Agronomic Evaluation of Bread Wheat Varieties from Participatory Breeding: A Combination of Performance and Robustness

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the PPB Populations and Commercial Varieties Studied
2.2. Agronomic Evaluation
2.3. Statistical Analyses
3. Results
3.1. Effects and Interactions in the ANOVA
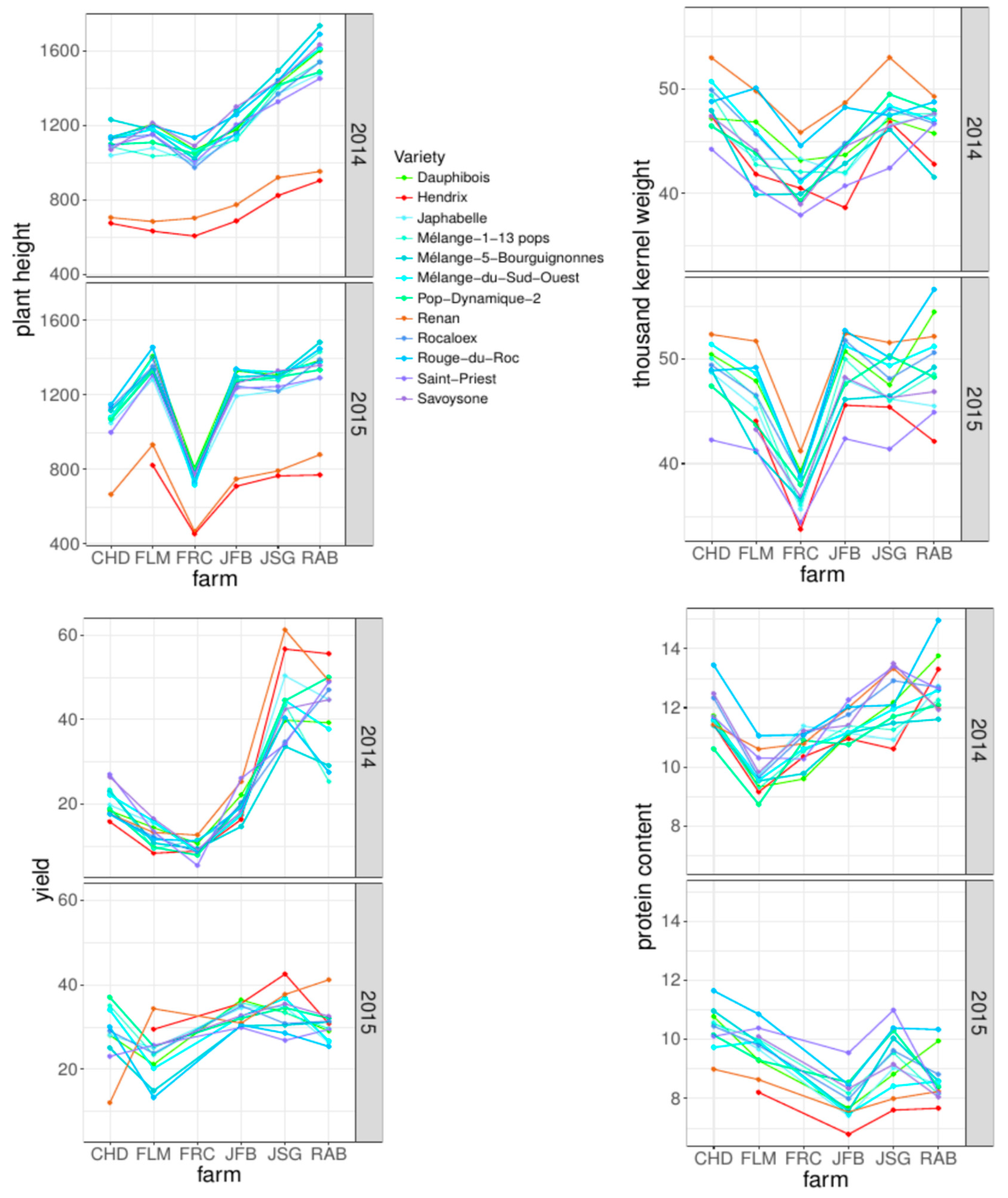
3.2. Comparison of PPB Populations and Commercial Varieties over Farms and Years
3.3. Overall Variety Means
3.4. Contribution of the Varieties to the Interaction
4. Discussion
4.1. The Whole Range of Outcomes of PPB Programs
4.2. Agronomic Performance and Robustness of PPB Varieties Compared to the Commercial Varieties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Atlin, G.N.; Cooper, M.; Bjornstad, A. A Comparison of Formal and Participatory Breeding Approaches Using Selection Theory. Euphytica 2001, 122, 463–475. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Erskine, W.; Hamblin, J.; Grando, S. Genotype by Environment Interaction and International Breeding Programs. Exp. Agric. 1994, 30, 177–187. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S.; Tutwiler, R.; Baha, J.; Martini, A.M.; Salahieh, H.; Goodchild, A.; Michael, M. A Methodological Study on Participatory Barley Breeding—I. Selection Phase. Euphytica 2000, 111, 91–104. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Participatory plant breeding: Who did it, who does it and where? Exp. Agric. 2019, 1–11. [Google Scholar] [CrossRef]
- Witcombe, J.R.; Yadavendra, J.P. How much evidence is needed before client-oriented breeding (COB) is institutionalised? Evidence from rice and maize in India. Field Crops Res. 2014, 167, 143–152. [Google Scholar] [CrossRef]
- Rivière, P.; Dawson, J.C.; Goldringer, I.; David, O. Hierarchical Bayesian Modeling for Flexible Experiments in Decentralized Participatory Plant Breeding. Crop. Sci. 2015, 55, 1053–1067. [Google Scholar] [CrossRef]
- Van Frank, G.; Goldringer, I.; Riviere, P.; David, O. Influence of experimental design on decentralized, on-farm evaluation of populations: A simulation study. Euphytica 2019, 215, 126. [Google Scholar] [CrossRef]
- Ceccarelli, S. Efficiency of Plant Breeding. Crop Sci. 2015, 55, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Goldringer, I.; Rivière, P. Methods and Tools for Decentralized on Farm Breeding. Booklet #3. Diversifood Project. Available online: http://www.diversifood.eu/wp-content/uploads/2018/12/booklet3_decentralized_on_farm_breeding_BAT_web_A4_2.pdf (accessed on 1 October 2019).
- Rivière, P.; van Frank, G.; Rouger, B.; David, O.; Munoz, F.; Vindras, C.; Thomas, M.; Goldringer, I. PPBstats: An R Package to Perform Analyses Found within PPB Programs. Available online: https://github.com/priviere/PPBstats_web_site (accessed on 1 October 2019).
- Diversifood Innovation Factsheeth #2. Varieties and Populations for On-Farm Participatory Plant Breeding. Available online: http://www.diversifood.eu/wp-content/uploads/2018/06/Diversifood_innovation_factsheet2_VarietiesPopulations.pdf (accessed on 1 October 2019).
- Dawson, J.C.; Rivière, P.; Berthellot, J.F.; Mercier, F.; De Kochko, P.; Galic, N.; Pin, S.; Serpolay, E.; Thomas, M.; Giuliano, S.; et al. Collaborative Plant Breeding for Organic Agricultural Systems in Developed Countries. Sustainability 2011, 3, 1206–1223. [Google Scholar] [CrossRef] [Green Version]
- Rivière, P.; Pin, S.; Galic, N.; De Oliveira, Y.; David, O.; Dawson, J.; Wanner, A.; Heckmann, R.; Obbellianne, S.; Ronot, B.; et al. Mise en place d’une méthodologie de sélection participative sur le blé en France. Innov. Agron. 2013, 32, 427–441. [Google Scholar]
- Gauch, H.G. Statistical Analysis of Yield Trials by AMMI and GGE. Crop Sci. 2006, 46, 1488–1500. [Google Scholar] [CrossRef]
- Wricke, G. Uber Eine Methode zur Erfassung der Okologischen Streubreite in Feldversuchen. Zeitschrift Fur Pflanzenzuchtung/J. Plant Breed. 1962, 47, 92–96. [Google Scholar]
- Becker, H.C.; Leon, J. Stability Analysis in Plant-Breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Decentralized-Participatory Plant Breeding: An Example of Demand Driven Research. Euphytica 2007, 155, 349–360. [Google Scholar] [CrossRef]
- Elise, D.; Bonneuil, C. Des Semences en partage. Construction sociale et identitaire d’un collectif paysan autour de pratiques semancières alternatives. Techniques & Culture. Rev. Semest. d’Anthropol. Tech. 2011, 15, 202–221. [Google Scholar] [CrossRef]
- Kloppenburg, J. Impeding dispossession, enabling repossession: Biological open source and the recovery of seed sovereignty. J. Agrar. Chang. 2010, 10, 367–388. [Google Scholar] [CrossRef]
- Chable, V.; Nuijten, E.; Costanzo, A.; Goldringer, I.; Bocci, R.; Oehen, B.; Rey, F.; Fasoula, D.; Feher, J.; Keskitalo, M.; et al. Embedding cultivated diversity in society for agro-ecological transition. Sustainability. submitted.
- Rivière, P.; Goldringer, I.; Berthellot, J.-F.; Galic, N.; Pin, S.; De Kochko, P.; Dawson, J.C. Response to farmer mass selection in early generation progeny of bread wheat landrace crosses. Renew. Agric. Food Syst. 2015, 30, 190–201. [Google Scholar] [CrossRef]
- Börner, A.; Schumann, E.; Fürste, A.; Cöster, H.; Leithold, B.; Röder, M.; Weber, W. Weber Mapping of Quantitative Trait Loci Determining Agronomic Important Characters in Hexaploid Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2002, 105, 921–936. [Google Scholar] [CrossRef]
- Raggi, L.; Ciancaleoni, S.; Torricelli, R.; Terzi, V.; Ceccarelli, S.; Negri, V. Evolutionary Breeding for Sustainable Agriculture: Selection and Multi-Environmental Evaluation of Barley Populations and Lines. Field Crop. Res. 2017, 204, 76–88. [Google Scholar] [CrossRef]
- Borg, J.; Kiær, L.; Lecarpentier, C.; Goldringer, I.; Gauffreteau, A.; Saint-Jean, S.; Barot, S.; Enjalbert, J. Unfolding the potential of wheat cultivar mixtures: A meta-analysis perspective and identification of knowledge gaps. Field Crops Res. 2018, 221, 298–313. [Google Scholar] [CrossRef]
- Klaedtke, S.M.; Caproni, L.; Klauck, J.; de la Grandville, P.; Dutartre, M.; Stassart, P.M.; Chable, V.; Negri, V.; Raggi, L. Short-Term Local Adaptation of Historical Common Bean (Phaseolus vulgaris L.) Varieties and Implications for In Situ Management of Bean Diversity. Int. J. Mol. Sci. 2017, 18, 493. [Google Scholar] [CrossRef] [PubMed]
- Döring, T.F.; Annicchiarico, P.; Clarke, S.; Haigh, Z.; Jones, H.E.; Pearce, H.; Snape, J.; Zhan, J.; Wolfe, M.S. Comparative analysis of performance and stability among composite cross populations, variety mixtures and pure lines of winter wheat in organic and conventional cropping systems. Field Crops Res. 2015, 183, 235–245. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S.; Bailey, E.; Amri, A.; El-Felah, M.; Nassif, F.; Rezgui, S.; Yahyaoui, A. Farmer Participation in Barley Breeding in Syria, Morocco and Tunisia. Euphytica 2001, 122, 521–536. [Google Scholar] [CrossRef]
- Van Etten, J.; de Sousa, K.; Aguilar, A.; Barrios, M.; Coto, A.; Dell’Acqua, M.; Fadda, C.; Gebrehawaryat, Y.; van de Gevel, J.; Gupta, A.; et al. Crop Variety Management for Climate Adaptation Supported by Citizen Science. Proc. Natl. Acad. Sci. USA 2019, 116, 4194–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vindras-Fouillet, C.; Ranke, O.; Anglade, J.P.; Taupier-Letage, B.; Chable, V.; Goldringer, I. Sensory analyses and nutritional qualities of hand-made breads with organic grown wheat bread populations. Food Nutr. Sci. 2014, 5, 1860–1874. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety Name | Origin (Farmer, Location) | Development Process |
|---|---|---|
| Saint-Priest | FLM, Maine-et-Loire | Derived from a Swedish variety registered in 1942 (Progress) |
| Rouge du Roc | JFB, Lot-et-Garonne | Mass selection within the landrace Rouge de Bordeaux in 2001 |
| Pop Dynamique 2 | FLM, Maine-et-Loire | Mix of three landraces and two more recent varieties |
| Mélange-5 Bourguignon | BER, Côte d’Or | Mix of 11 local landraces |
| Mélange du Sud-Ouest | JFB, Lot-et-Garonne | Mix of around 20 landraces from the south-west of France |
| Savoysone | RAB, Haute-Savoie | Population derived from a cross between two local landraces |
| Rocaloex | RAB, Haute-Savoie | Mix of 11 populations derived from crosses |
| Mélange1 13 Pops | BER, Côte d’Or | Mix of 13 populations derived from crosses |
| Dauphibois | CHD, Isère | Mix of around 25 local landraces, populations derived from crosses, and more recent varieties |
| Japhabelle | JFB, Lot-et-Garonne | Mix of around 25 populations derived from crosses and selected on farm |
| Renan | INRA | Pure line commercial variety registered in 1989 |
| Hendrix | INRA | Pure line commercial variety for organic agriculture registered in 2013 |
| Farms | Meteorological Stations | Minimum Temperature in Winter | Maximum Temperature in June-July | Cumulative Rainfall from February to June |
|---|---|---|---|---|
| CHD | Grenoble | −1.2 °C | +26.9 °C | 380 mm |
| FLM | Beaucouzé | +2.5 °C | +25.3 °C | 264 mm |
| FRC | Nîmes | +2.7 °C | +31.0 °C | 252 mm |
| JFB | Agen | +2.1 °C | +27.6 °C | 304 mm |
| JSC | Clermont-Ferrand | −0.1 °C | +26.5 °C | 251 mm |
| RAB | Chambéry | −1.4 °C | +27.4 °C | 482 mm |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldringer, I.; van Frank, G.; Bouvier d’Yvoire, C.; Forst, E.; Galic, N.; Garnault, M.; Locqueville, J.; Pin, S.; Bailly, J.; Baltassat, R.; et al. Agronomic Evaluation of Bread Wheat Varieties from Participatory Breeding: A Combination of Performance and Robustness. Sustainability 2020, 12, 128. https://doi.org/10.3390/su12010128
Goldringer I, van Frank G, Bouvier d’Yvoire C, Forst E, Galic N, Garnault M, Locqueville J, Pin S, Bailly J, Baltassat R, et al. Agronomic Evaluation of Bread Wheat Varieties from Participatory Breeding: A Combination of Performance and Robustness. Sustainability. 2020; 12(1):128. https://doi.org/10.3390/su12010128
Chicago/Turabian StyleGoldringer, Isabelle, Gaëlle van Frank, Caroline Bouvier d’Yvoire, Emma Forst, Nathalie Galic, Maxime Garnault, Jonathan Locqueville, Sophie Pin, Julien Bailly, Raphael Baltassat, and et al. 2020. "Agronomic Evaluation of Bread Wheat Varieties from Participatory Breeding: A Combination of Performance and Robustness" Sustainability 12, no. 1: 128. https://doi.org/10.3390/su12010128
APA StyleGoldringer, I., van Frank, G., Bouvier d’Yvoire, C., Forst, E., Galic, N., Garnault, M., Locqueville, J., Pin, S., Bailly, J., Baltassat, R., Berthellot, J.-F., Caizergues, F., Dalmasso, C., de Kochko, P., Gascuel, J.-S., Hyacinthe, A., Lacanette, J., Mercier, F., Montaz, H., ... Rivière, P. (2020). Agronomic Evaluation of Bread Wheat Varieties from Participatory Breeding: A Combination of Performance and Robustness. Sustainability, 12(1), 128. https://doi.org/10.3390/su12010128