The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem


Abstract
:1. Introduction
2. Materials and Methods
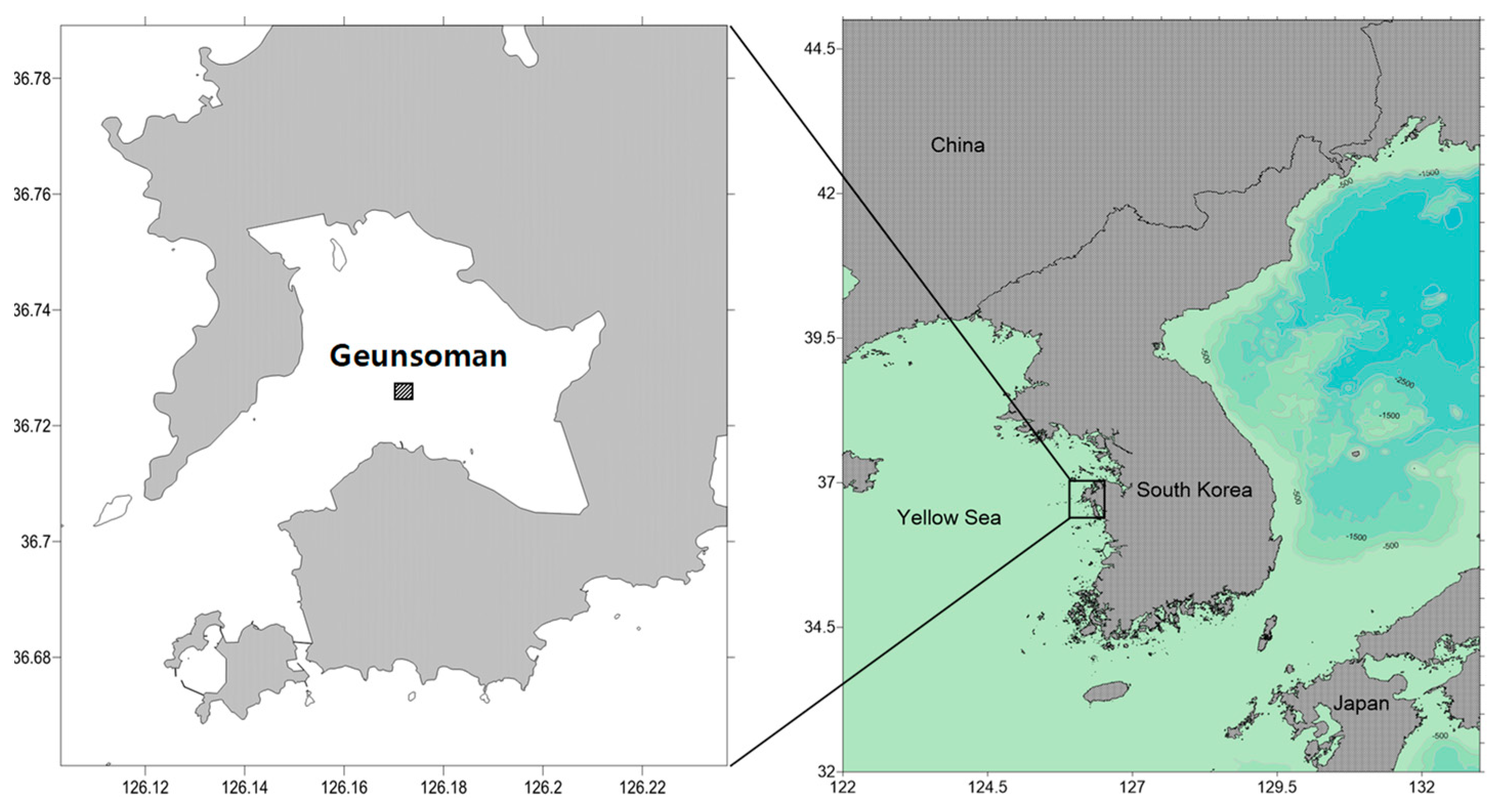
2.1. Study Area and Sampling Routine
2.2. Sediment Analysis
2.3. Statistical Analyses
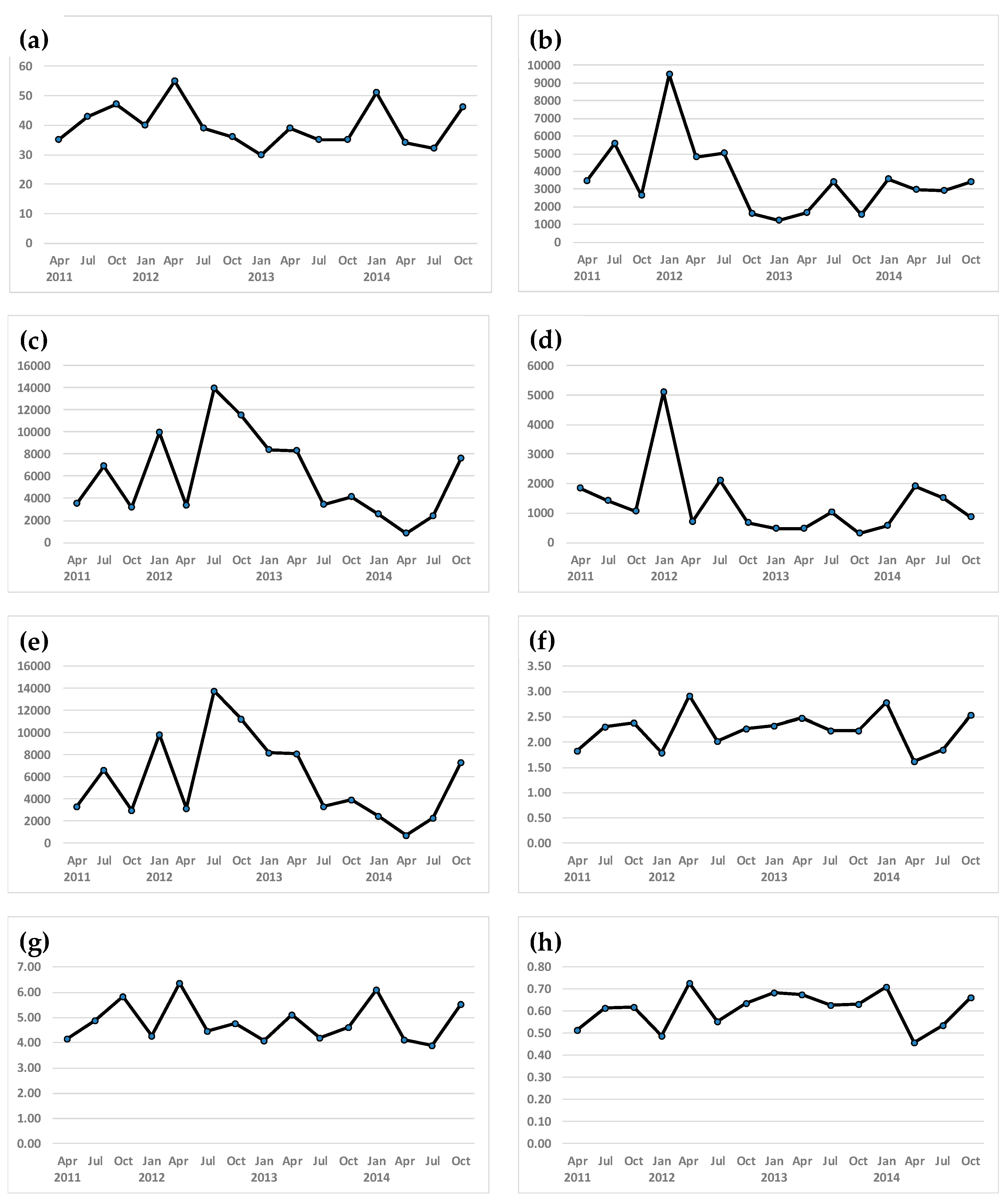
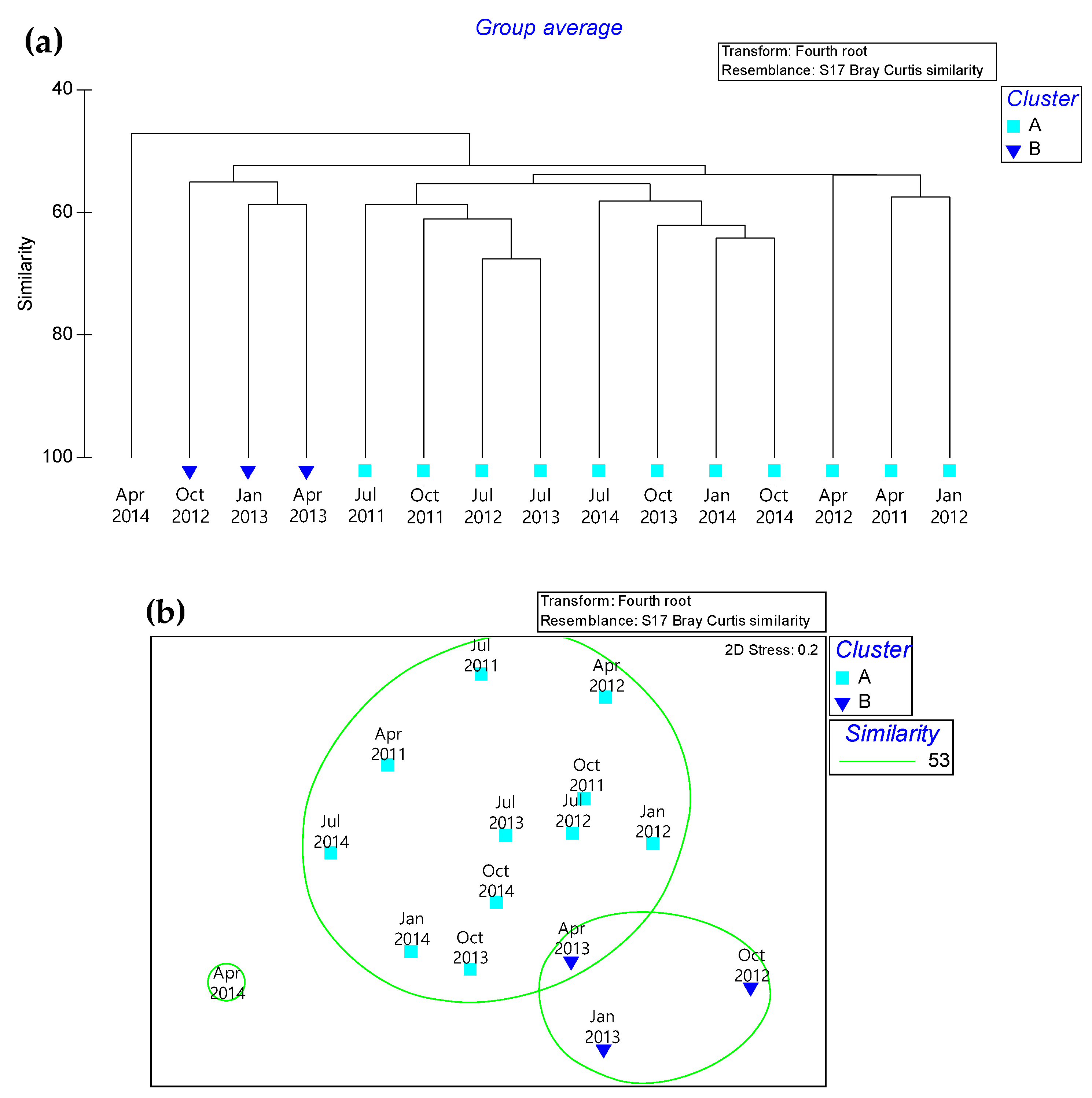
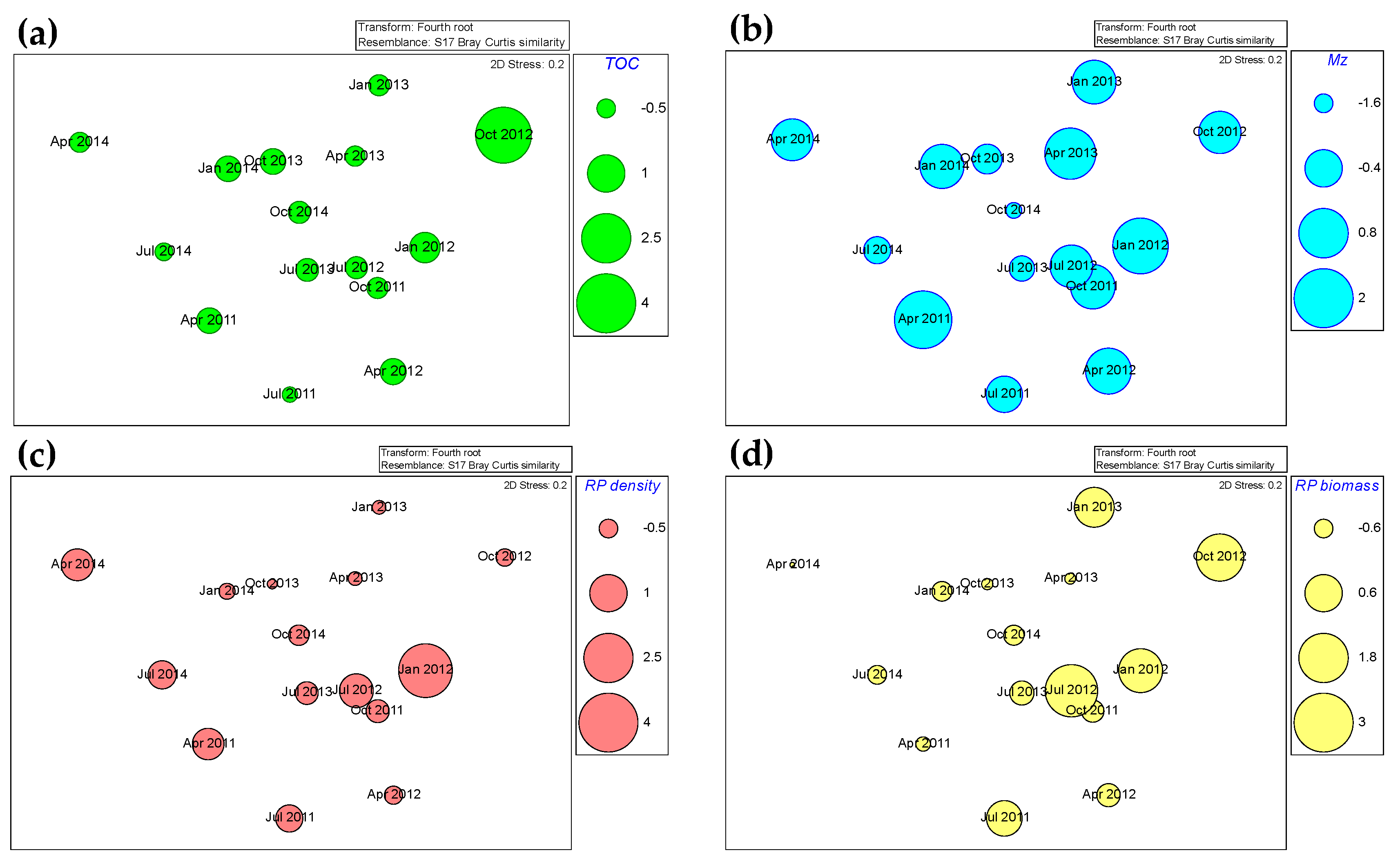
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Kim, S.L.; Kwon, S.H.; Lee, H.G.; Yu, O.H. Erratum to: Effects of environmental and biological conditions on the recruitment and growth of the Manila clam Ruditapes philippinarum on the west coast of Korea. Ocean Sci. J. 2017, 52, 91–101. [Google Scholar] [CrossRef]
- Oug, E.; Fleddum, A.; Rygg, B.; Olsgard, F. Biological traits analyses in the study of pollution gradients and ecologicalfunctioning of marine soft bottom species assemblages in a fjord ecosystem. J. Exp. Mar. Bio. Eco. 2012, 432, 90–105. [Google Scholar]
- Nordhaus, I.; Wolff, M. Feeding ecology of the mangrove crab Ucides cordatus (Ocypodidae): Food choice, food quality and assimilation efficiency. Mar. Biol. 2007, 151, 1665–1681. [Google Scholar] [CrossRef]
- Lee, S.Y. Mangrove macrobenthos: Assemblages, services, and linkages. J. Sea Res. 2008, 59, 16–29. [Google Scholar] [CrossRef]
- Bamber, R.N.; Spencer, J.F. The benthos of a coastal power station thermal discharge canal. J. Mar. Biol. Assoc. 1984, 64, 603–623. [Google Scholar] [CrossRef]
- Kristensen, E. Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. J. Sea Res. 2008, 59, 30–43. [Google Scholar] [CrossRef]
- Cairns, J.; Pratt, J.R. A history of biological monitoring using benthic macroinvertebrates. In Freshwater Biomonitoring and Benthic Macroinvertebrates; Rosenberg, D.M., Resh, V.H., Eds.; Chapman & Hall: London, UK, 1993; pp. 10–27. [Google Scholar]
- Alexander, C.R.; de Master, D.J.; Nittrouer, C.A. Sediment accumulation in a modern epicontinental-shelf setting: The Yellow Sea. Mar. Geol. 1991, 98, 51–72. [Google Scholar] [CrossRef]
- Clark, G.F.; Johnston, E.L. Temporal change in the diversity-invasibility relationship in the presence of a disturbance regime. Ecol. Lett. 2011, 14, 52–57. [Google Scholar] [CrossRef]
- Ferguson, N.; White, C.R.; Marshall, D.J. Competition in benthic marine invertebrates: The unrecognized role of exploitative competition for oxygen. Ecology 2013, 94, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Braeckman, U.; Van Colen, C.; Soetaert, K.; Vincx, M.; Vanaverbeke, J. Contrasting macrobenthic activities differentially affect nematode density and diversity in a shallow subtidal marine sediment. Mar. Ecol. Prog. Ser. 2011, 422, 170–191. [Google Scholar] [CrossRef] [Green Version]
- Schratzberger, M.; Warwick, R.M. Differential effects of various types of disturbances on the structure of nematode assemblages: An experimental approach. Mar. Ecol. Prog. Ser. 1999, 181, 227–236. [Google Scholar] [CrossRef]
- Marshall, D.J.; Keough, M.J. Does interspecific competition affect offspring provisioning? Ecology 2009, 90, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mckinney, F.K. Competitive interactions between related clades: Evolutionary implications of overgrowth interactions between encrustin cyclostome and cheilostome bryozoans. Mar. Biol. 1992, 114, 645–652. [Google Scholar] [CrossRef]
- Yang, S.Y.; Jung, H.S.; Lim, D.I.; Li, C.X. A review on the provenance discrimination of sediments in the Yellow Sea. Ear. Sci. Rev. 2003, 63, 93–120. [Google Scholar] [CrossRef]
- Yang, S.; Youn, J.S. Geochemical compositions and provenance discrimination of the central south Yellow Sea sediments. Mar. Geol. 2007, 243, 229–241. [Google Scholar] [CrossRef]
- Won, S.; Jung, H.; Sang, J.; Lee, T.K.; Shin, K.S. Planktonic bivalve larvae identification and quanti fi cation in Gomso Bay, South Korea, using next-generation sequencing analysis and microscopic observations. Aquaculture 2018, 490, 297–302. [Google Scholar]
- Flassch, J.P.; Leborgne, Y. Introduction in Europe, from 1972 to 1980, of the Japanese Manila clam (Tapes philippinarum) and the effects on aquacultur eproduction and natural settlement. Mar.Sci. 1992, 194, 92–96. [Google Scholar]
- Anderson, G.J. Comments on the settlement of manila clam spats (Tapes philippinarum) at Filucy Bay. J Shellfi. Res. 1982, 2, 115. [Google Scholar]
- Lim, H.S.; Choi, J.W. Macrobenthic community at the subtidal area aroud Taebudo in Kyeonggi Bay, Korea. J. Kor. Soc. Fish Res. 1998, 31, 453–462. [Google Scholar]
- Kim, S.H.; Jeoung, H.D.; Park, K.I. Comparison of diseases and immune parameters in the imported and domestic juvenile Manila clams, Ruditapes philippinarum. Kor. J. Mal. 2019, 35, 93–102. [Google Scholar]
- Song, J.H.; Kim, C.H.; Park, S.W.; Yu, J.; Jo, E.J. Seasonality of the Biological Activity Factors of the hard clam Meretrix lusoria in the Western Coast of Korea. J. Aquac. 2008, 21, 111–122. [Google Scholar]
- Nam, K.W.; Jeung, H.D.; Song, J.H.; Park, K.H.; Choi, K.S.; Park, K.I. High parasite burden increases the surfacing and mortality of the manila clam (Ruditapes philippinarum) in intertidal sandy mudflats on the west coast of Korea during hot summer. Parasit Vectors 2018, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Forrest, B.M.; Keeley, N.B.; Hopkins, G.A.; Webb, S.C.; Clement, D.M. Bivalve aquaculture in estuaries: Review and synthesis of oyster cultivation effects. Aquaculture 2009, 298, 1–15. [Google Scholar] [CrossRef]
- Filgueira, R.; Guyondet, T.; Comeau, L.A.; Grant, J. Physiological indices as indicators of ecosystem status in shellfish aquaculture sites. Ecol. Indic. 2014, 39, 134–143. [Google Scholar] [CrossRef]
- Frau, D.; Molina, F.R.; Mayora, G. Feeding selectivity of the invasive mussel Limnoperna fortunei (Dunker, 1857) on a natural phytoplankton assemblage: What really matters? Limnology 2015, 17, 1–11. [Google Scholar] [CrossRef]
- Wang, L.; Fan, Y.; Yan, C.; Gao, C.; Xu, Z.; Liu, X. Assessing benthic ecological impacts of bottom aquaculture using macrofaunal assemblages. Mar. Pollut. Bull. 2017, 114, 258–268. [Google Scholar] [CrossRef]
- Pearson, T.H.; Rosenberg, R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Ocean Mar. Ecol. 1978, 16, 229–311. [Google Scholar]
- Diaz, R.J.; Rosenberg, R. Marine benthic hypoxia: A review of its ecological effects and the behavioral responses of benthic macrofauna. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 245–303. [Google Scholar]
- Drummond, L.; Mulcahy, M.; Culloty, S. The reproductive biology of the manila clam, Ruditape sphilippinaurm, form the North-west of Ireland. J. Aqua. 2006, 245, 326–340. [Google Scholar] [CrossRef]
- Park, K.J.; Yoon, S.P.; Song, J.H.; Han, H.S. Improvement of Manila Clam (Ruditapes philippinarum) Habitat Condition by Adding Crushed Oyster (Crassostrea gigas) Shells to the Substratum. Kor. J. Mal. 2011, 27, 291–297. [Google Scholar]
- Griffiths, C.L.; Griffiths, R.J. Bivalvia. In Animal Energetics; Pandian, T.J., Vernberg, F.J., Eds.; Academic Press: New York, NY, USA, 1987; pp. 1–88. [Google Scholar]
- Zhang, G.; Yan, X. A new three-phase culture method for Manila clam, Ruditapes philippinarum, farming in northern China. Auaculture 2006, 258, 452–461. [Google Scholar] [CrossRef]
- Hoffman, K.H. Metabolic and enzyme adaptation to temperature and pressure. In The Mollusca; Hochachka, P.W., Ed.; Academic Press: New York, NY, USA, 1983; pp. 220–256. [Google Scholar]
- Shin, Y.K.; Kim, Y.; Chung, E.Y.; Hur, S.B. Temperature and salinity tolerance of the manila clam, Ruditapes philippinarum. J. Kor. Soc. Fish Res. 2000, 33, 213–218. [Google Scholar]
- Baud, J.P.; Bacher, C. Use of saline ground water for intensive rearing of Ruditapes philippinarum juveniles in anursery. Aquaculture 1990, 88, 157–178. [Google Scholar] [CrossRef]
- Delgado, M.; Pérez-Camacho, A. A study of gonadal development in Ruditapes decussatus(L.) (Mollusca: Bivalvia), using image analysis: Influence of food ration and energy balance. J. Shellfi. Res. 2003, 22, 435–441. [Google Scholar]
- Dang, C.; de Montaudouin, X.; Gam, M.; Paroissin, C.; Bru, N.; Caill-Milly, N. The manila clam population in Arcachon Bay (SW France): Can it be kept sustainable? J. Sea Res. 2010, 63, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Wilding, T.A. Changes in benthos associated with mussel (Mytilus edulis L.) farmson the west-coast of Scotland. PLoS ONE 2013, 8, e68313. [Google Scholar] [CrossRef]
- Lacoste, E.; Gueguen, Y.; Moullac, G.L.; Koua, M.S.; Gaertner-Mazouni, N. Influence of farmed pearl oysters and associated biofouling communities on nutrient regeneration in lagoons of French Polynesia. Aquac. Environ. Interact. 2014, 5, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Folk, R.L.; Ward, W.C. Brazos River Bar: A Study in the Significance of Grain Size Parameters. J. Sed. Res. 1957, 27, 3–26. [Google Scholar] [CrossRef]
- Walters, K.; Coen, L.D. A comparison of statistical approaches to analyzing community convergence between natural and constructed oyster reefs. J. Exp. Mar. Biol. Ecol. 2006, 330, 81–95. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual/Tutorial. PRIMER-E; Plymouth Marine Laboratory: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Zhang, J.; Xiao, N.; Zhang, S.; Xu, F.; Zhang, S. A comparative study on the macrobenthic community over a half century in the Yellow Sea, China. J. Oceanogr. 2016, 72, 189–205. [Google Scholar] [CrossRef]
- Piló, D.; Carvalho, A.N.; Pereira, F.; Coelho, H.E.; Gaspar, M.B. Evaluation of macrobenthic community responses to dredging through a multimetric approach: Effective or apparent recovery? Ecol. Indic. 2019, 9, 656–668. [Google Scholar] [CrossRef]
- Coates, D.A.; van Hoey, G.; Colson, L.; Vincx, M.; Vanaverbeke, J. Rapid macrobenthic recovery after dredging activities in an offshore wind farm in the Belgian part of the North Sea. Hydrobiologia 2015, 756, 3–18. [Google Scholar] [CrossRef]
- Somerfield, P.J.; Arvanitidis, C.; Vanden Berghe, E.; van Avesaath, P.; Hummel, H.; Heip, C.H.R. Assessing evidence for random assembly of marine benthic communities from regional species pools. Mar. Ecol. Prog. Ser. 2009, 382, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, S.K.J.; Denisenko, S.G.; Renaud, P.E.; Emblow, C.S.; Ambrose, W.G.; Ellingsen, I.H.; Skardhamar, J. Benthic macrofauna and productivity regimes in the Barents Sea-Ecological implications in a changing Arctic. J. Sea Res. 2009, 61, 222–233. [Google Scholar] [CrossRef]
- Silberberger, M.J.; Renaud, P.E.; Buhl-Mortensen, L.; Ellingsen, I.H.; Reiss, H. Spatial patterns in sub-Arctic benthos: Multiscale analysis reveals structural differences between community components. Ecol. Monogr. 2019, 89, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Hyun, S.M.; Chang, M. The Summer Benthic Environmental Conditions Assessed by the Functional Groups of Macrobenthic Fauna in Gwangyang Bay, Southern Coast of Korea. Kor. J. Environ. Biol 2003, 21, 101–113. [Google Scholar]
- Dobbs, F.C.; Scholly, T.A. Sediment processing and selective feeding by Pectinaria koreni (Polychaeta: Pectinariidae). Mar. Ecol. 1986, 29, 165–176. [Google Scholar] [CrossRef]
- Borja, A.; Muxika, I.; Franco, J. The application of a Marine Biotic Index to different impact sources affecting soft-bottom benthic communities along European coasts. Mar. Pollut. Bull. 2003, 46, 835–845. [Google Scholar] [CrossRef]
- Lee, H.J.; Chu, Y.S.; Park, Y.A. Sedimentary processes of fine-grained material and the effect of seawall construction in the Daeho macrotidal flat–nearshore area, northern west coast of Korea. Mar. Geol. 1999, 157, 171–184. [Google Scholar] [CrossRef]
- Silina, A.V. Population characteristics of the bivalve Ruditapes philippinarum from Cheju Island coasts, Korea. Korean J. Malacol. 2010, 26, 227–234. [Google Scholar]
- Icho, S.; Osima, Y. Test Book of Clam Culture; Sugiyama Book. Co.: Okayama, Japan, 1931; pp. 1–188. [Google Scholar]
- Choi, D.L.; Kwon, J.N.; Park, S.W. Infection and rapid detection of Perkinsus sp. in cultured babyneck clam, Ruditapes philippinarum from western coast of Korea. J. Fish Pathol. 1998, 11, 69–76. [Google Scholar]
- Ribeiro, L.F.; Eça, G.F.; Barros, F.; Hatje, V. Impacts of shrimp farming cultivation cycles on macrobenthic assemblages and chemistry of sediments. Environ. Poll. 2016, 211, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J. A Comparative Study on the Growth, Spawning and Recruitment of the Manila Clam, Ruditapes philippinarum According to Their Environmental Factors in Seonjaedo, Incheon, Korea. Ph.D. Thesis, Inha University, Incheon, Korea, 2005. [Google Scholar]
- Albayrak, S.; Balkis, H.; Zenetos, A.; Kurun, A.; Kubanc, C. Ecological quality status of coastal benthic ecosystems in the Sea of Marmara. Mar. Pollut Bull. 2006, 52, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Olapoju, O.A.; Edokpayi, C.A. Response of Macrobenthic Invertebrate to Organic Carbon and Particle Size in Lagos Lagoon, Nigeria. J. Fish Aquat. Sci. 2018, 2, 1–13. [Google Scholar] [CrossRef]
- Yu, O.H.; Lee, H.G.; Lee, J.H. The influence of environmental variables on macrobenthic communities after dike construction in Asan Bay, Korea. Korean J. Environ. Biol. 2011, 29, 326–344. [Google Scholar]
- Spiridonov, V.A.; Kolyuchkina, G.A.; Belyaev, N.A.; Basin, A.B.; Kozlovsky, V.V. Contemporary condition of macrozoobenthos in ultrashallow zone of Taman Bay, Sea of Azov. Oceanology 2016, 56, 248–258. [Google Scholar] [CrossRef]
- Domínguez Castanedo, N.; Hernández Alcántara, P.; Solís-Weiss, V.; Granados Barba, A. Distribution of polychaete feeding guilds in sedimentary environments of the Campeche Bank, Southern Gulf of Mexico. Helgol. Mar. Res. 2012, 66, 469–478. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of Worms Emended: An Update of Polychaete Feeding Guilds. Ann. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Antoniadou, C.; Voultsiadou, E.; Chintiroglou, C. Benthic colonization and succession on temperate sublittoral rocky cliffs. J. Exp. Mar. Biol. Ecol. 2010, 382, 145–153. [Google Scholar] [CrossRef]
- Jung, Y.H. Ecological Studies and Spatial Patterns of Macrobenthos on Tidal Flat around Anmyou-do. Ph.D. Thesis, Soonchunhyang University, Asan, South Korea, 2006. [Google Scholar]
- Loreau, M. Biodiversity and ecosystem functioning: Recent theoretical advances. Oikos 2000, 91, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Chin, P. Quantitative importance and species composition of polychaetes in the benthic community of the Yellow Sea. Kor. Soc. Fish Aquat. Sci. 1989, 22, 189–195. [Google Scholar]
- Díaz-Jaramillo, M.; Miglioranza, K.S.B.; Carriquiriborde, P.; Marino, D.; Pegoraro, C.N.; Valenzuela, G.; Barra, R. Sublethal effects in Perinereis gualpensis (Polychaeta: Nereididae) exposed to mercury-pyrene sediment mixture observed in a multipolluted estuary. Ecotoxicology 2017, 26, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.P.; Jung, R.H.; Kim, Y.J.; Kim, S.G.; Choi, M.K.; Lee, W.C.; Oh, H.T.; Hong, S.J. Macrobenthic Community Structure Along the Environmental Gradients of Ulsan Bay, Korea. J. Kor. Soc. Ocean 2009, 14, 102–117. [Google Scholar]
- Musale, A.S.; Desai, D.V. Distribution and abundance of macrobenthic polychaetes along the South Indian coast. Environ. Monit. Assess 2011, 178, 423–436. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Month | Type | Mz (ø) | TOC (%) |
|---|---|---|---|---|
| 2011 | Apr | sM | 6.46 | 0.58 |
| 2011 | Jul | gmS | 3.39 | 0.31 |
| 2011 | Oct | (g)sM | 4.40 | 0.44 |
| 2012 | Jan | (g)sM | 6.15 | 0.75 |
| 2012 | Apr | (g)mS | 4.59 | 0.59 |
| 2012 | Jul | gmS | 4.16 | 0.47 |
| 2012 | Oct | gM | 4.11 | 2.15 |
| 2013 | Jan | (g)mS | 4.32 | 0.44 |
| 2013 | Apr | (g)mS | 5.40 | 0.42 |
| 2013 | Jul | gmS | 2.34 | 0.49 |
| 2013 | Oct | gmS | 2.72 | 0.58 |
| 2014 | Jan | gM | 4.34 | 0.58 |
| 2014 | Apr | zS | 4.06 | 0.42 |
| 2014 | Jul | gmS | 2.49 | 0.36 |
| 2014 | Oct | S | 1.72 | 0.47 |
| Species | Group A Average Density | Group B Average Density | Average Dissimilarity | Contribution % | Cumulative % | Groups A & B Dissimilarity |
|---|---|---|---|---|---|---|
| Ampharete arctica (P) | 3.05 | 0 | 1.86 | 3.89 | 3.89 | 49.85 |
| Musculus senhousia (M) | 3.15 | 0.7 | 1.54 | 3.23 | 7.12 | |
| Crangon affinis (C) | 2.12 | 0.95 | 1.21 | 2.54 | 9.67 | |
| Chone teres (P) | 1.85 | 0 | 1.1 | 2.3 | 11.97 | |
| Ilyoplax pingi (C) | 1.87 | 1.2 | 0.85 | 1.77 | 13.74 | |
| Amaeana occidentalis (P) | 1.32 | 0 | 0.78 | 1.64 | 15.38 | |
| Diastylis paratricincta (C) | 1.8 | 1.47 | 0.78 | 1.63 | 17.01 | |
| Mediomastus californiensis (P) | 1.51 | 0.5 | 0.77 | 1.61 | 18.63 | |
| Ruditapes philippinarum (M) | 5.93 | 4.79 | 0.76 | 1.59 | 20.22 | |
| Philine argentata (M) | 1.16 | 0.81 | 0.75 | 1.56 | 21.78 | |
| Palaemon serrifer (C) | 0 | 1.19 | 0.73 | 1.53 | 23.31 | |
| Grandidierella japonica (C) | 2.51 | 1.34 | 0.72 | 1.52 | 24.83 | |
| Reticunassa festiva (M) | 1.05 | 1.9 | 0.65 | 1.37 | 26.2 | |
| Anoides oxycephala (P) | 1.31 | 0.5 | 0.65 | 1.37 | 27.56 | |
| Eteone longa (P) | 1.42 | 1.2 | 0.61 | 1.28 | 28.84 |
| Number of Variables | Correlation (%) | Best Variables |
|---|---|---|
| 3 | 0.320 | TOC, RP density, RP biomass |
| 2 | 0.299 | TOC, RP biomass |
| 2 | 0.289 | TOC, RP density |
| 1 | 0.270 | TOC |
| 3 | 0.251 | Mz, TOC, RP biomass |
| 1 | 0.249 | RP biomass |
| 4 | 0.247 | Mz, TOC, BP density, RP biomass |
| 2 | 0.244 | BP density, RP biomass |
| 2 | 0.208 | Mz, TOC |
| 3 | 0.199 | Mz, TOC, BP density |
| TOC | Species | Density | Biomass | H’ | RPSu den | RPSu bio | HFDe | CEDt | AADe | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mz | 0.320 | 0.271 | 0.211 | 0.146 | 0.043 | 0.118 | 0.100 | 0.004 | 0.182 | −0.460 |
| TOC | 0.271 | 0.144 | 0.286 | 0.050 | −0.115 | 0.279 | 0.273 | 0.363 | −0.374 | |
| Species | 0.515 * | 0.004 | 0.648 ** | −0.020 | 0.004 | 0.608 * | 0.705 ** | 0.133 | ||
| Density | 0.021 | −0.125 | 0.657 ** | 0.018 | 0.493 | 0.714 ** | 0.453 | |||
| Biomass | −0.004 | −0.054 | 0.996 *** | 0.339 | 0.300 | −0.357 | ||||
| H’ | −0.679 ** | 0.007 | 0.232 | 0.289 | −0.072 | |||||
| RPSu den | −0.068 | −0.004 | 0.179 | 0.422 | ||||||
| RPSu bio | 0.346 | 0.318 | −0.327 | |||||||
| HFDe | 0.779 *** | −0.027 | ||||||||
| CEDt | 0.164 | |||||||||
| AADe |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.L.; Lee, H.G.; Kang, S.M.; Yu, O.H. The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem. Sustainability 2020, 12, 4205. https://doi.org/10.3390/su12104205
Kim SL, Lee HG, Kang SM, Yu OH. The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem. Sustainability. 2020; 12(10):4205. https://doi.org/10.3390/su12104205
Chicago/Turabian StyleKim, Sang Lyeol, Hyung Gon Lee, Su Min Kang, and Ok Hwan Yu. 2020. "The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem" Sustainability 12, no. 10: 4205. https://doi.org/10.3390/su12104205
APA StyleKim, S. L., Lee, H. G., Kang, S. M., & Yu, O. H. (2020). The Influence of Manila Clam (Ruditapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem. Sustainability, 12(10), 4205. https://doi.org/10.3390/su12104205