 | 415 | 442 | 485 | 0.69 | ||
| dry | 4 | 308.50 | 49.10 | 243 | 317 | 358 | ||||
| Stem | Ac | con | 3 | 6162.33 | 1204.59 | 5015 | 6055 | 7417 | 0.92 | P = 0.0544 n.s. P < 0.001 *** |
| dry | 2 | 5652.50 | 127.99 | 5562 | 5652 | 5743 | ||||
| Cb | con | 4 | 5226.75 | 315.97 | 4862 | 5276 | 5492 | 0.96 | ||
| dry | 3 | 5007.67 | 389.79 | 4592 | 5066 | 5365 | ||||
| Tc | con | 4 | 4089.25 | 220.16 | 3817 | 4102 | 4336 | 0.83 | ||
| dry | 4 | 3407.00 | 322.40 | 2962 | 3480 | 3707 | ||||
| Roots | Ac | con | 3 | 5280.00 | 467.73 | 4938 | 5089 | 5813 | 0.73 | P < 0.001 *** P < 0.001 *** |
| dry | 3 | 3830.00 | 186.62 | 3628 | 3866 | 3996 | ||||
| Cb | con | 4 | 2440.25 | 219.50 | 2193 | 2422 | 2725 | 0.55 | ||
| dry | 3 | 1338.67 | 89.39 | 1271 | 1305 | 1440 | ||||
| Tc | con | 4 | 1869.00 | 63.59 | 1807 | 1863 | 1943 | 0.69 | ||
| dry | 4 | 1291.75 | 25.66 | 1266 | 1290 | 1321 | ||||
| Root:Shoot Ratio | Ac | con | 3 | 0.73 | 0.08 | 0.68 | 0.70 | 0.82 | 0.81 | P = 0.005 ** P < 0.001 *** |
| dry | 3 | 0.59 | 0.02 | 0.58 | 0.59 | 0.60 | ||||
| Cb | con | 4 | 0.42 | 0.02 | 0.40 | 0.41 | 0.45 | 0.60 | ||
| dry | 3 | 0.25 | 0.01 | 0.24 | 0.25 | 0.26 | ||||
| Tc | con | 4 | 0.41 | 0.03 | 0.39 | 0.40 | 0.45 | 0.85 | ||
| dry | 4 | 0.35 | 0.03 | 0.33 | 0.34 | 0.39 | ||||
| Above-Ground | ||||||||
| Code | Height [m ± sd] | Max. Canopy Radius [cm ± sd] | Canopy Volume [m3 ± sd] | LAI [m2 m−2 ± sd] | ||||
| Control | Dry | Control | Dry | Control | Dry | Control | Dry | |
| Ac | 5.90 ± 0.14 | 6.00 ±0.00 | 48.33 ± 8.06 | 50.00 ± 9.63 | 0.64 ± 0.25 | 0.67 ± 0.23 | 1.52 ± 0.11 | 1.81 ± 0.41 |
| Cb | 5.90 ± 0.25 | 5.53 ±0.17 | 28.50 ± 1.50 | 21.33 ± 0.94 | 0.36 ± 0.09 | 0.11 ± 0.55 | 3.01 ± 0.87 | 4.11 ± 1.49 |
| Tc | 5.13 ± 0.22 | 5.23 ±0.18 | 43.00 ± 17.45 | 52.50 ±11.37 | 0.65 ± 0.20 | 0.68 ± 0.22 | 2.53 ± 0.25 | 2.13 ± 0.24 |
| Below-Ground | ||||||||
| Code | Rooting Depth [cm ± sd] | Max. Rooting Width [cm ± sd] | Root Volume [m3 ± sd] | Porosity Root System [% ± sd] | ||||
| Control | Dry | Control | Dry | Control | Dry | Control | Dry | |
| Ac | 30.33 ± 7.85 | 29.67 ± 0.47 | 31.00 ± 2.75 | 29.92 ± 2.00 | 0.050 ± 0.019 | 0.056 ± 0.008 | 0.30 ± 0.03 | 0.30 ± 0.05 |
| Cb | 30.00 ± 0.00 | 30.00 ± 0.00 | 32.25 ± 0.98 | 29.00 ± 2.01 | 0.057 ± 0.005 | 0.036 ± 0.007 | 0.33 ± 0.05 | 0.34 ± 0.03 |
| Tc | 28.50 ± 1.50 | 29.00 ± 1.41 | 30.75 ± 1.75 | 28.67 ± 2.77 | 0.058 ± 0.000 | 0.047 ± 0.009 | 0.24 ± 0.01 | 0.38 ± 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stratópoulos, L.M.F.; Zhang, C.; Häberle, K.-H.; Pauleit, S.; Duthweiler, S.; Pretzsch, H.; Rötzer, T. Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars. Sustainability 2019, 11, 5117. https://doi.org/10.3390/su11185117
Stratópoulos LMF, Zhang C, Häberle K-H, Pauleit S, Duthweiler S, Pretzsch H, Rötzer T. Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars. Sustainability. 2019; 11(18):5117. https://doi.org/10.3390/su11185117
Chicago/Turabian StyleStratópoulos, Laura Myrtiá Faní, Chi Zhang, Karl-Heinz Häberle, Stephan Pauleit, Swantje Duthweiler, Hans Pretzsch, and Thomas Rötzer. 2019. "Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars" Sustainability 11, no. 18: 5117. https://doi.org/10.3390/su11185117
APA StyleStratópoulos, L. M. F., Zhang, C., Häberle, K.-H., Pauleit, S., Duthweiler, S., Pretzsch, H., & Rötzer, T. (2019). Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars. Sustainability, 11(18), 5117. https://doi.org/10.3390/su11185117