
Management to Support Multiple Ecosystem Services from Productive Grasslands

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experiment
2.2. Provisioning Services (Agronomic Production; Forage Yield and Quality)
2.3. Regulating Services (Pollination and Pest Control)
2.4. Supporting and Regulating Services (Soil Processes)
2.5. Cultural Services (Food Resources for Farmland Birds (BIRDFOOD))
2.6. Data Analysis
3. Results
3.1. Provisioning Services (Agronomic Production, Forage Yield, and Quality)
3.2. Regulating Services (Pollination and Pest Control)
3.3. Supporting and Regulating Services (Soil Processes)
3.3.1. Soil Compaction (Bulk Density)
3.3.2. Soil Nutrients: Total Carbon and Total Nitrogen
3.4. Cultural Services (Food Resources for Farmland Birds (BIRDFOOD))
4. Discussion
4.1. Using Seed Treatments to Achieve Multiple Ecosystem Service Delivery
4.2. Management: Cutting Benefits Multiple Ecosystem Services but Management Intensity Creates Trade-Offs
4.3. Cultivation Techniques Generate Trade-Offs between the Services
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the Fate of Wild Nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [Green Version]
- Bullock, J.M.; Jefferson, R.J.; Blackstock, T.H.; Pakeman, R.J.; Emmett, B.A.; Pywell, R.J.; Grime, J.P.; Silvertown, J. Semi-natural grasslands. In The UK National Ecosystem Assessment Technical Report; UNEP-WCMC: Cambridge, UK, 2011. [Google Scholar]
- Accatino, F.; Tonda, A.; Dross, C.; Léger, F.; Tichit, M. Trade-offs and synergies between livestock production and other ecosystem services. Agric. Syst. 2019, 168, 58–72. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; et al. (Eds.) Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES: Bonn, Germany, 2019. [Google Scholar]
- European Comission. Policy Areas. Agri-Environment Measures. 2017. Available online: https://ec.europa.eu/agriculture/envir/measures_en (accessed on 2 October 2018).
- Batáry, P.; Dicks, L.V.; Kleijn, D.; Sutherland, W.J. The role of agri-environment schemes in conservation and environmental management. Conserv. Biol. 2015, 29, 1006–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pe’Er, G.; Bonn, A.; Bruelheide, H.; Dieker, P.; Eisenhauer, N.; Feindt, P.H.; Hagedorn, G.; Hansjürgens, B.; Herzon, I.; Lomba, Â.; et al. Action needed for the EU Common Agricultural Policy to address sustainability challenges. People Nat. 2020, 2, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Boetzl, F.A.; Krauss, J.; Heinze, J.; Hoffmann, H.; Juffa, J.; Krauss, J.; König, S.; Krimmer, E.; Prante, M.; Martin, E.A.; et al. A multitaxa assessment of the effectiveness of agri-environmental schemes for biodiversity management. Proc. Natl. Acad. Sci. USA 2021, 118, e2016038118. [Google Scholar] [CrossRef] [PubMed]
- Gamero, A.; Brotons, L.; Brunner, A.; Foppen, R.P.B.; Fornasari, L.; Gregory, R.; Herrando, S.; Hořák, D.; Jiguet, F.; Kmecl, P.; et al. Tracking Progress Toward EU Biodiversity Strategy Targets: EU Policy Effects in Preserving its Common Farmland Birds. Conserv. Lett. 2017, 10, 395–402. [Google Scholar] [CrossRef]
- Bradbury, R.B.; Stoate, C.; Tallowin, J.R.B. FORUM: Lowland farmland bird conservation in the context of wider ecosystem service delivery. J. Appl. Ecol. 2010, 47, 986–993. [Google Scholar] [CrossRef]
- Fiedler, A.K.; Landis, D.A.; Wratten, S.D. Maximizing ecosystem services from conservation biological control: The role of habitat management. Biol. Control 2008, 45, 254–271. [Google Scholar] [CrossRef]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- UK NEA. The UK National Ecosystem Assessment: Synthesis of Key Findings; Food and Rural Affairs: London, UK, 2011. [Google Scholar]
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Palm, C.; Blanco-Canqui, H.; DeClerck, F.; Gatere, L.; Grace, P. Conservation agriculture and ecosystem services: An overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef] [Green Version]
- DEFRA. Agriculture in the United Kingdom 2017; DEFRA: London, UK, 2017. [Google Scholar]
- Bianchi, F.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deguines, N.; Jono, C.; Baude, M.; Henry, M.; Julliard, R.; Fontaine, C. Large-scale trade-off between agricultural intensification and crop pollination services. Front. Ecol. Environ. 2014, 12, 212–217. [Google Scholar] [CrossRef]
- Carvell, C.; Bourke, A.F.; Osborne, J.L.; Heard, M.S. Effects of an agri-environment scheme on bumblebee reproduction at local and landscape scales. Basic Appl. Ecol. 2015, 16, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of pollinators and natural enemies: A meta-analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. Camb. Philos. Soc. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Pywell, R.; Macgregor, N.; Edwards, M.; Redhead, J.; Ridding, L.; Batáry, P.; Czerwiński, M.; Duffield, S. Historical, local and landscape factors determine the success of grassland restoration for arthropods. Agric. Ecosyst. Environ. 2021, 308, 107271. [Google Scholar] [CrossRef]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem Services Provided by Birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef]
- Raudsepp-Hearne, C.; Peterson, G.; Bennett, E.M. Ecosystem service bundles for analyzing tradeoffs in diverse landscapes. Proc. Natl. Acad. Sci. USA 2010, 107, 5242–5247. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Woodcock, B.A.; Roberts, S.P.M.; Tscheulin, T.; Pilgrim, E.S.; Brown, V.K.; Tallowin, J.R. Enhancing pollinator biodiversity in intensive grasslands. J. Appl. Ecol. 2009, 46, 369–379. [Google Scholar] [CrossRef]
- Frame, J.; Laidlow, A. Improved Grassland Management 2001; Crowood Press: Marlborough, UK, 2001. [Google Scholar]
- Morris, M.G.; Pollard, E.; Yates, T.J. Monitoring Butterflies for Ecology and Conservation; Chapman and Hall: London, UK, 1993. [Google Scholar]
- Tscharntke, T.; Rand, T.A.; Bianchi, F. The landscape context of trophic interactions: Insect spillover across the crop-noncrop interface. Ann. Zool. Fenn. 2005, 42, 421–432. [Google Scholar]
- Kromp, B. Carabid beetles in sustainable agriculture: A review on pest control efficacy, cultivation impacts and enhancement. Agric. Ecosyst. Environ. 1999, 74, 187–228. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, A.J.; Woodcock, B.A.; Sinka, M.; Vanbergen, A.J. Experimental verification of suction sampler capture efficiency in grasslands of differing vegetation height and structure. J. Appl. Ecol. 2008, 45, 1357–1363. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell: Oxford, UK, 1974. [Google Scholar]
- MAFF. Analysis of Agricultural Materials (RB427); HMSO: London, UK, 1986. [Google Scholar]
- Boeri, M.; Stojanovic, T.A.; Wright, L.J.; Burton, N.H.; Hockley, N.; Bradbury, R.B. Public preferences for multiple dimensions of bird biodiversity at the coast: Insights for the cultural ecosystem services framework. Estuarine, Coast. Shelf Sci. 2020, 235, 106571. [Google Scholar] [CrossRef]
- RSPB. The RSPB Annual Report 2020; RSPB: Sandy, UK, 2020. [Google Scholar]
- U.S. Department of the Interior; USFA World Series; U.S. Department of Commerce; U.S. Census Bureau. National Survey of Fishing, Hunting, and Wildlife-Associated Recreation; Government Printing Office: Washington, DC, USA, 2016. [Google Scholar]
- Vickery, J.A.; Bradbury, R.B.; Henderson, I.G.; Eaton, M.A.; Grice, P.V. The role of agri-environment schemes and farm management practices in reversing the decline of farmland birds in England. Biol. Conserv. 2004, 119, 19–39. [Google Scholar] [CrossRef]
- Møller, A.P. Parallel declines in abundance of insects and insectivorous birds in Denmark over 22 years. Ecol. Evol. 2019, 9, 6581–6587. [Google Scholar] [CrossRef] [Green Version]
- Westbury, D.B.; Woodcock, B.; Harris, S.; Brown, V.; Potts, S. Buffer strip management to deliver plant and invertebrate resources for farmland birds in agricultural landscapes. Agric. Ecosyst. Environ. 2017, 240 (Suppl. C), 215–223. [Google Scholar] [CrossRef]
- Rogers, L.E.; Hinds, W.T.; Buschbom, R.L. A general weight vs. length relationship for insects. Ann. Entomol. Soc. N. Am. 1976, 69, 387–389. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B. Lme4: Linear Mixed-Effects Models Using S4 Classes.; R package version 0.999999-2. 2013. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 1 April 2020).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.9.13. 2013. Available online: http://mtweb.cs.ucl.ac.uk/mus/www/preCC/Sparse_R_LIB/lme4/html/00Index.html (accessed on 1 April 2020).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practice Information-Theoretic Approach; Springer: New York, NY, USA, 1998. [Google Scholar]
- Symonds, M.R.E.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Woodcock, B.; Savage, J.; Bullock, J.; Nowakowski, M.; Orr, R.; Tallowin, J.; Pywell, R. Enhancing floral resources for pollinators in productive agricultural grasslands. Biol. Conserv. 2014, 171, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Met Office National Climate Information Centre. Available online: https://www.metoffice.gov.uk/research/climate/maps-and-data/historic-station-data (accessed on 11 May 2021).
- Qi, A.; Murray, P.J.; Richter, G.M. Modelling productivity and resource use efficiency for grassland ecosystems in the UK. Eur. J. Agron. 2017, 89, 148–158. [Google Scholar] [CrossRef]
- Brophy, C.; Finn, J.A.; Luscher, A.; Suter, M.; Kirwan, L.; Sebastia, T.; Helgadottir, A.; Baadshaug, O.H.; Belanger, G.; Black, A.; et al. Major shifts in species’ relative abundance in grassland mixtures alongside positive effects of species diversity in yield: A continental-scale experiment. J. Ecol. 2017, 105, 1210–1222. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.; Buchmann, N.; Buscot, F.; et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.-F.; Rees, R.M.; Peyraud, J.-L. Potential of legume-based grassland—livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wright, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Natl. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Dukes, J.S. Overyielding among plant functional groups in a long-term experiment. Ecol. Lett. 2004, 7, 95–105. [Google Scholar] [CrossRef]
- Sutton, M.A.; Howard, C.M.; Erisman, J.W.; Billen, G.; Bleeker, A.; Grennfelt, P.; van Grinsven, H.; Grizzetti, B. The European Nitrogen Assessment (ENA); Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Kormann, U.; Rösch, V.; Batáry, P.; Tscharntke, T.; Orci, K.M.; Samu, F.; Scherber, C. Local and landscape management drive trait-mediated biodiversity of nine taxa on small grassland fragments. Divers. Distrib. 2015, 21, 1204–1217. [Google Scholar] [CrossRef]
- Williams, N.M.; Ward, K.L.; Pope, N.; Isaacs, R.; Wilson, J.; May, E.A.; Ellis, J.; Daniels, J.C.; Pence, A.; Ullmann, K.S.; et al. Native wildflower plantings support wild bee abundance and diversity in agricultural landscapes across the United States. Ecol. Appl. 2015, 25, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- He, X.; Kiær, L.P.; Jensen, P.M.; Sigsgaard, L. The effect of floral resources on predator longevity and fecundity: A systematic review and meta-analysis. Biol. Control 2021, 153, 104476. [Google Scholar] [CrossRef]
- M’Gonigle, L.K.; Williams, N.M.; Lonsdorf, E.; Kremen, C. A Tool for Selecting Plants When Restoring Habitat for Pollinators. Conserv. Lett. 2017, 10, 105–111. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Goulson, D.; Nowakowski, M. Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. J. Appl. Ecol. 2007, 44, 29–40. [Google Scholar] [CrossRef]
- Vickery, J.A.; Tallowin, J.R.; Feber, R.E.; Asteraki, E.J.; Atkinson, P.W.; Fuller, R.J.; Brown, V.K. The management of lowland neutral grasslands in Britain: Effects of agricultural practices on birds and their food resources. J. Appl. Ecol. 2001, 38, 647–664. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Wu, J. Grassland ecosystem services: A systematic review of research advances and future directions. Landsc. Ecol. 2020, 35, 793–814. [Google Scholar] [CrossRef]
- Marselle, M.R.; Hartig, T.; Cox, D.T.; de Bell, S.; Knapp, S.; Lindley, S.; Triguero-Mas, M.; Böhning-Gaese, K.; Braubach, M.; Cook, P.A.; et al. Pathways linking biodiversity to human health: A conceptual framework. Environ. Int. 2021, 150, 106420. [Google Scholar] [CrossRef] [PubMed]
- Sandifer, P.A.; Sutton-Grier, A.E.; Ward, B.P. Exploring connections among nature, biodiversity, ecosystem services, and human health and well-being: Opportunities to enhance health and biodiversity conservation. Ecosyst. Serv. 2015, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Keniger, L.E.; Gaston, K.J.; Irvine, K.N.; Fuller, R.A. What are the benefits of interacting with nature? Int. J. Environ. Res. Public Health 2013, 10, 913–935. [Google Scholar] [CrossRef] [Green Version]
- Clark, N.E.; Lovell, R.; Wheeler, B.W.; Higgins, S.L.; Depledge, M.H.; Norris, K. Biodiversity, cultural pathways, and human health: A framework. Trends Ecol. Evol. 2014, 29, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Bratman, G.N.; Anderson, C.B.; Berman, M.G.; Cochran, B.; De Vries, S.; Flanders, J.; Folke, C.; Frumkin, H.; Gross, J.J.; Hartig, T.; et al. Nature and mental health: An ecosystem service perspective. Sci. Adv. 2019, 5, eaax0903. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Health Assembly 65. Global Burden of Mental Disorders and the Need for a Comprehensive, Coordinated Response from Health and Social Sectors at the Country Level: Report by the Secretariat; World Health Organization: Geneva, Switzerland, 2012; Available online: https://apps.who.int/iris/handle/10665/78898 (accessed on 18 May 2021).
- D’Aniello, B.; Stanislao, I.; Bonelli, S.; Balletto, E. Haying and grazing effects on the butterfly communities of two Mediterranean-area grasslands. Biodivers. Conserv. 2011, 20, 1731. [Google Scholar] [CrossRef]
- Smith, L.M.; Cherry, R. Effects of Management Techniques on Grassland Butterfly Species Composition and Community Structure. Am. Midl. Nat. 2014, 172, 227–235. [Google Scholar] [CrossRef]
- Tälle, M.; Deák, B.; Poschlod, P.; Valkó, O.; Westerberg, L.; Milberg, P. Grazing vs. mowing: A meta-analysis of biodiversity benefits for grassland management. Agric. Ecosyst. Environ. 2016, 222, 200–212. [Google Scholar] [CrossRef]
- Turner, K.G.; Odgaard, M.V.; Bøcher, P.K.; Dalgaard, T.; Svenning, J.-C. Bundling ecosystem services in Denmark: Trade-offs and synergies in a cultural landscape. Landsc. Urban. Plan. 2014, 125, 89–104. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhao, Y.; Yu, C.; Luo, L.; Pan, Y. Land management influences trade-offs and the total supply of ecosystem services in alpine grassland in Tibet, China. J. Environ. Manag. 2017, 193, 70–78. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, T.; Oteros-Rozas, E.; Ripoll-Bosch, R.; Tichit, M.; Martin-Lopez, B.; Bernués, A. Applying the ecosystem services framework to pasture-based livestock farming systems in Europe. Animal 2014, 8, 1361–1372. [Google Scholar] [CrossRef] [Green Version]
- Teillard, F.; Doyen, L.; Dross, C.; Jiguet, F.; Tichit, M. Optimal allocations of agricultural intensity reveal win-no loss solutions for food production and biodiversity. Reg. Environ. Chang. 2017, 17, 1397–1408. [Google Scholar] [CrossRef]
- Nelson, E.J.; Daily, G.C. Daily, Modelling ecosystem services in terrestrial systems. F1000 Biol. Rep. 2010, 2, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kaisi, M.M.; Yin, X.; Licht, M.A. Soil carbon and nitrogen changes as affected by tillage system and crop biomass in a corn–soybean rotation. Appl. Soil Ecol. 2005, 30, 174–191. [Google Scholar] [CrossRef]
- West, T.; Post, W.M. Soil Organic Carbon Sequestration Rates by Tillage and Crop Rotation. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef] [Green Version]
- Powlson, D.S.; Stirling, C.M.; Jat, M.L.; Gerard, B.G.; Palm, C.A.; Sanchez, P.A.; Cassman, K.G. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Chang. 2014, 4, 678–683. [Google Scholar] [CrossRef]
- Malinga, R.; Gordon, L.J.; Jewitt, G.; Lindborg, R. Mapping ecosystem services across scales and continents—A review. Ecosyst. Serv. 2015, 13, 57–63. [Google Scholar] [CrossRef]
- Englund, O.; Berndes, G.; Cederberg, C. How to analyse ecosystem services in landscapes—A systematic review. Ecol. Indic. 2017, 73, 492–504. [Google Scholar] [CrossRef]
- Agudelo, C.A.R.; Bustos, S.L.H.; Moreno, C.A.P. Moreno, Modeling interactions among multiple ecosystem services. A critical review. Ecol. Model. 2020, 429, 109103. [Google Scholar] [CrossRef]
- Dunford, R.; Harrison, P.; Smith, A.; Dick, J.; Barton, D.N.; Martin-Lopez, B.; Kelemen, E.; Jacobs, S.; Saarikoski, H.; Turkelboom, F.; et al. Integrating methods for ecosystem service assessment: Experiences from real world situations. Ecosyst. Serv. 2018, 29, 499–514. [Google Scholar] [CrossRef] [Green Version]
- O’Mara, F.P. The role of grasslands in food security and climate change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- de Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Savage, J.; Woodcock, B.A.; Bullock, J.M.; Peyton, J.; Hulmes, S.; Hulmes, L.; Nowakowski, M.; Pywell, R.F. Impact of Grassland Management on Biomass Production and Nutritional Quality, Invertebrate Communities, and Soil Health in Berkshire (UK) 2009–2012; NERC Environmental Information Data Centre, Ed.; NERC Environmental Information Data Centre: London, UK, 2019. [Google Scholar]


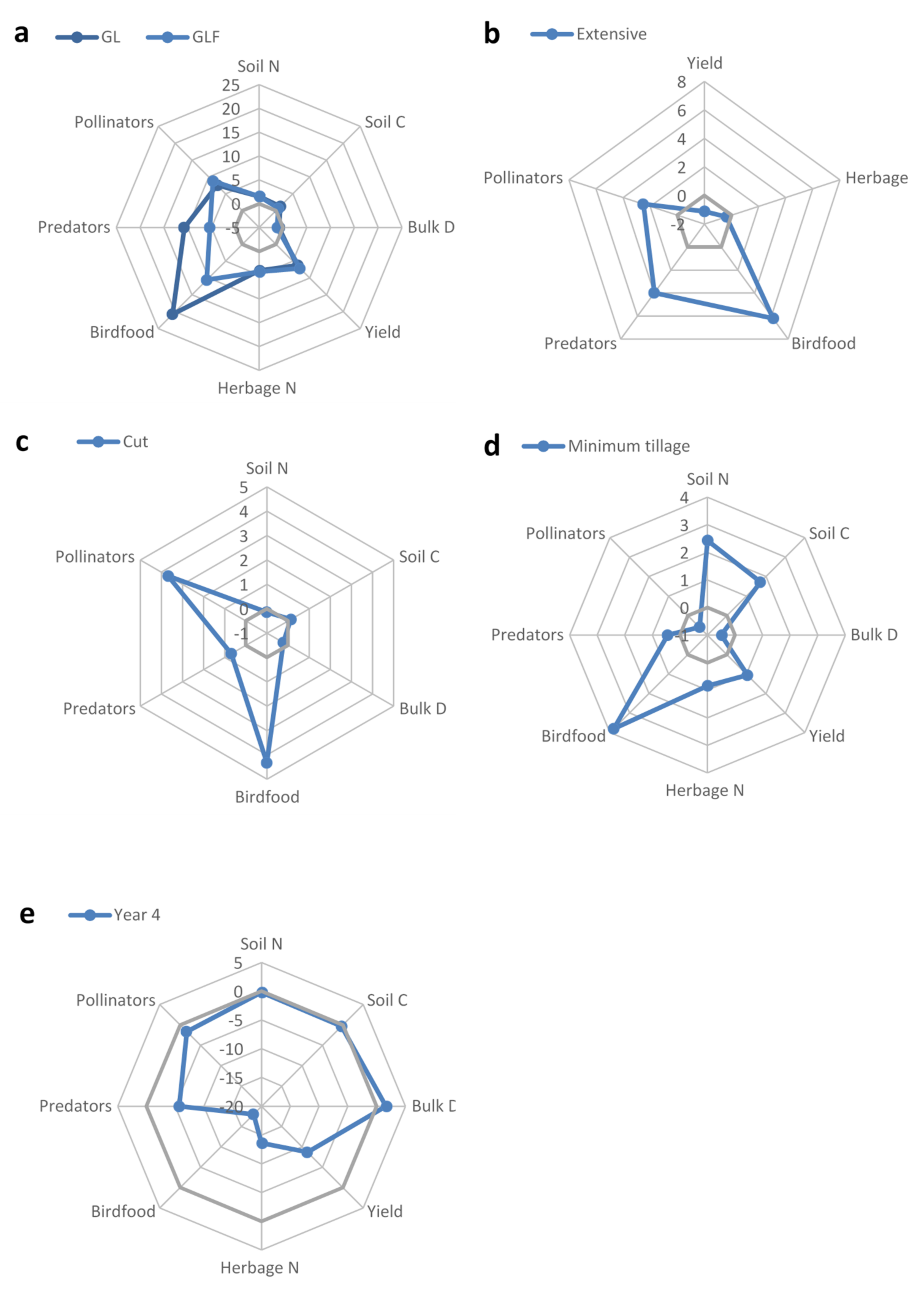{kind=link}
| Response | Models ΔAIC < 2 | Seed Mix | Management | Management Intensity | Cultivation | Year |
|---|---|---|---|---|---|---|
| Agronomic Production; Forage Yield and Quality | ||||||
| Dry Matter Yield | 3 | GLF > GL > G (μ: 7.83 > 6.94 > 3.33) ∑wi = 1.00 | -- | Int. > Ext. (μ: 7.36 > 6.94) ∑wi = 0.77 | Min.till. > Deep (μ: 7.3 > 6.94) ∑wi = 0.71 | Yr1 > Yr4 (μ: 6.94 > 2.73) ∑wi = 1.00 |
| Herbage Nitrogen Content | 3 | GLF > GL > G (μ: 1.98 > 1.97 > 1.64) ∑wi = 1.00 | -- | ∑wi = 0.16 | ∑wi = 0.42 | Yr1 > Yr4 (μ: 1.97 > 1.10) ∑wi = 1.00 |
| Pollination & Pest Control | ||||||
| Pollinator Abundance | 2 | GLF > GL > G (μ: 92.4 > 50.2 > 3.50) ∑wi = 1.00 | Cut > Graze (μ: 50.2 > 32.5) ∑wi = 1.00 | Ext. > Int. (μ: 50.4 > 31.4) ∑wi = 1.00 | Deep > Min.till. (μ: 50.2 > 46.7) ∑wi = 0.79 | Yr1 > Yr4 (μ: 50.2 > 27.9) ∑wi = 1.00 |
| Predator Abundance | 3 | GL > GLF > G (μ: 32.6 > 32.2 > 10.0) ∑wi = 1.00 | ∑wi = 0.25 | Ext. > Int. (μ: 32.6 > 19.4) ∑wi = 1.00 | ∑wi = 0.25 | Yr1 > Yr4 (μ: 32.6 > 11.4) ∑wi = 1.00 |
| Soil Processes | ||||||
| Bulk Density | 6 | ∑wi = 0.24 | ∑wi = 0.23 | -- | ∑wi = 0.24 | ∑wi = 0.20 |
| Total Carbon | 6 | ∑wi = 0.24 | ∑wi = 0.19 | -- | Min.till. > Deep (μ = 2.88 > 2.7) ∑wi = 1.00 | ∑wi = 0.34 |
| Total Nitrogen | 6 | GLF = GL > G (μ: 0.26 = 0.26 > 0.25) ∑wi = 0.69 | ∑wi = 0.17 | -- | Min.till. > Deep (μ: 0.27 > 0.26) ∑wi = 1.00 | ∑wi = 0.39 |
| Cultural Services (Food Resources for Farmland Birds (BIRDFOOD) | ||||||
| Total Invertebrate Biomass | 1 | GLF > GL > G (μ: 144.3 > 133.5 > 67.8) ∑wi = 1.00 | Cut > Graze (μ: 133.5 > 112.2) ∑wi = 1.00 | Ext. > Int. (μ: 133.5 > 107.3) ∑wi = 1.00 | Min.till. > Deep (μ: 149.7 > 133.5) ∑wi = 0.90 | Yr1 > Yr4 (μ: 133.5 > 65.5) ∑wi = 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savage, J.; Woodcock, B.A.; Bullock, J.M.; Nowakowski, M.; Tallowin, J.R.B.; Pywell, R.F. Management to Support Multiple Ecosystem Services from Productive Grasslands. Sustainability 2021, 13, 6263. https://doi.org/10.3390/su13116263
Savage J, Woodcock BA, Bullock JM, Nowakowski M, Tallowin JRB, Pywell RF. Management to Support Multiple Ecosystem Services from Productive Grasslands. Sustainability. 2021; 13(11):6263. https://doi.org/10.3390/su13116263
Chicago/Turabian StyleSavage, Joanna, Ben A. Woodcock, James M. Bullock, Marek Nowakowski, Jeremy R. B. Tallowin, and Richard F. Pywell. 2021. "Management to Support Multiple Ecosystem Services from Productive Grasslands" Sustainability 13, no. 11: 6263. https://doi.org/10.3390/su13116263
APA StyleSavage, J., Woodcock, B. A., Bullock, J. M., Nowakowski, M., Tallowin, J. R. B., & Pywell, R. F. (2021). Management to Support Multiple Ecosystem Services from Productive Grasslands. Sustainability, 13(11), 6263. https://doi.org/10.3390/su13116263