Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle



Abstract
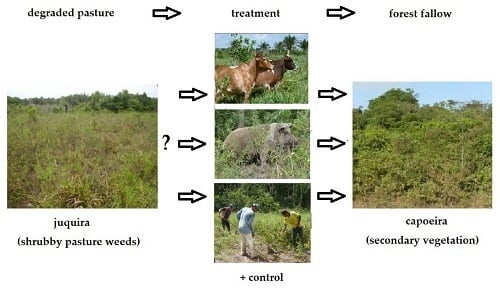
1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Design
2.2. Vegetation Sampling
2.3. Soil Compaction
- µ = overall mean,
- at = effect of treatment (i = trt1, trt2, trt3, trt4),
- bf = block effect (j = farm1, farm2, farm3),
- ci = effect of time (k = time1, time2, time3),
- abtf = interaction of treatment and block effect,
- acti = interaction of treatment and time effect,
- dtfi = residual deviation.
2.4. Economic Costs and Benefits
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Serrão, E.A.S.; Falesi, I.C.; da Veiga, J.B.; Teixeira Neto, J.F. Productivity of Cultivated Pastures on Low Fertility Soils of the Amazon Brazil. In Pasture Production in Acid Soils of the Tropics; Sánchez, P.A., Tergas, L.E., Eds.; Centro Internacional de Agricultura Tropical: Cali, Colombia, 1979; pp. 195–225. [Google Scholar]
- Serrão, E.A.S.; Nepstad, D.C. Pastures on Amazonian forestlands: A review of environmental and economic performance. In Interdisciplinary Research on the Conservation and Sustainable Use of the Amazonian Rainforest and Its Information Requirements; Lieberei, R., Reisdorff, C., Dantas Machado, A., Eds.; SBCS: Geesthacht, Germany; Stern-Werke: Hamburg, Germany, 1996; pp. 221–239. ISBN 3-00-000909-4. [Google Scholar]
- Dias-Filho, M.B. Degradação de pastagens: Processos, causas e estratégias de recuperação, 4th ed.; MBDF: Belém, Brazil, 2015. [Google Scholar]
- Buschbacher, R.; Uhl, C.; Serrão, E.A.S. Abandoned pastures in Eastern Amazonia. II. Nutrient stocks in the soil and vegetation. J. Ecol. 1988, 76, 682–699. [Google Scholar] [CrossRef]
- Müller, M.M.L.; Guimarães, M.F.; Desjardins, T.; Mitja, D. The relationship between pasture degradation and soil properties in the Brazilian Amazon: A case study. Agric. Ecosyst. Environ. 2004, 103, 279–288. [Google Scholar] [CrossRef]
- Davidson, E.A.; Reis de Carvalho, C.J.; Figueira, A.M.; Ishida, F.Y.; Ometto, J.P.H.B.; Nardoto, G.B.; Sabá, R.T.; Hayashi, S.N.; Leal, E.C.; Vieira, I.C.G.; et al. Recuperation of nitrogen cycling in Amazonian forests following agricultural abandonment. Nature 2007, 447, 995–998. [Google Scholar] [CrossRef]
- Uhl, C.; Buschbacher, R.; Serrão, E.A.S. Abandoned pastures in Eastern Amazonia. I. Patterns of plant succession. J. Ecol. 1988, 76, 663–681. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Pascarella, J.B.; Rivera, L.; Marcano-Vega, H. Forest Regeneration in a Chronosequence of Tropical Abandoned Pastures: Implications for Restoration Ecology. Rest. Ecol. 2000, 8, 328–338. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscape. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef]
- Bruel, B.O.; Marques, M.C.M.; Britez, R.M. Survival and Growth of Tree Species under Two Direct Seedling Planting Systems. Rest. Ecol. 2010, 18, 414–417. [Google Scholar] [CrossRef]
- Bertacchi, M.I.F.; Amazonas, N.T.; Brancalion, P.H.; Brondani, G.E.; Oliveira, A.; Pascoa, M.A.; Rodrigues, R.R. Establishment of tree seedlings in the understory of restoration plantations: Natural regeneration and enrichment plantings. Rest. Ecol. 2015, 24, 100–108. [Google Scholar] [CrossRef]
- Denich, M. Estudo da importância de uma vegetação secundária nova para o incremento da produtividade do sistema de produção na Amazônia oriental brasileira, 2nd ed.; GTZ, EMBRAPA/CPATU: Eschborn, Germany; Belém, Brazil, 1991. [Google Scholar]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014. [Google Scholar]
- Holl, K.D. Factors Limiting Tropical Rain Forest Regeneration in Abandoned Pasture: Seed Rain, Seed Germination, Microclimate, and Soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Rondon, M.A.; Fernandes, E.C.M.; Riha, S.J.; Wandelli, E. Carbon and nutrient accumulation in secondary forests regenerating on pastures in central Amazonia. Ecol. Appl. 2004, 14, 164–176. [Google Scholar] [CrossRef]
- De Rouw, A. The fallow period as a weed-break in shifting cultivation (tropical wet forests). Agric. Ecosyst. Environ. 1995, 54, 31–43. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Uhl, C.; Pereira, C.A.; Cardoso da Silva, J.M. A comparative study of tree establishment in abandoned pasture and mature forest of eastern Amazonia. Oikos 1996, 76, 25–39. [Google Scholar] [CrossRef]
- Uhl, C.; Nepstad, D.; da Silva, J.M.C.; Vieira, I. Restauração da floresta em pastagens degradadas. Ciência Hoje 1991, 13, 23–31. [Google Scholar]
- Aide, T.M.; Cavelier, J. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta Colombia. Rest. Ecol. 1994, 2, 219–229. [Google Scholar] [CrossRef]
- Da Silva, J.M.C.; Uhl, C.; Murray, G. Plant Succession, Landscape Management, and the Ecology of Frugivorous Birds in Abandoned Amazonian Pastures. Conserv. Biol. 1996, 10, 491–543. [Google Scholar] [CrossRef]
- Muscarella, R.; Fleming, T.H. The role of frugivorous bats in tropical forest succession. Biol. Rev. 2007, 82, 573–590. [Google Scholar] [CrossRef]
- Reid, J.L.; Holl, K.D.; Zahawi, R.A. Seed dispersal limitations shift over time in tropical forest restoration. Ecol. Appl. 2015, 25, 1072–1082. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Herrera, L.; Rosario, M.; Serrano, M. Forest recovery in abandoned tropical. pastures in Puerto Rico. For. Ecol. Man. 1995, 77, 77–86. [Google Scholar] [CrossRef]
- Engel, V.L.; Parrotta, J.A. An evaluation of direct seeding for reforestation of degraded lands in central São Paulo state Brazil. For. Ecol. Man. 2001, 152, 169–181. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Augspurger, C.K. Early plant succession in abandoned pastures in Ecuador. Biotropica 1999, 31, 540–552. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Rosario, M.; Marcano, H. Forest recovery in abandoned cattle pastures along an elevational gradient in northeastern Puerto Rico. Biotropica 1996, 28, 537–548. [Google Scholar] [CrossRef]
- Uhl, C. Factors controlling succession following slash-and-burn agriculture in Amazonia. J. Ecol. 1987, 75, 377–407. [Google Scholar] [CrossRef]
- Lindell, C.A.; Reid, J.L.; Cole, R.J. Planting design effects on avian seed dispersers in a tropical forest restoration experiment. Rest. Ecol. 2013, 21, 515–522. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Uhl, C.; Serrão, E.A.S. Surmounting barriers to forest regeneration in abandoned, highly degraded pastures: A case study from Paragominas, Pará, Brazil. In Alternatives to Deforestation: Steps toward Sustainable Use of the Amazon Rain Forest; Anderson, A.B., Ed.; Columbia University Press: New York, NY, USA, 1990; pp. 215–229. [Google Scholar]
- Nepstad, D.C.; Uhl, C.; Serrão, E.A.S. Restoration of degraded Amazonian landscape: Forest recovery and agricultural restoration. Ambio 1991, 20, 248–255. [Google Scholar]
- Zahawi, R.A.; Holl, K.D. Comparing the performance of tree stakes and seedlings to restore abandoned tropical pastures. Rest. Ecol. 2009, 17, 854–864. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Holl, K.D.; Cole, R.J.; Reid, J.L. Testing applied nucleation as a strategy to facilitate tropical forest recovery. J. Appl. Ecol. 2013, 50, 88–96. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; Camarão, A.P.; Schultze-Kraft, R.; Rodrigues Filho, J.A.; King, J.M. Integrating cattle into the slash-and-burn cycle on smallholdings in the Eastern Amazon: Grass-capoeira pasture versus grass-legume pasture. Agric. Ecosyst. Environ. 2006, 117, 266–276. [Google Scholar] [CrossRef]
- Hohnwald, S.; Trautwein, J.; Camarão, A.P.; Wollny, C.B.A. Relative palatability and growth performance of capoeira species as supplementary forages in the NE-Amazon. Agric. Ecosyst. Environ. 2016, 218, 107–115. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Holl, K.D.; Peneireiro, F.M. Agro-Successional Restoration as a Strategy to Facilitate Tropical Forest Recovery. Rest. Ecol. 2009, 17, 451–459. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; Schultze-Kraft, R.; Rodrigues-Filho, J.A.; Camarão, A.P. Experiences with legumes as part of a ley pasture in a low input farming system of North-Eastern Pará, Brazil. Pasturas Trop. 2005, 27, 2–12. [Google Scholar]
- Baar, R.; dos Reis Cordeiro, M.; Denich, M.; Fölster, H. Floristic inventory of secondary vegetation in agricultural systems of East-Amazonia. Biodivers. Conserv. 2004, 13, 501–528. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Lima, R.A.F.; Gandolfi, S.; Nave, A. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol. Conserv. 2009, 142, 1242–1251. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. Tropical secondary forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Denich, M.; Vielhauer, K.; Kato, M.D.A.; Block, A.; Kato, O.R.; de Sá Abreu, T.D.; Lücke, W.; Vlek, P.L.G. Mechanized land preparation in forest-based fallow systems: The experience from Eastern Amazonia. Agric. Syst. 2004, 61, 91–106. [Google Scholar] [CrossRef]
- Kato, M.S.A.; Kato, O.R.; Denich, M.; Vlek, P.L.G. Fire-free alternatives to slash and-burn for shifting cultivation in the eastern Amazon region: The role of fertilizers. Field Crops Res. 1999, 62, 225–237. [Google Scholar] [CrossRef]
- Sommer, R.; Vlek, P.L.G.; de Sá, T.D.A.; Vielhauer, K.; Rodrigues Coelho, R.F.; Fölster, H. Nutrient balance of shifting cultivation by burning or mulching in the Eastern Amazon—Evidence for subsoil nutrient accumulation. Nutr. Cycl. Agroecosyst. 2004, 68, 257–271. [Google Scholar] [CrossRef]
- Denich, M.; Vlek, P.L.G.; de Sá, T.D.A.; Vielhauer, K.; Lücke, W. A concept for the development of fire-free fallow management in the Eastern Amazon, Brazil. Agric. Ecosyst. Environ. 2005, 110, 43–58. [Google Scholar] [CrossRef]
- Hohnwald, S. Bird Records from the Rural Landscape of Igarapé-Açu Municipality, Northeastern Pará. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 2009, 4, 119–131. [Google Scholar]
- Hohnwald, S.; Acioli de Abréu, E.; Krummel, T.; da Veiga, J.B.; Wollny, C.B.A.; Calandrini de Azevedo, C.M.B.; Gerold, G. Degraded Pasture Expansion and Woody Enrichment Strategies for Pasture Fertility Preservation in the Bragantina Region, North-Eastern Amazon. Erdkunde 2010, 64, 17–31. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; King, J.M.; Camarão, A.P.; Rodrigues Filho, J.A.; Zeppenfeld, T. Intensive Cattle Browsing Did Not Prevent Fallow Recuperation on Smallholder Grass-Capoeira Pastures in the NE-Amazon. Agric. Syst. 2015, 89, 813–828. [Google Scholar] [CrossRef]
- Staver, C. The Role of Weeds in the Productivity of Amazonian Bush Fallow Agriculture. Exp. Agric. 1991, 27, 287–304. [Google Scholar] [CrossRef]
- Dias-Filho, M.B. Germination and emergence of Stachytarpheta cayennensis and Ipomoea asarifolia. Planta Daninha 1996, 14, 118–126. [Google Scholar] [CrossRef]
- Krummel, T.; Hohnwald, S.; Gerold, G. Biologisch degradierte Rinderweiden und ihre agrarökologischen Regradationsmöglichkeiten für Kleinbauern in Nordost-Amazonien. Innsbr. Geogr. Stud. 2011, 38, 43–56. [Google Scholar]
- Vieira, I.C.G.; Uhl, C.; Nepstad, D.C. The role of shrub Cordia multispicata Cham. as a ‘succession facilitator’ in an abandoned pasture, Paragominas, Amazônia. Vegetatio 1994, 115, 91–99. [Google Scholar]
- Holl, K.D.; Loik, M.E.; Lin, E.H.V.; Samuels, I.A. Tropical Montane Forest Restoration in Costa Rica: Overcoming Barriers to Dispersal and Establishment. Rest. Ecol. 2000, 8, 339–349. [Google Scholar] [CrossRef]
- Reid, J.L.; Holl, K.D. Arrival ≠ Survival. Rest. Ecol. 2013, 21, 153–155. [Google Scholar] [CrossRef]
- Peña-Domene, M.; Martínez-Garza, C.; Palmas-Pérez, S.; Rivas-Alonso, E.; Howe, H.F. Roles of Birds and Bats in Early Tropical-Forest Restoration. PLoS ONE 2014, 9, e104656. [Google Scholar] [CrossRef]
- Posada, J.M.; Aide, T.M.; Cavalier, J. Cattle and Weedy Shrubs as Restoration Tools of Tropical Montane Rainforest. Rest. Ecol. 2000, 8, 370–379. [Google Scholar] [CrossRef]
- Elledge, A.E.; McAlpine, C.A.; Murray, P.J.; Gordon, I.J. Modelling habitat preferences of feral pigs for rooting in lowland rainforest. Biol. Invasion 2013, 15, 1523–1535. [Google Scholar] [CrossRef]
- Nogueira-Filho, S.L.G.; Nogueira, S.S.C.; Fragoso, J.M.V. Ecological impacts of feral pigs in the Hawaiian Islands. Biodivers. Conserv. 2009, 18, 3677–3683. [Google Scholar] [CrossRef]
- Spatz, G.; Mueller-Dombois, D. Succession patterns after pig diggings in grassland communities on Mauna Loa, Hawaii. Phytocoenologia 1975, 3, 346–373. [Google Scholar]
- Censo Agropecuário 2006 and Produção da Pecuária Municipal-2014 of Igarapé-Açu; IBGE: Brasilia, Brazil, 2017. Available online: http://cod.ibge.gov.br/2358p (accessed on 15 December 2017).
- Pachêco, N.A.; Bastos, T.X. Boletim Agrometeorológico de 2007 para Igarape-Açu, PA. In Documentos 354; Embrapa Amazônia Oriental: Belém, Brazil, 2009. [Google Scholar]
- Pachêco, N.A.; Bastos, T.X. Boletim Agrometeorológico de 2008 para Igarape-Açu, PA. In Documentos 372; Embrapa Amazônia Oriental: Belém, Brazil, 2011. [Google Scholar]
- Baena, A.R.C.; Falesi, I.C.; Dutra, S. Características Físio-Químicas do Solo em Diferentes Agroecossistemas na Região Bragantina do Nordeste Paraense; Embrapa Amazônia Oriental: Belém, Brazil, 1998. [Google Scholar]
- R: A Language and Environment for Statistical Computing; R Core Team R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 17 January 2018).
- Oanda cooperation; Currency converter. 2019. Available online: https://www.oanda.com/currency/converter/ (accessed on 15 February 2019).
- Kotanen, P.M. Responses of vegetation to a changing regime of disturbance: Effects of feral pigs in a Californian coastal prairie. Ecography 1995, 18, 190–198. [Google Scholar] [CrossRef]
- Aplet, G.H.; Anderson, S.J.; Stone, C.P. Association between feral pig disturbance and the composition of some alien plant assemblages in Hawaii Volcanoes National Park. Plant Ecol. 1991, 95, 55–62. [Google Scholar] [CrossRef]
- Sharrow, S.H. Soil compaction by grazing livestock in silvopastures as evidenced by changes in soil physical properties. Agric. Syst. 2007, 71, 215–223. [Google Scholar] [CrossRef]
- Veloso, H.P. As communidades e as estações botânicas de Teresópolis, Estado do Rio de Janeiro. Boletim do Museu Nacional. Botânica. 1945, 3, 1–95. [Google Scholar]
- Albuquerque, N.I.; Guimarães, D.A.; Tavares Dias, H.L.; le Pendu, Y.; Reis Kahwage, P.; Rossetto Garcia, A. Intensive production system of collared peccary (Pecari tajacu) in Brazilian Amazon. Adv. Anim. Biosci. 2010, 1, 480–481. [Google Scholar] [CrossRef]
- Garnero, A.D.V.; Marcondes, C.R.; Albuquerque, N.I.; Araújo, R.O.; de Pendu, Y.; Guimarães, D.A. Growth curve of female collared peccaries (Pecari tajacu) raised in captivity in the Brazilian Amazon Region. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2013, 65, 961–966. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin-Young, M. Revision of the Common International Classification for Ecosystem Services (CICES V5. 1): A Policy Brief. One Ecosyst. 2018, 3, e27108. [Google Scholar] [CrossRef]
- Díaz, S.; Pascual, U.; Stenseke, M.; Martín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.; Baste, I.A.; Brauman, K.A. Assessing nature’s contributions to people. Science 2018, 359, 270–272. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Plant (Sub) Family | Life-Form |
|---|---|---|
| a) Spontaneous Legume Species | ||
| Zornia latifolia Sm. | Papilionoideae | herb |
| Mimosa pudica L. | Mimosoideae | herb |
| Stylosanthes gracilis Kunth | Papilionoideae | shrub |
| Senna chrysocarpa (Desv.) H.S. Irwin & Barneby | Caesalpinioideae | shrub |
| Mimosa quadrivalvis L. | Mimosoideae | herb |
| Machaerium madeirense Pittier | Papilionoideae | liana |
| Machaerium froesii Rudd | Papilionoideae | liana |
| Desmodium barbatum (L.) Benth. | Papilionoideae | herb |
| Desmodium canum (J.F. Gmel.) Schinz & Thell. | Papilionoideae | herb |
| Bauhinia guianensis Aubl. | Caesalpinioideae | liana |
| b) Capoeira | ||
| Vismia guianensis (Aubl.) Choisy | Clusiaceae | tree |
| Lacistema pubescens Mart. | Lacistemataceae | tree |
| Myrcia sylvatica (G. Mey.) DC. | Myrtaceae | shrub |
| Myrcia deflexa (Poir.) DC. | Myrtaceae | shrub |
| Myrcia bracteata (Rich.) DC. | Myrtaceae | shrub |
| Banara guianensis Aubl. | Connaraceae | tree |
| Lecythis lurida (Miers) S.A. Mori | Lecythidaceae | tree |
| Abarema cochleata (Willd.) Barneby & J.W. Grimes | Mimosoideae | tree |
| c) Juquira | ||
| Borreria verticillata (L.) G. Mey. | Rubiaceae | shrub |
| Myrciaria tenella (DC.) O. Berg | Myrtaceae | shrub |
| Borreria latifolia (Aubl.) K. Schum. | Rubiaceae | shrub |
| Stachytarpheta cayennensis (Rich.) M. Vahl | Verbenaceae | shrub |
| Paspalum maritimum Trin. | Poaceae | herb |
| Paspalum conjugatum P.J. Bergius | Poaceae | herb |
| Panicum pilosum Sw. | Poaceae | herb |
| Scleria pterota C. Presl | Cyperaceae | herb |
| Rourea ligulata Baker | Connaraceae | liana |
| Rolandra argentea Rottb. | Asteraceae | shrub |
| Rollinia exsucca (DC. ex Dunal) A. DC. | Annonaceae | tree |
| Hyptis atrorubens Poit. | Lamiaceae | herb |
| Imperata brasiliensis Trin. | Poaceae | herb |
| Emilia sonchifolia (L.) DC. | Asteraceae | herb |
| Ipomoea asarifolia (Desr.) Roem. & Schult. | Verbenaceae | liana |
| Borreria suaveolens G. Mey. | Rubiaceae | shrub |
| Andropogon bicornis L. | Poaceae | herb |
| Andropogon leucostachyus Kunth | Poaceae | herb |
| Capoeira Trees time1 [#] | Capoeira Trees time2 [#] | Mean Tree Heights time1 [m] | Mean Tree Heights time2 [m] | Capoeira Saplings time3 [#] | |
|---|---|---|---|---|---|
| n = 1440 | n = 1129 | n = 960 | |||
| trt1 | 5.84 (0.47) ab1 | 6.13 (0.54) a1 | 1.0 a1 (0.6) | 1.1 a1 (0.4) | 2.56 (2.8) a |
| trt2 | 7.07 (0.45) a1 | 6.42 (0.41) a1 | 1.3 b1 (0.7) | 1.3 b1 (0.6) | 2.64 (2.8) a |
| trt3 | 5.52 (0.40) b1 | 4.55 (0.45) b1 | 0.9 a1 (0.7) | 1.3 b2 (0.4) | 2.84 (3.0) a |
| trt4 | 6.52 (0.41) ab1 | 3.77 (0.29) b2 | 1.1 a1 (0.6) | 1.5 c2 (0.5) | 1.33 (1.8) b |
| Legume Cover time1 [%] | Legume Cover time2 [%] | |
|---|---|---|
| n = 1360 | ||
| trt1 | 6.9 a1 (11.0) | 14.4 a2 (19.4) |
| trt2 | 15.6 b1 (20.6) | 15.4 a1 (18.5) |
| trt3 | 20.3 bc1 (25.1) | 29.2 b2 (27.0) |
| trt4 | 24.0 c1 (28.4) | 14.4 a2 (19.7) |
| time1 | ||||
| Depths | trt1 | trt2 | trt3 | trt4 |
| 2.5 cm | 1.88 (0.13) a | 1.95 (0.13) a | 1.71 (0.14) a | 2.05 (0.14) a |
| 5 cm | 2.07 (0.13) ab | 2.21 (0.13) ab | 1.82 (0.15) a | 2.25 (0.12) b |
| 10 cm | 2.21 (0.13) ab | 2.37 (0.11) a | 1.90 (0.16) b | 2.43 (0.10) a |
| 15 cm | 2.18 (0.13) ab | 2.32 (0.16) ab | 1.90 (0.16) a | 2.40 (0.11) b |
| 20 cm | 2.12 (0.14) a | 2.24 (0.18) a | 1.68 (0.12) b | 1.98 (0.14) ab |
| 40 cm | 1.56 (0.09) a | 1.34 (0.17) a | 1.38 (0.11) a | 1.40 (0.05) a |
| time2 | ||||
| Depths | trt1 | trt2 | trt3 | trt4 |
| 2.5 cm | 2.95 (0.18) b | 3.06 (0.19) bc | 2.63 (0.19) bc | 2.44 (0.15) ac |
| 5 cm | 3.04 (0.15) cd | 3.51 (0.19) bcd | 3.43 (0.20) c | 2.84 (0.17) bd |
| 10 cm | 3.06 (0.14) c | 3.77 (0.15) c | 4.20 (0.18) c | 4.01 (0.19) c |
| 15 cm | 3.26 (0.15) c | 3.86 (0.17) c | 4.05 (0.25) c | 4.67 (0.14) c |
| 20 cm | 3.34 (0.20) c | 3.61 (0.19) c | 3.45 (0.27) c | 3.42 (0.29) c |
| 40 cm | 2.51 (0.16) b | 3.26 (0.22) ab | 4,29 (0.19) b | 4.21 (0.21) b |
| Number of Pigs | Mean Pig Weights [kg/pig] | Total Pig Weight before [kg/0.5 ha] | Total Pig Weight after [kg/0.5 ha] | Total Weight Gain [kg; %] | Economic Gain from Pigs [US$/ha] | |
|---|---|---|---|---|---|---|
| farm1 | 14 | 20.1 | 281 (20.0) | 572 (25.0) | 291; +104% | 727.50 |
| farm2 | 11 | 24.0 | 264 (17.3) | 467 (18.7) | 203; +77% | 507.50 |
| farm3 | 10 | 29.8 | 298 (15.5) | 624 (21.6) | 326; +109% | 815.00 |
| mean | 11.7 | 24.6 | 281.0 | 554.3 | 273.3 | 683.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hohnwald, S.; Kato, O.R.; Walentowski, H. Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle. Sustainability 2019, 11, 1729. https://doi.org/10.3390/su11061729
Hohnwald S, Kato OR, Walentowski H. Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle. Sustainability. 2019; 11(6):1729. https://doi.org/10.3390/su11061729
Chicago/Turabian StyleHohnwald, Stefan, Osvaldo Ryohei Kato, and Helge Walentowski. 2019. "Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle" Sustainability 11, no. 6: 1729. https://doi.org/10.3390/su11061729
APA StyleHohnwald, S., Kato, O. R., & Walentowski, H. (2019). Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle. Sustainability, 11(6), 1729. https://doi.org/10.3390/su11061729