1. Introduction
Microbes, as decomposers of ecosystems, participate in the carbon, nitrogen, and phosphorus cycle and energy flow in various types of ecosystems around the world, which are important drivers of biogeochemical cycles [
1,
2]. As an important part of the soil system, soil microorganisms play an important role in the element balance of terrestrial ecosystems by regulating the decomposition of soil organic matter, plant litter, and the availability of plant nutrients [
3]. The grassland ecosystem is one of the most common types of terrestrial ecosystems, and grazing is an important usage of land [
4,
5]. The composition of soil microbial communities in grassland ecosystems can reflect not only the changes in soil ecological environment under the influences of grazing and human activities but also the changes in the composition and function of the grassland ecosystem. The composition and function of the grassland ecosystem are often affected by human activities and climate change [
6,
7,
8,
9]. Therefore, studying the change mechanism of soil microbial communities under experimental warming, grazing, and their interaction can provide a theoretical basis for grazing management of grassland ecosystems and the construction of a multi-functional ecological security barrier [
10,
11].
To date, through controlled experiments, previous studies have been carried out on the effects of climate warming on the composition of soil microbial communities in ecosystems such as
Stipa grassland, tundra, and forest. The effects of experimental warming on the soil microbial community composition varied between being inconsistent, showing no changes [
12,
13] and showing significant changes [
3,
14]. Previous studies examining the effects of grazing on the composition of soil microbial communities were carried out in the
Stipa grassland and
Leymus chinensis grassland. In one situation, the effects of grazing on the composition of a soil microbial community caused a significant change [
15], while in another, no change was observed [
3,
16]. These inconsistent results of previous studies may be related to their different ecosystem types, community composition, and climatic conditions.
The Qinghai-Tibet Plateau is an ideal area to study the impact mechanism of global change on the ecosystem. The Qinghai-Tibet Plateau, as the highest altitude region in the world, with an average elevation of more than 4000 m, is known as the “third pole” of the world [
17]. It is one of the most sensitive areas to global climate change and is the regulator and opener of the northern hemisphere climate [
18]. Alpine meadow is an important grassland type on the Tibetan Plateau, which is showing an increase of 0.4 °C every ten years, twice the rate of global warming [
19]. To date, some experimental studies have been carried out on the composition of soil microbial communities in the alpine meadow area of the Qinghai-Tibet Plateau [
11,
13]. Nonetheless, previous studies have mainly focused on the same type of alpine meadow. Therefore, there is a need for further exploration regarding whether soil microbial communities of various dominant species in different types of alpine meadows, as well as those in the same type of alpine meadow but under different environmental conditions, respond differently to warming and clipping. We hypothesized that the response of soil microbial communities of different alpine meadows to warming and clipping would be different. In order to test our hypothesis, this study deployed three experimental warming and clipping experimental platforms (two alpine steppe meadows and one alpine
Kobresia meadow) at the grassland station in Damxung County in the Tibet Autonomous Region. The main objective of this study was to explore the response mechanism of soil microbial community compositions to the experimental warming and clipping treatment in alpine meadows in northern Tibet. It is expected to provide basic research data and a theoretical basis for how to maintain the soil quality and stability of soil microbial communities in the alpine meadows of northern Tibet in the context of global change.
4. Discussion
Although neither warming nor clipping changed the soil G
+/G
− and F/B of the alpine meadow, clipping changed the composition of soil microbial community at the low and high altitude plots and the soil bacteria at high altitudes at
p < 0.10. The interaction of warming and clipping changed the composition of soil bacteria at the moderate altitude plot, which was consistent with the previous research results [
36]. For example, both Wang et al. and Ford et al. found that warming and clipping did not change the F/B value, but the composition of soil microbial communities changed [
36,
37]. Furthermore, Wang et al. found that warming and clipping significantly reduced the value of F/B, while the composition of soil microbial communities did not change in alpine meadows in the Hongyuan County [
38]. Therefore, in the context of global change, changes in the composition of soil microbial communities cannot be inferred just by changes in G
+/G
− and F/B.
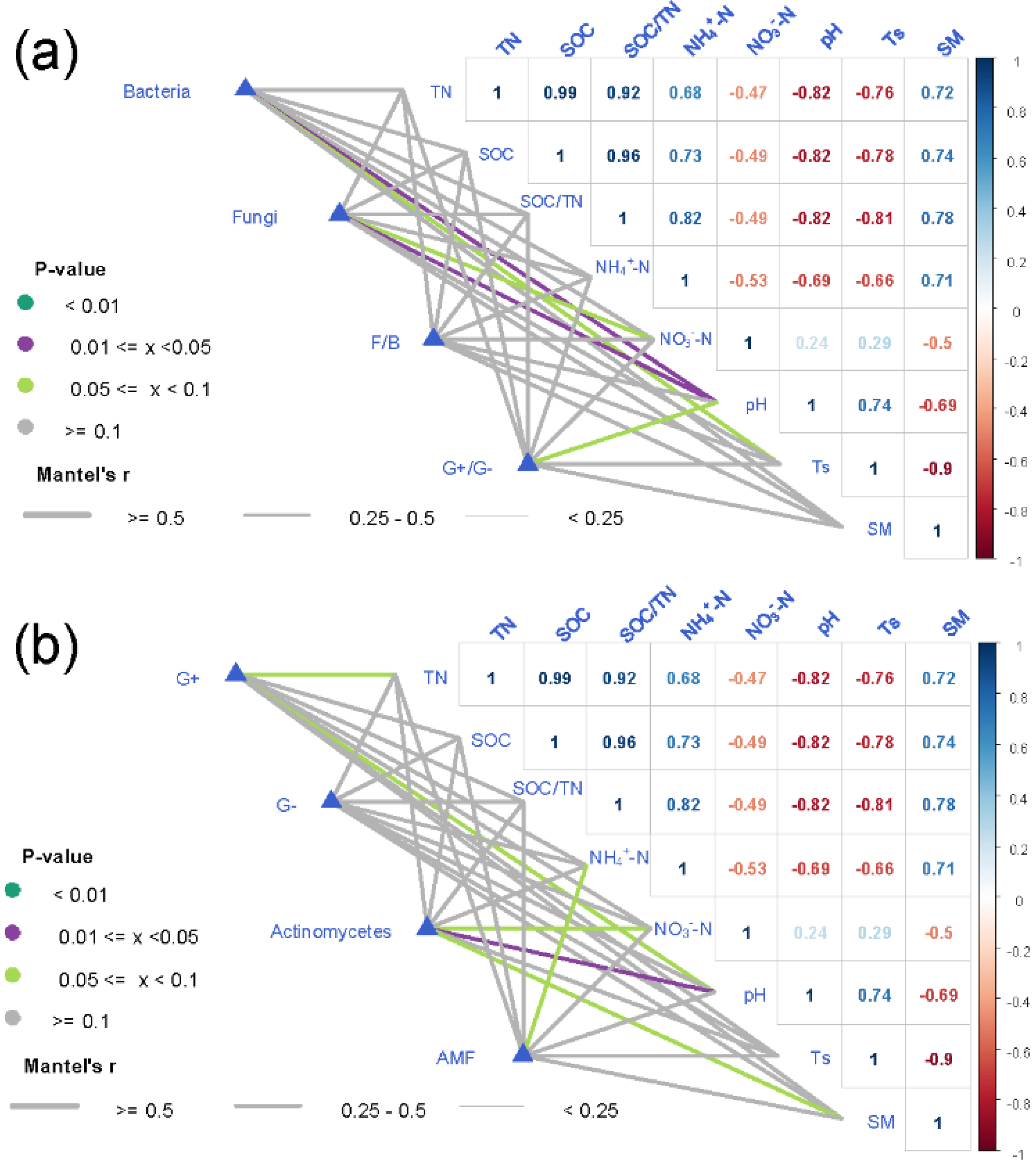
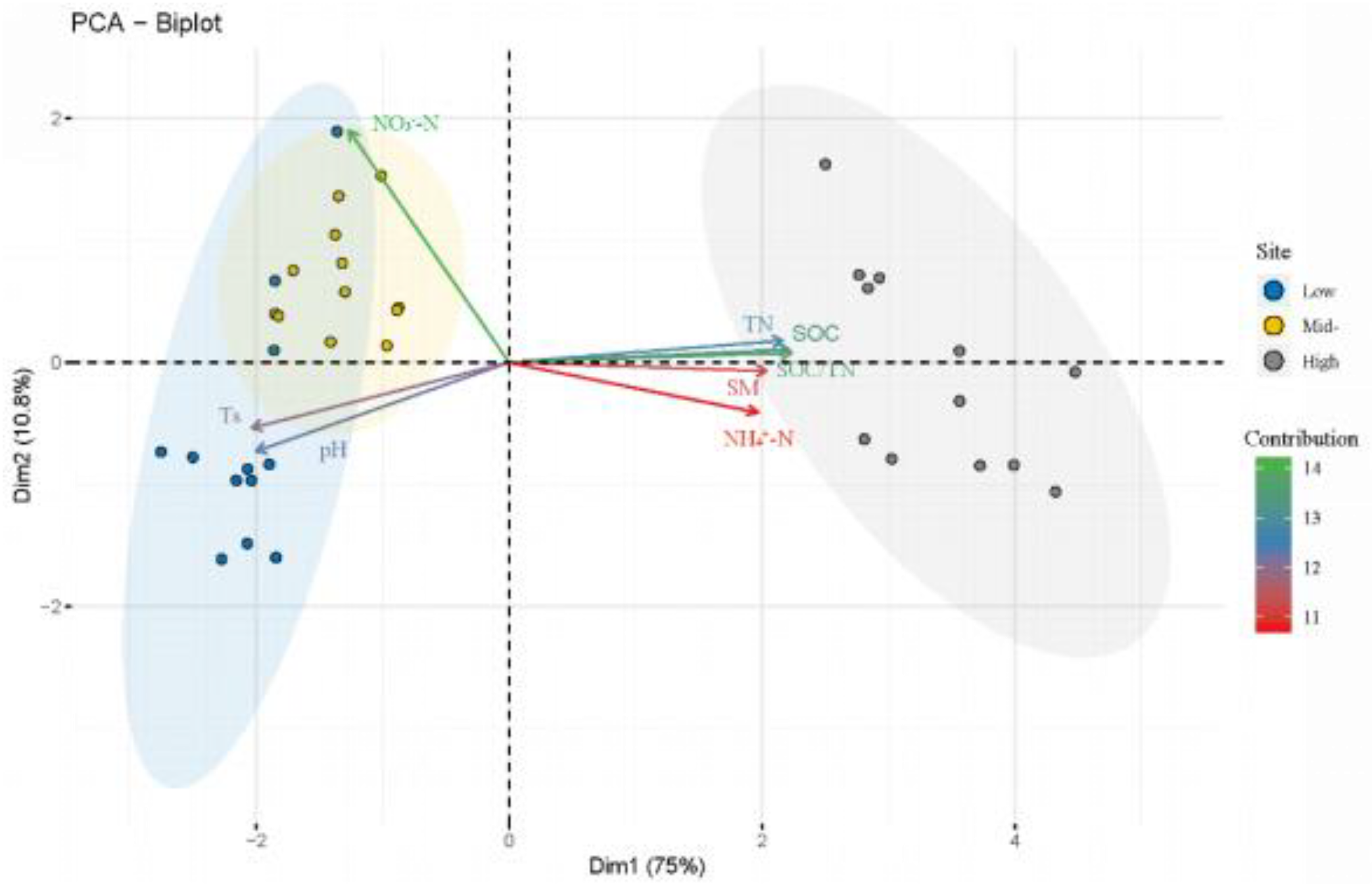
The composition of soil bacteria and fungi was closely related to soil pH (
Figure 2), which was consistent with previous research results [
39]. Soil pH can directly affect the composition of soil microbial community by affecting the enzyme activity, metabolic function, and the ability to absorb nutrients of soil microorganisms [
40,
41], as well as indirectly by affecting the plant community [
42].
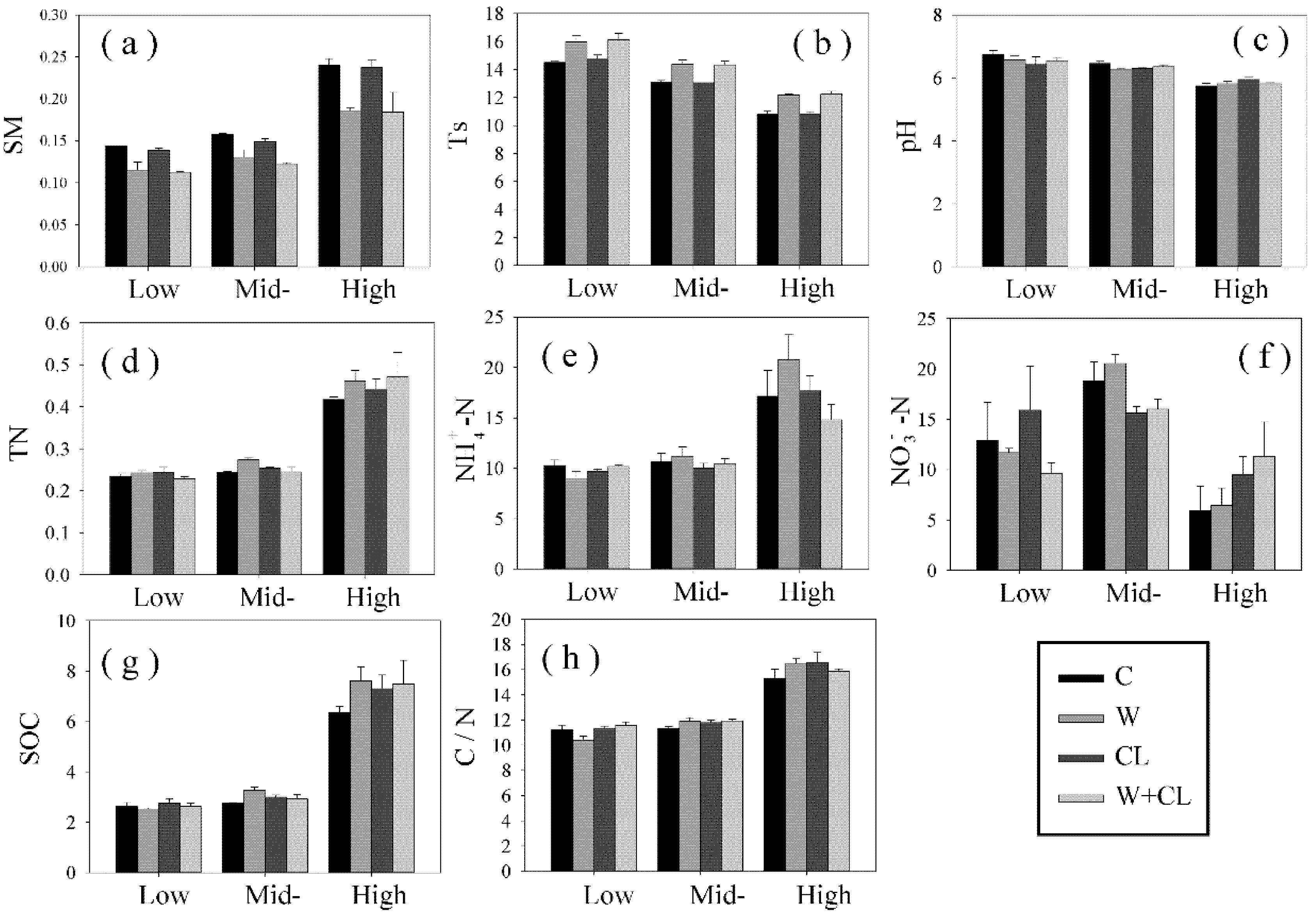
The content of soil PLFA was significantly reduced by warming and clipping. The negative effect of warming may be related to the decrease in soil moisture (
Figure 3 and
Figure A4). The inhibitory effect of soil drought on microorganisms was greater than the promoting effect of soil temperature on microorganisms, which was consistent with the previous research results [
36]. Soil moisture is an important factor affecting soil biological activity [
43]. Soil moisture can limit the transfer of soil nutrients in the soil and inhibit the metabolism of microorganisms. The decrease in soil moisture may lead to a reduction in microbial biomass [
44]. The negative effect of clipping on the microbial community of alpine meadow may be due to the decrease in soil quality and the destruction of nutrient balance caused by clipping [
45,
46]. Sorensen et al. believed that clipping would reduce the conversion rate of soil nitrogen, and the utilization efficiency of biological nitrogen would have a negative effect [
47].
Compared with warming, clipping had a greater impact on the microbial biomass and community composition of bacteria (
Table 3 and
Table A2). This is probably related to the following reasons. (1) Our previous study found that clipping had a greater impact on the productivity of alpine meadow communities in the Northern Tibet than warming [
48]. Community productivity is closely related to the composition of soil microbial communities [
49,
50,
51]. (2) Compared with warming, clipping has a more materiality influence on the composition of plant communities in alpine meadows [
52]. The composition of the plant community can change the quality and quantity of litters, thus causing changes in the microbial population and community composition [
53]. The reduction of litter mass caused by clipping is an important reason for the decrease in microbial quantity. (3) Clipping will reduce plant leaf area and photosynthesis [
54]. Plants meet the demand for nutrients and energy of plants by increasing the absorption of nutrients from the underground part of the soil, thus increasing the competition between plants and microorganisms. (4) Compared with warming, clipping may have a greater impact on soil NO
3−-N. Soil NO
3−-N is an important factor affecting bacterial biomass and community composition [
55,
56,
57].
This study found that soil microbial communities of different types of meadows have different responses to warming and clipping, which was consistent with previous studies [
16,
58]. Compared with alpine meadows, the soil microbial biomass and community composition of typical
Kobresia meadows in the alpine region may be more sensitive to short-term (<6 years) warming and mowing. This may be due to the following reasons. Firstly, the climate conditions of the two types of meadows were different. Compared with the alpine steppe meadow, the alpine meadow was located at a higher altitude, with lower air temperature and soil temperature [
59]. Joseph et al. [
60] found that microbial communities were more susceptible to interference from external environmental factors and more sensitive to climate change at lower temperatures. Fu et al. found that the effect of simulated warming on soil PLFA content and G
− content increased with the increase of altitude [
58]. Zhang et al. also found that the positive effects of warming on soil microbial biomass carbon and nitrogen decreased with increasing mean annual temperature [
61]. Secondly, warming had a stronger promoting effect on soil respiration in the alpine
Kobresia meadow, and the temperature sensitivity of soil respiration increases with the elevation [
62]. The increase of soil respiration reduced the available substrates of soil [
63], and the decrease of soil nutrients may increase the competition among microorganisms. Thirdly, compared with the alpine
Kobresia meadow, the composition of soil bacteria community, fungal community composition, and species alpha diversity of alpine steppe meadow based on high-throughput sequencing technology may have a more intense response to experimental warming (>7 years) [
20]. Short-term simulated warming (<2 years) had no significant effect on the soil microbial carbon and the nitrogen content and carbon-nitrogen ratio of the alpine meadow and alpine
Kobresia meadow [
64]. Therefore, the response of soil microorganisms to experimental warming in the alpine
Kobresia meadow and alpine steppe meadow may also be related to the duration of the experimental warming, the adopted soil microbial indices, and the observation techniques. Fourthly, the alpine steppe meadow and alpine
Kobresia meadow had different sensitivity to grazing. Grazing significantly reduced the soil microbial biomass of the alpine
Kobresia meadow but had no significant effect on the soil microbial biomass of the alpine steppe meadow [
11,
59].






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


