
Investigating the Patterns and Controls of Ecosystem Light Use Efficiency with the Data from the Global Farmland Fluxdata Network



Abstract
:1. Introduction
2. Materials and Methods
2.1. EC Fluxes and Meteorological Data
2.2. Normalized Difference Vegetation Index (NDVI)
2.3. Derived Variables
2.4. Path Analysis Theory
2.5. Regression Analysis
3. Results and Discussion
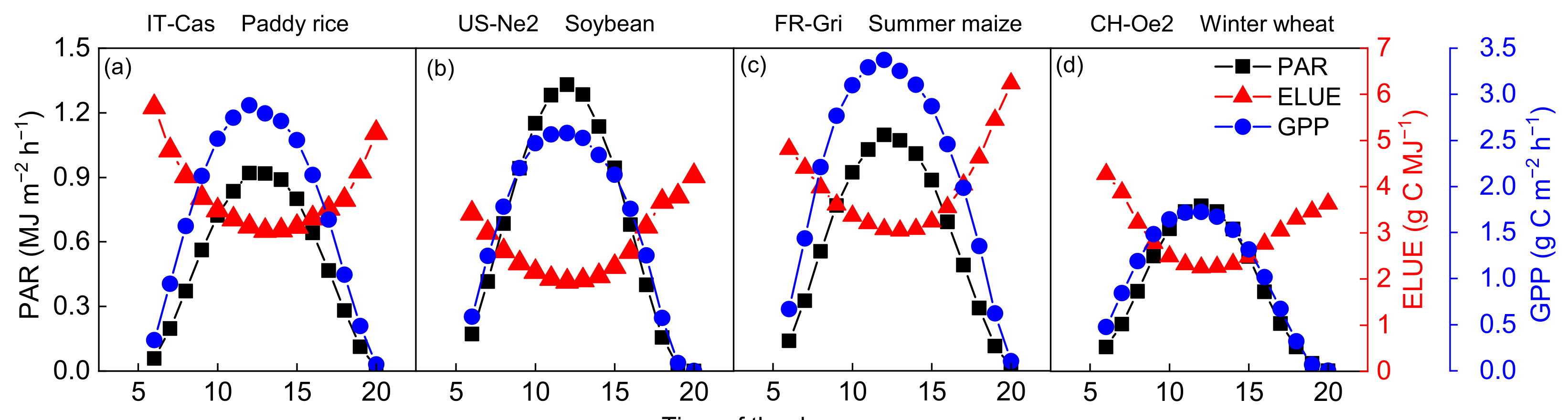
3.1. Variation Patterns of GPP, PAR and ELUE on Different Time Scales
3.1.1. Diurnal Characteristics
3.1.2. Seasonal Variations
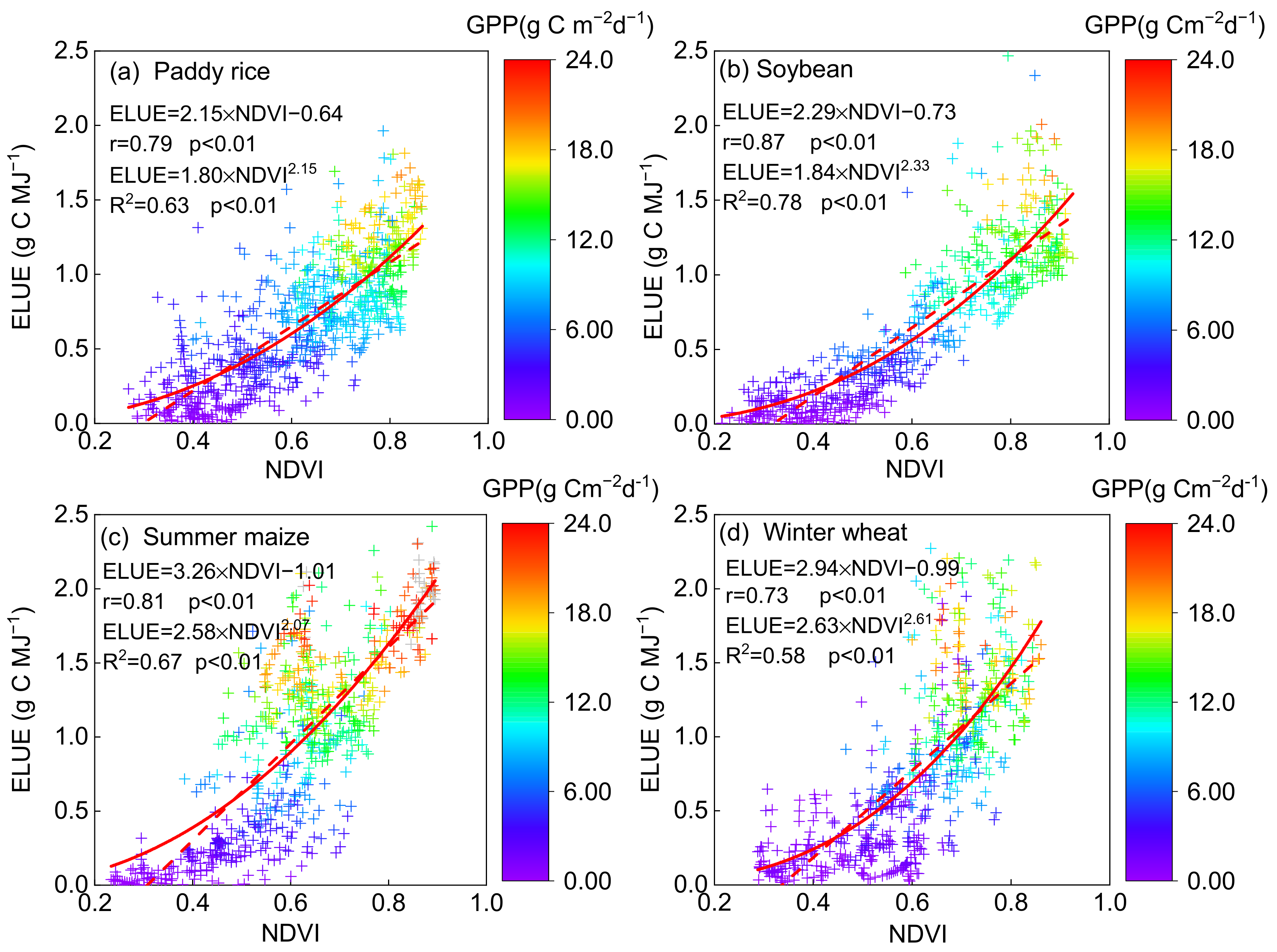
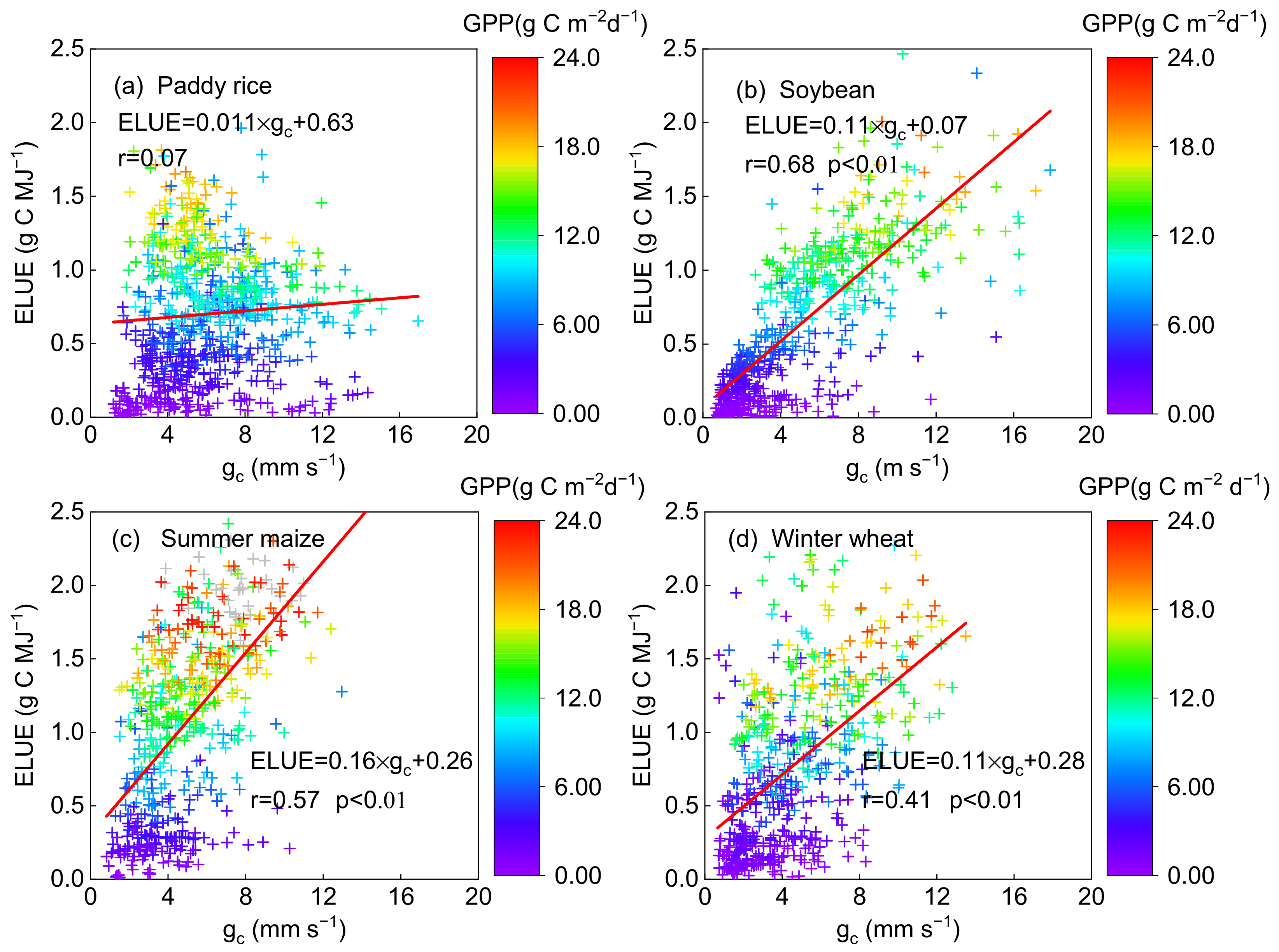
3.2. Responses of ELUE and GPP to Bio-Physical and Environmental Factors
3.2.1. Responses of ELUE and GPP to Vegetation and Physiological Factors
3.2.2. Responses of ELUE and GPP to Climatic Factors
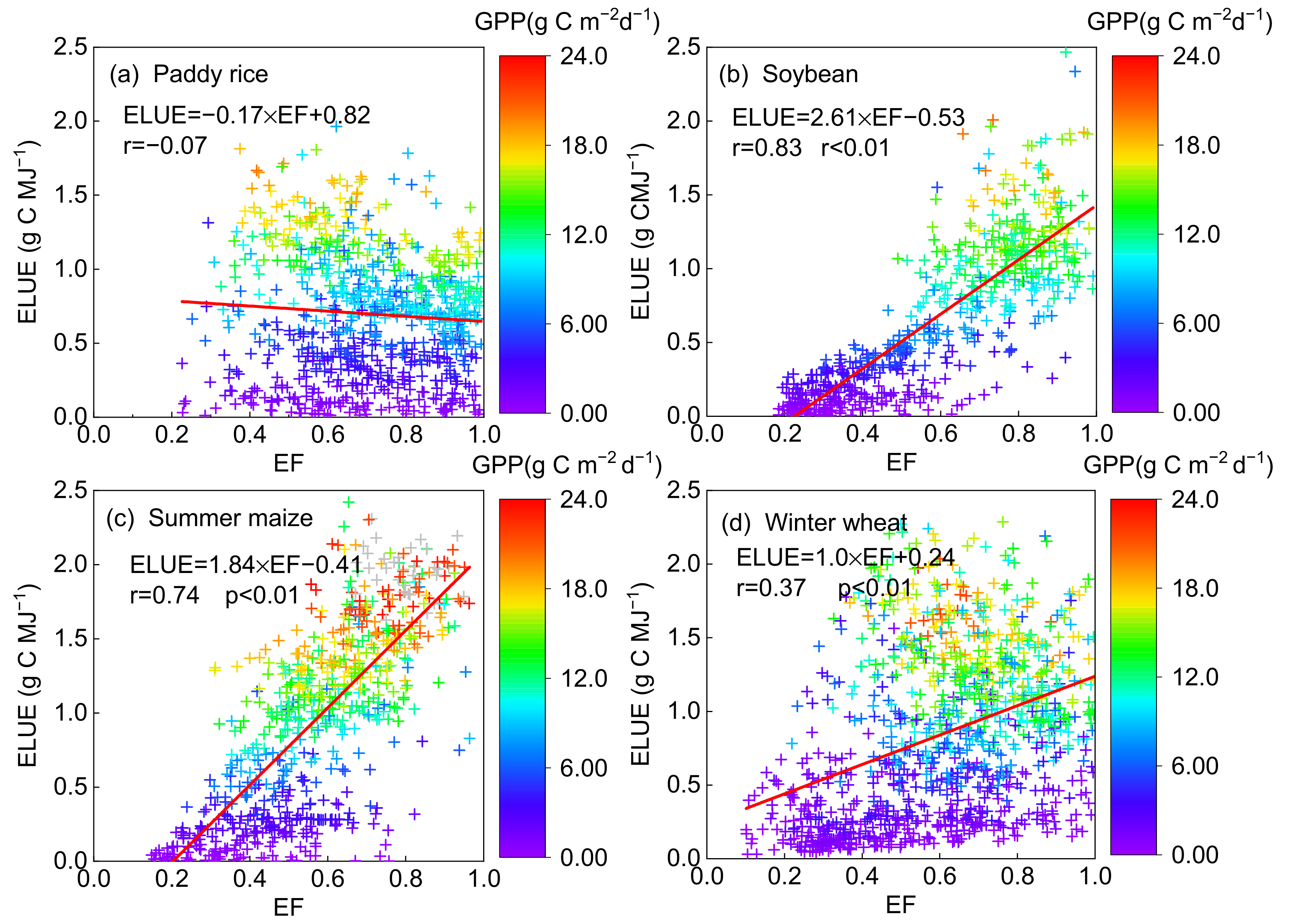
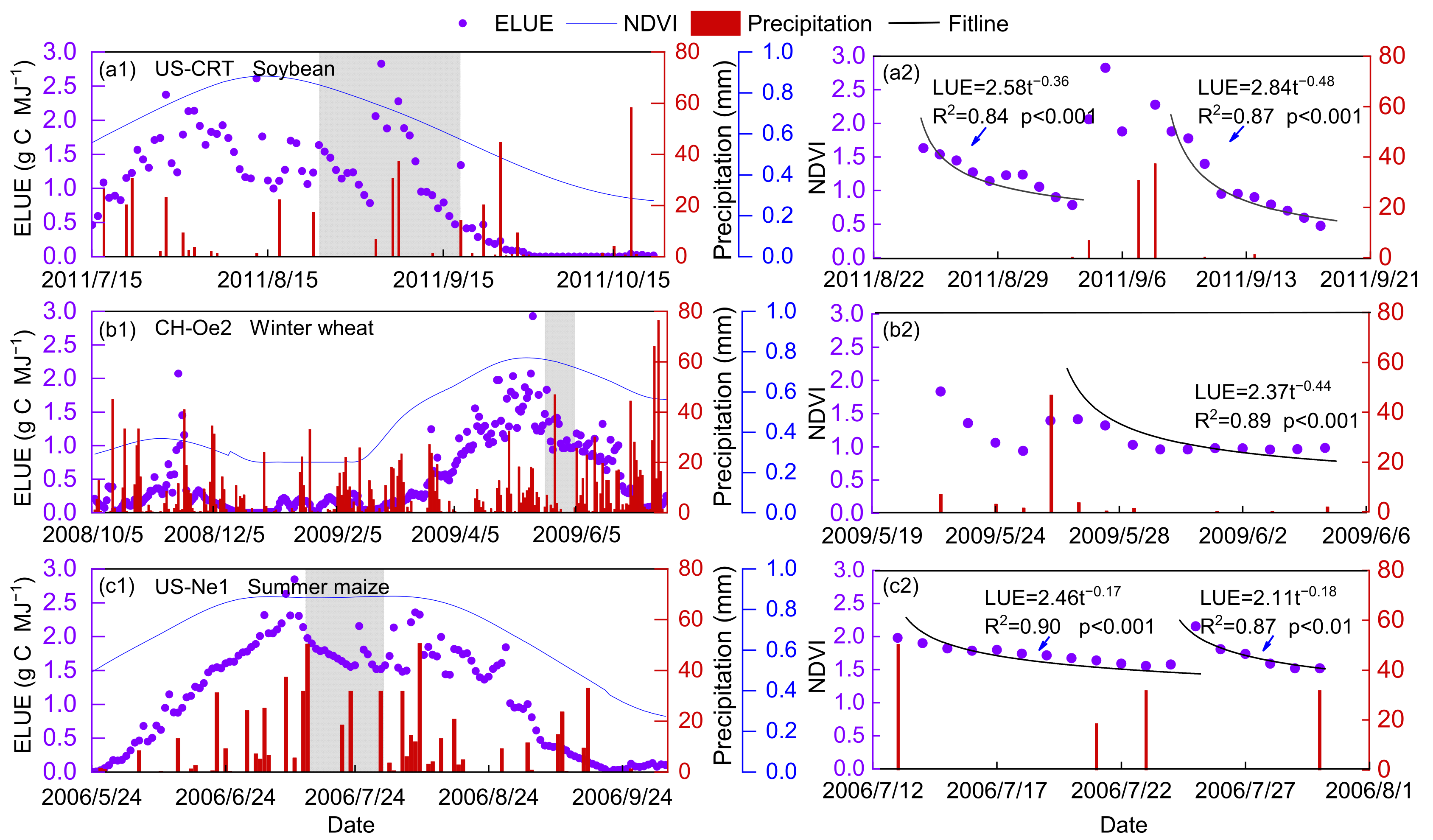
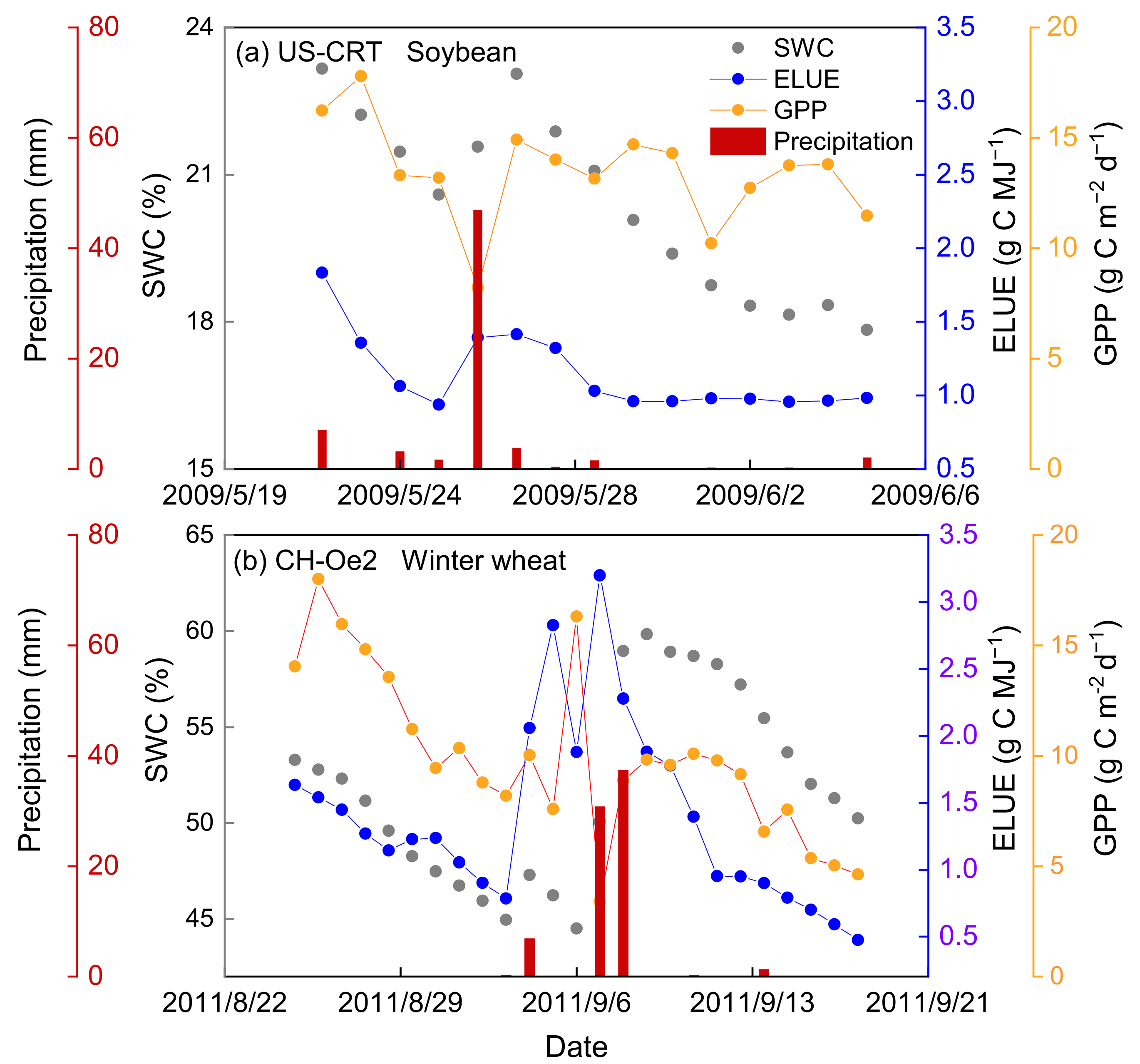
3.2.3. Responses of ELUE and GPP to Water Availability
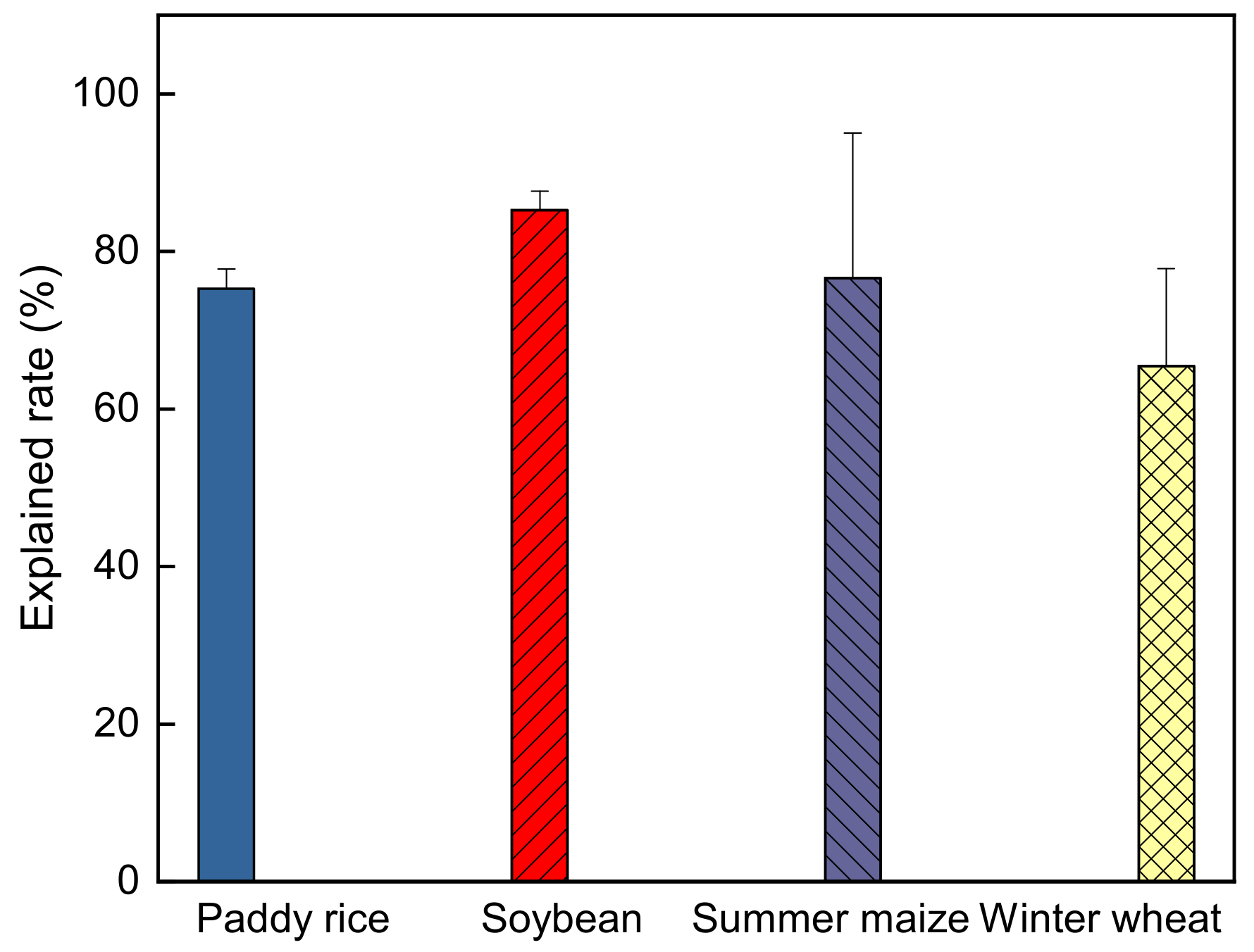
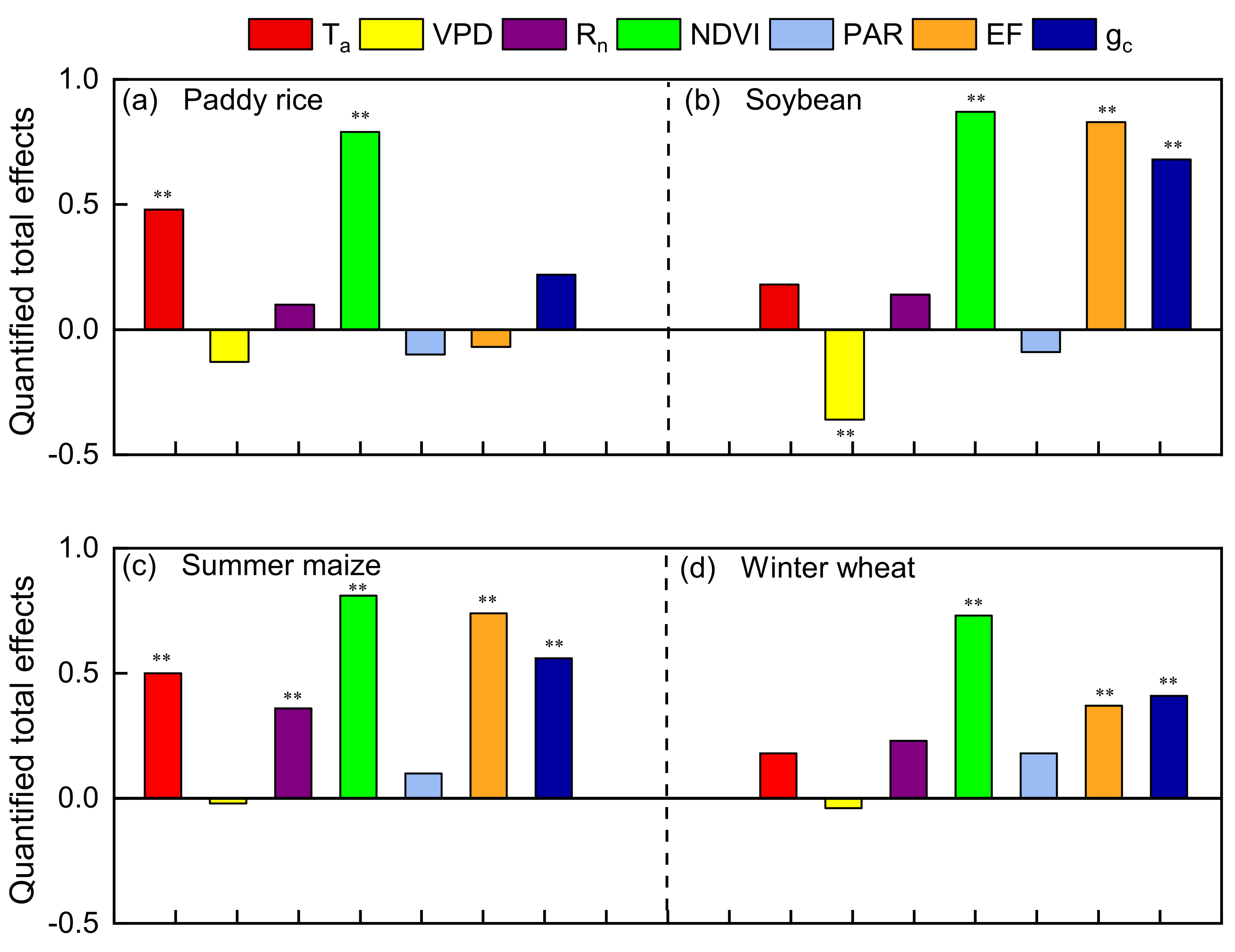
3.3. Comparison of Relative Effects of Critical Factors on ELUE by Path Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
| ELUE | Ecosystem light use efficiency (g C MJ−1) |
| GPP | Gross primary productivity (g C m−2 d−1) |
| NEE | Net ecosystem exchange of CO2 (g C m−2 d−1) |
| ER | Ecosystem respiration (g C m−2 d−1) |
| Ta | Air temperature (°C) |
| NDVI | Normalized difference vegetation index (dimensionless) |
| gc | Canopy conductance (mm s−1) |
| EF | evaporative fraction (dimensionless) |
| VPD | Vapor water deficit (hPa) |
| Rg | Downward shortwave radiation (W m−2) |
| PAR | Photosynthetically active radiation (W m−2) |
| Rn | Net solar radiation (W m−2) |
| LE | Latent heat flux (W m−2) |
| H | Sensible heat flux (W m−2) |
| RH | Relative humidity (%) |
| VPD | Vapor pressure deficit (hpa) |
| γ | Psychrometric constant (0.066 kPa °C−1) |
| Δ | Slope of saturation vapor pressure curve at Ta (kPa °C−1) |
| ρa | Air density (kg m−3) |
| cp | Air specific heat capacity (J kg−1 K−1) |
| ga | Aerodynamic conductance (m s−1) |
| u* | Faction velocity (m s−1) |
References
- Ruimy, A.; Kergoat, L.; Bondeau, A. Comparing global models of terrestrial net primary productivity (NPP): Analysis of differences in light absorption and light-use efficiency. Glob. Chang. Biol. 1999, 5, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, R.A.; Bertero, H.D. Light interception and radiation use efficiency in temperate quinoa (Chenopodium quinoa Willd.) cultivars. Eur. J. Agron. 2008, 29, 144–152. [Google Scholar] [CrossRef]
- Kukal, M.S.; Irmak, S. Evidence of arithmetical uncertainty in estimation of light and water use effciency. Sustainability 2020, 12, 2271. [Google Scholar] [CrossRef] [Green Version]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, G.; Yan, H.; Zhu, X.; Li, S.; Wang, Q.; Zhang, J.; Wang, Y.; Li, Y.; Zhao, L.; et al. A MODIS-based Photosynthetic Capacity Model to estimate gross primary production in Northern China and the Tibetan Plateau. Remote Sens. Environ. 2014, 148, 108–118. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbulsky, M.F.; Peñuelas, J.; Papale, D.; Ardö, J.; Goulden, M.L.; Kiely, G.; Richardson, A.D.; Rotenberg, E.; Veenendaal, E.M.; Filella, I. Patterns and controls of the variability of radiation use efficiency and primary productivity across terrestrial ecosystems. Glob. Ecol. Biogeogr. 2010, 19, 253–267. [Google Scholar] [CrossRef]
- Williams, C.A.; Albertson, J.D. Soil moisture controls on canopy-scale water and carbon fluxes in an African savanna. Water Resour. Res. 2004, 40, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Kromdijk, J.; Heuvelink, E.; van Noort, F.R.; Kaiser, E.; Marcelis, L.F.M. Effects of diffuse light on radiation use efficiency of two Anthurium cultivars depend on the response of stomatal conductance to dynamic light intensity. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Peng, Y.; Arkebauer, T.J.; Schepers, J. Relationships between gross primary production, green LAI, and canopy chlorophyll content in maize: Implications for remote sensing of primary production. Remote Sens. Environ. 2014, 144, 65–72. [Google Scholar] [CrossRef]
- Medlyn, B.E. Physiological basis of the light use efficiency model. Tree Physiol. 1998, 18, 167–176. [Google Scholar] [CrossRef]
- Baldocchi, D.; Knox, S.; Dronova, I.; Verfaillie, J.; Oikawa, P.; Sturtevant, C.; Matthes, J.H.; Detto, M. The impact of expanding flooded land area on the annual evaporation of rice. Agric. For. Meteorol. 2016, 223, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Peñuelas, J.; Seneviratne, S.I. Quantifying soil moisture impacts on light use efficiency across biomes. New Phytol. 2018, 218, 1430–1449. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Peng, Y.; Huemmrich, K.F. Relationship between fraction of radiation absorbed by photosynthesizing maize and soybean canopies and NDVI from remotely sensed data taken at close range and from MODIS 250m resolution data. Remote Sens. Environ. 2014, 147, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Jin, J.; Jiang, H. Strong impacts of autumn phenology on grassland ecosystem water use efficiency on the Tibetan Plateau. Ecol. Indic. 2021, 126, 107682. [Google Scholar] [CrossRef]
- Alton, P.B.; North, P.R.; Los, S.O. The impact of diffuse sunlight on canopy light-use efficiency, gross photosynthetic product and net ecosystem exchange in three forest biomes. Glob. Chang. Biol. 2007, 13, 776–787. [Google Scholar] [CrossRef]
- Niu, S.; Wu, M.; Han, Y.; Xia, J.; Li, L.; Wan, S. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. New Phytol. 2008, 177, 209–219. [Google Scholar] [CrossRef]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. Correlation and causation. J. Agric. Food Res. 1921, 20, 557–585. [Google Scholar]
- Xing, X.; Liu, Y.; Zhao, W.; Kang, D.; Yu, M.; Ma, X. Determination of dominant weather parameters on reference evapotranspiration by path analysis theory. Comput. Electron. Agric. 2016, 120, 10–16. [Google Scholar] [CrossRef]
- Fei, X.H.; Song, Q.H.; Zhang, Y.P.; Yu, G.R.; Zhang, L.M.; Sha, L.Q.; Liu, Y.T.; Xu, K.; Chen, H.; Wu, C.S.; et al. Patterns and Controls of Light Use Efficiency in Four Contrasting Forest Ecosystems in Yunnan, Southwest China. J. Geophys. Res.-Biogeosci. 2019, 124, 293–311. [Google Scholar] [CrossRef]
- Jiang, S.; Liang, C.; Cui, N.; Zhao, L.; Liu, C.; Feng, Y.; Hu, X.; Gong, D.; Zou, Q. Water use efficiency and its drivers in four typical agroecosystems based on flux tower measurements. Agric. For. Meteorol. 2020, 295. [Google Scholar] [CrossRef]
- Heinsch, F.A.; Zhao, M.; Running, S.W.; Kimball, J.S.; Nemani, R.R.; Davis, K.J.; Bolstad, P.V.; Cook, B.D.; Desai, A.R.; Ricciuto, D.M.; et al. Evaluation of remote sensing based terrestrial productivity from MODIS using regional tower eddy flux network observations. IEEE Trans. Geosci. Remote Sens. 2006, 44, 1908–1923. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Tang, X.; Zheng, C.; Gu, Q.; Wei, J.; Ma, M. Differences in ecosystem water-use efficiency among the typical croplands. Agric. Water Manag. 2018, 209, 142–150. [Google Scholar] [CrossRef]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Cai, W.; Xia, J.; Chen, J.; Liu, S.; Dong, W.; Merbold, L.; Law, B.; Arain, A.; Beringer, J.; et al. Global comparison of light use efficiency models for simulating terrestrial vegetation gross primary production based on the LaThuile database. Agric. For. Meteorol. 2014, 192–193, 108–120. [Google Scholar] [CrossRef]
- Wagle, P.; Zhang, Y.; Jin, C.; Xiao, X. Comparison of solar-induced chlorophyll fluorescence, light-use efficiency, and process-based GPP models in maize. Ecol. Appl. 2016, 26, 1211–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Li, J.; Yu, Q.; Ma, Y.; Tong, X.; Feng, Y.; Tong, Y. Impacts of diffuse radiation fraction on light use efficiency and gross primary production of winter wheat in the North China Plain. Agric. For. Meteorol. 2019, 275, 233–242. [Google Scholar] [CrossRef]
- Lasslop, G.; Reichstein, M.; Papale, D.; Richardson, A.; Arneth, A.; Barr, A.; Stoy, P.; Wohlfahrt, G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: Critical issues and global evaluation. Glob. Chang. Biol. 2010, 16, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.; Taylor, J.A. On the Temperature Dependence of Soil Respiration. Funct. Ecol. 1994, 8, 315–323. Available online: http://www.jstor.org/stable/2389824 (accessed on 12 November 2019).
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Sun, X.; Wen, X.; Zhou, Y.; Tian, J.; Yuan, G. Study on the processing method of nighttime CO2 eddy covariance flux data in ChinaFLUX. Sci. China Ser. D Earth Sci. 2006, 49, 36–46. [Google Scholar] [CrossRef]
- Saito, M.; Miyata, A.; Nagai, H.; Yamada, T. Seasonal variation of carbon dioxide exchange in rice paddy field in Japan. Agric. For. Meteorol. 2005, 135, 93–109. [Google Scholar] [CrossRef]
- Skiba, U.; Drewer, J.; Tang, Y.S.; van Dijk, N.; Helfter, C.; Nemitz, E.; Famulari, D.; Cape, J.N.; Jones, S.K.; Twigg, M.; et al. Biosphere-atmosphere exchange of reactive nitrogen and greenhouse gases at the NitroEurope core flux measurement sites: Measurement strategy and first data sets. Agric. Ecosyst. Environ. 2009, 133, 139–149. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B. Gross primary production and ecosystem respiration of irrigated and rainfed maize-soybean cropping systems over 8 years. Agric. For. Meteorol. 2012, 165, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Chen, J.; Gottgens, J.F.; Ouyang, Z.; John, R.; Czajkowski, K.; Becker, R. Net ecosystem methane and carbon dioxide exchanges in a Lake Erie coastal marsh and a nearby cropland. J. Geophys. Res.-Biogeosci. 2014, 119, 722–740. [Google Scholar] [CrossRef]
- Verma, S.B.; Dobermann, A.; Cassman, K.G.; Walters, D.T.; Knops, J.M.; Arkebauer, T.J.; Suyker, A.E.; Burba, G.G.; Amos, B.; Yang, H.; et al. Annual carbon dioxide exchange in irrigated and rainfed maize-based agroecosystems. Agric. For. Meteorol. 2005, 131, 77–96. [Google Scholar] [CrossRef] [Green Version]
- Wattenbach, M.; Sus, O.; Vuichard, N.; Lehuger, S.; Gottschalk, P.; Li, L.; Leip, A.; Williams, M.; Tomelleri, E.; Kutsch, W.L.; et al. The carbon balance of European croplands: A cross-site comparison of simulation models. Agric. Ecosyst. Environ. 2010, 139, 419–453. [Google Scholar] [CrossRef]
- Allison, V.J.; Miller, R.M.; Jastrow, J.D.; Matamala, R.; Zak, D.R. Changes in Soil Microbial Community Structure in a Tallgrass Prairie Chronosequence. Soil Sci. Soc. Am. J. 2005, 69, 1412–1421. [Google Scholar] [CrossRef] [Green Version]
- Monteith, J.L. Solar-radiation and productivity in tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747–766. Available online: http://www.jstor.org/stable/ (accessed on 7 June 2017). [CrossRef] [Green Version]
- Alton, P.B. The sensitivity of models of gross primary productivity to meteorological and leaf area forcing: A comparison between a Penman-Monteith ecophysiological approach and the MODIS Light-Use Efficiency algorithm. Agric. For. Meteorol. 2016, 218–219, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Boschetti, M.; Mauri, E.; Gadda, C.; Busetto, L.; Confalonieri, R.; Bocchi, S.; Brivio, P.A. Monitoring paddy rice crops through remote sensing: Productivity estimation by light use efficiency model. In Remote Sensing for Agriculture, Ecosystems, and Hydrology VI; International Society for Optics and Photonics: Bellingham, WA, USA, 2004; Volume 5568, pp. 46–56. [Google Scholar]
- Evrendilek, F.; Ben-Asher, J.; Aydin, M. Diurnal photosynthesis, water use efficiency and light use efficiency of wheat under Mediterranean field conditions. J. Environ. Biol. 2008, 29, 397–406. [Google Scholar] [PubMed]
- Evrendilek, F.; Ben-Asher, J.; Aydin, M.; Çelik, I. Spatial and temporal variations in diurnal CO2 fluxes of different Mediterranean ecosystems in Turkey. J. Environ. Monitor. 2005, 7, 151–157. [Google Scholar] [CrossRef]
- Xiao, X. Light absorption by leaf chlorophyll and maximum light use efficiency. IEEE Trans. Geosci. Remote Sens. 2006, 44, 1933–1935. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Miloslavina, Y.; Jahns, P.; Holzwarth, A.R. On the relationship between non-photochemical quenching and photoprotection of Photosystem II. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 760–769. [Google Scholar] [CrossRef] [Green Version]
- van Amerongen, H.; Chmeliov, J. Instantaneous switching between different modes of non-photochemical quenching in plants. Consequences for increasing biomass production. Biochim. Biophys. Acta-Bioenerg. 2020, 1861. [Google Scholar] [CrossRef] [PubMed]
- Ventre-Lespiaucq, A.; Flanagan, N.S.; Ospina-Calderón, N.H.; Delgado, J.A.; Escudero, A. Midday depression vs. Midday peak in diurnal light interception: Contrasting patterns at crown and leaf scales in a tropical evergreen tree. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Li, Q.; Dong, T.; Jia, K. Winter wheat biomass estimation using high temporal and spatial resolution satellite data combined with a light use efficiency model. Geocarto Int. 2015, 30, 258–269. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Arkebauer, T.J.; Suyker, A.E. Convergence of daily light use efficiency in irrigated and rainfed C3 and C4 crops. Remote Sens. Environ. 2018, 217, 30–37. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Liu, X. Directly estimating diurnal changes in GPP for C3 and C4 crops using far-red sun-induced chlorophyll fluorescence. Agric. For. Meteorol. 2017, 232, 1–9. [Google Scholar] [CrossRef]
- Xin, Q.; Broich, M.; Suyker, A.E.; Yu, L.; Gong, P. Multi-scale evaluation of light use efficiency in MODIS gross primary productivity for croplands in the Midwestern United States. Agric. For. Meteorol. 2015, 201, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Cai, W.; Nguy-Robertson, A.L.; Fang, H.; Suyker, A.E.; Chen, Y.; Dong, W.; Liu, S.; Zhang, H. Uncertainty in simulating gross primary production of cropland ecosystem from satellite-based models. Agric. For. Meteorol. 2015, 207, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.; Campos, I.; Plaza, C.; Calera, M.; Jiménez, N.; Bodas, V.; Calera, A. Water use efficiency and light use efficiency in garlic using a remote sensing-based approach. Agric. Water Manag. 2019, 219, 40–48. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Huang, C.; Qiao, N. An NDVI-based vegetation phenology is improved to be more consistent with photosynthesis dynamics through applying a light use efficiency model over boreal high-latitude forests. Remote Sens. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W.; Rivard, B.; Feng, J. Remote estimation of gross primary production in wheat using chlorophyll-related vegetation indices. Agric. For. Meteorol. 2009, 149, 1015–1021. [Google Scholar] [CrossRef]
- Baležentiene, L.; Mikša, O.; Baležentis, T.; Streimikiene, D. Seasonal net carbon exchange in rotation crops in the temperate climate of central Lithuania. Sustainability 2019, 11, 1966. [Google Scholar] [CrossRef] [Green Version]
- Schubert, P.; Lund, M.; Ström, L.; Eklundh, L. Impact of nutrients on peatland GPP estimations using MODIS time series data. Remote Sens. Environ. 2010, 114, 2137–2145. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.S. Evaluation of MODIS LAI, fAPAR and the relation between fAPAR and NDVI in a semi-arid environment using in situ measurements. Remote Sens. Environ. 2004, 91, 490–507. [Google Scholar] [CrossRef]
- Yebra, M.; Van Dijk, A.I.J.M.; Leuning, R.; Guerschman, J.P. Global vegetation gross primary production estimation using satellite-derived light-use efficiency and canopy conductance. Remote Sens. Environ. 2015, 163, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Li, X.; Zhou, S.; Yang, X.; Yu, K.; Wang, M.; Liu, S.; Wang, P.; Wu, X.; Wang, X.; et al. Quantifying plant transpiration and canopy conductance using eddy flux data: An underlying water use efficiency method. Agric. For. Meteorol. 2019, 271, 375–384. [Google Scholar] [CrossRef]
- Whitley, R.; Medlyn, B.; Zeppel, M.; Macinnis-Ng, C.; Eamus, D. Comparing the Penman-Monteith equation and a modified Jarvis-Stewart model with an artificial neural network to estimate stand-scale transpiration and canopy conductance. J. Hydrol. 2009, 373, 256–266. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Lee, X.; Wen, X.; Xiao, W. Evapotranspiration partitioning for three agro-ecosystems with contrasting moisture conditions: A comparison of an isotope method and a two-source model calculation. Agric. For. Meteorol. 2018, 252, 296–310. [Google Scholar] [CrossRef]
- Shuttleworth, W.J.; Wallace, J.S. Evaporation from sparse crops-an energy combination theory. Q. J. Roy. Meteor. Soc. 1985, 111, 839–855. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z. Modelling light use efficiency using vegetation index and land surface temperature from MODIS in Harvard Forest. Int. J. Remote Sens. 2012, 33, 2261–2276. [Google Scholar] [CrossRef]
- Chen, F.; Lin, A.; Zhu, H.; Niu, J. Quantifying climate change and ecological responses within the Yangtze River Basin, China. Sustainability 2018, 10, 3026. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, J.; Hu, Z. Comparison of carbon-use efficiency among different land-use patterns of the temperate steppe in the northern China pastoral farming ecotone. Sustainability 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W.; Noguchi, K.; Hikosaka, K.; Terashima, I. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances. Plant Physiol. 2010, 152, 388–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Song, C.; Sun, G.; Band, L.E.; Noormets, A.; Zhang, Q. Understanding moisture stress on light use efficiency across terrestrial ecosystems based on global flux and remote-sensing data. J. Geophys. Res.-Biogeosci. 2015, 120, 2053–2066. [Google Scholar] [CrossRef]
- Leuning, R.; Cleugh, H.A.; Zegelin, S.J.; Hughes, D. Carbon and water fluxes over a temperate Eucalyptus forest and a tropical wet/dry savanna in Australia: Measurements and comparison with MODIS remote sensing estimates. Agric. For. Meteorol. 2005, 129, 151–173. [Google Scholar] [CrossRef]
- Keenan, T.; García, R.; Friend, A.D.; Zaehle, S.; Gracia, C.; Sabate, S. Improved understanding of drought controls on seasonal variation in mediterranean forest canopy CO2 and water fluxes through combined in situ measurements and ecosystem modelling. Biogeosciences 2009, 6, 1423–1444. [Google Scholar] [CrossRef] [Green Version]
- Beer, C.; Ciais, P.; Reichstein, M.; Baldocchi, D.; Law, B.E.; Papale, D.; Soussana, J.F.; Ammann, C.; Buchmann, N.; Frank, D.; et al. Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Glob. Biogeochem. Cycles 2009, 23, 1–13. [Google Scholar] [CrossRef]
- Kanniah, K.D.; Beringer, J.; Hutley, L.B. Response of savanna gross primary productivity to interannual variability in rainfall: Results of a remote sensing based light use efficiency model. Prog. Phys. Geog. 2013, 37, 642–663. [Google Scholar] [CrossRef]
- Campoe, O.C.; Stape, J.L.; Albaugh, T.J.; Lee Allen, H.; Fox, T.R.; Rubilar, R.; Binkley, D. Fertilization and irrigation effects on tree level aboveground net primary production, light interception and light use efficiency in a loblolly pine plantation. For. Ecol. Manag. 2013, 288, 43–48. [Google Scholar] [CrossRef]
- Mishra, A.K.; Tripathi, P.; Pal, R.K.; Mishra, S.R. Light interception and radiation use efficiency of wheat varieties as influenced by number of irrigations. J. Agrometeorol. 2009, 11, 140–143. [Google Scholar]
- Camargo, D.C.; Montoya, F.; Moreno, M.A.; Ortega, J.F.; Córcoles, J.I. Impact of water deficit on light interception, radiation use efficiency and leaf area index in a potato crop (Solanum tuberosum L.). J. Agric. Sci. 2016, 154, 662–673. [Google Scholar] [CrossRef]
- Konings, A.G.; Gentine, P. Global variations in ecosystem-scale isohydricity. Glob. Chang. Biol. 2017, 23, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yu, G.; Fu, Y.; Sun, X.; Li, Y.; Shi, P.; Wang, Y.; Zheng, Z. Effects of vegetation control on ecosystem water use efficiency within and among four grassland ecosystems in China. Glob. Chang. Biol. 2008, 14, 1609–1619. [Google Scholar] [CrossRef]
- Puma, M.J.; Koster, R.D.; Cook, B.I. Phenological versus meteorological controls on land-atmosphere water and carbon fluxes. J. Geophys. Res.-Biogeosci. 2013, 118, 14–29. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Q.; Jiang, J.; Tang, Y. Calibration of Terra/MODIS gross primary production over an irrigated cropland on the North China Plain and an alpine meadow on the Tibetan Plateau. Glob. Chang. Biol. 2008, 14, 757–767. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W. Ecosystem Water Use Efficiency in the Three-North Region of China Based on Long-Term Satellite Data. Sustainability 2021, 13, 7977. [Google Scholar] [CrossRef]
- McCallum, I.; Franklin, O.; Moltchanova, E.; Merbold, L.; Schmullius, C.; Shvidenko, A.; Schepaschenko, D.; Fritz, S. Improved light and temperature responses for light-use-efficiency-based GPP models. Biogeosciences 2013, 10, 6577–6590. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.; Maselli, F.; Gilabert, M.A.; Chiesi, M.; Martínez, B.; Seufert, G. Assessment of MODIS imagery to track light-use efficiency in a water-limited Mediterranean pine forest. Remote Sens. Environ. 2012, 123, 359–367. [Google Scholar] [CrossRef]
- Reddy, T.Y.; Reddy, V.R.; Anbumozhi, V. Physiological responses of groundnut (Arachis hypogaea L.) to drought stress and its amelioration: A review. Plant Growth Regul. 2003, 41, 75–88. [Google Scholar] [CrossRef]
- Schull, M.A.; Anderson, M.C.; Houborg, R.; Gitelson, A.; Kustas, W.P. Thermal-based modeling of coupled carbon, water, and energy fluxes using nominal light use efficiencies constrained by leaf chlorophyll observations. Biogeosciences 2015, 12, 1511–1523. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Fu, Y.H.; Zhu, Z.; Liu, Y.; Liu, Z.; Huang, M.; Janssens, I.A.; Piao, S. Delayed autumn phenology in the Northern Hemisphere is related to change in both climate and spring phenology. Glob. Chang. Biol. 2016, 22, 3702–3711. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
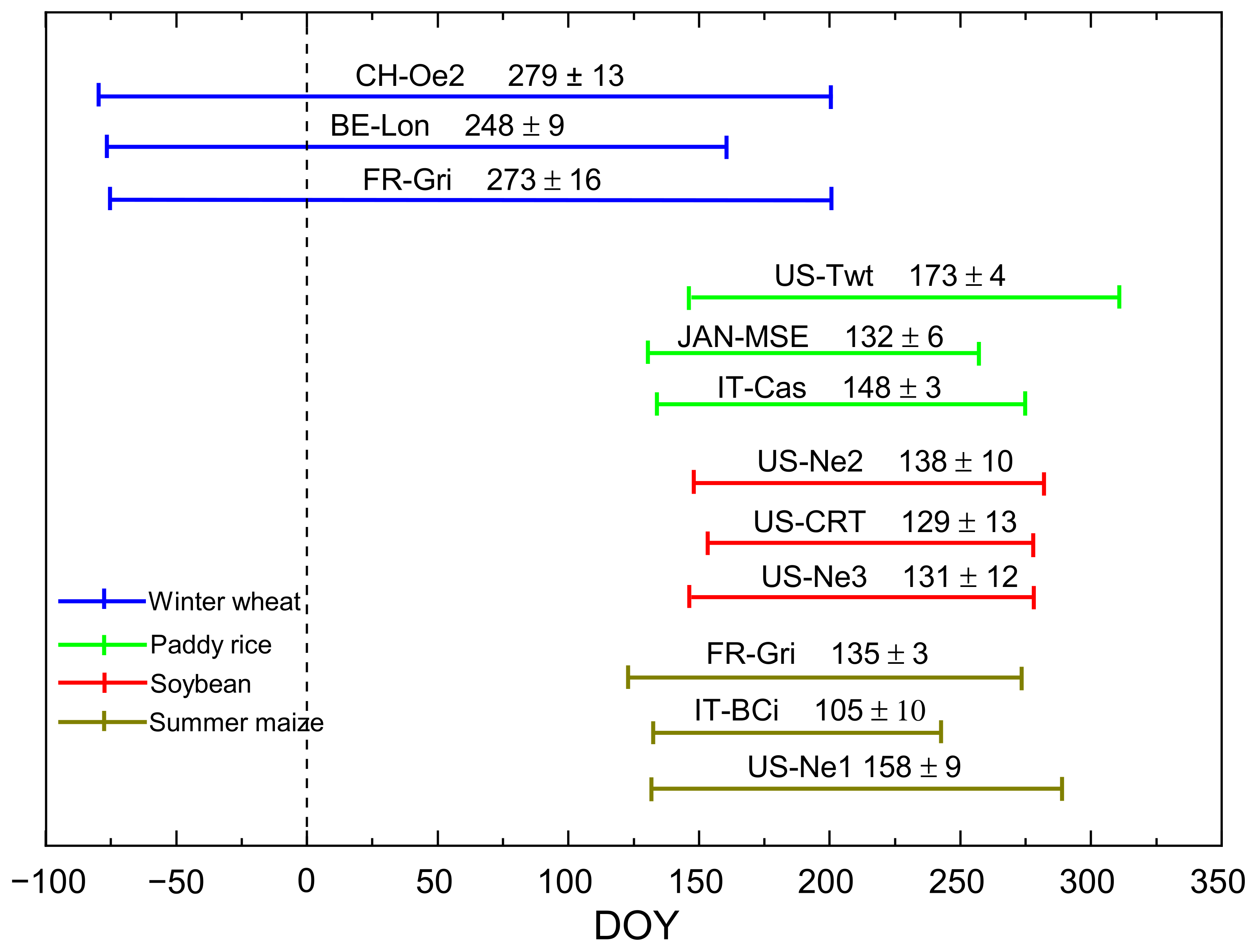
| Crop Type | Site ID | Country | Latitude (º) | Longitude (º) | MAT (°C) | MAP (mm) | Period | References |
|---|---|---|---|---|---|---|---|---|
| Paddy rice | US-Twt | USA | 38.11 | −121.6 | 15.6 | 421 | 2011–2013 | [12] |
| JAN-MSE | Japan | 36.05 | 140.02 | 13.7 | 1200 | 2004–2006 | [33] | |
| IT-Cas | Italy | 45.06 | 8.66 | 13.2 | 576 | 2007–2009 | [34] | |
| Soybean | US-Ne2 | USA | 41.16 | −96.47 | 10.1 | 789 | 2002/2004/2006 | [35] |
| US-CRT | USA | 41.62 | −83.34 | 10.1 | 849 | 2011–2012 | [36] | |
| US-Ne3 | USA | 41.17 | −96.43 | 10.1 | 784 | 2002/2004/2006 | [35] | |
| Summer maize | FR-Gri | France | 48.84 | 1.95 | 12.0 | 650 | 2005/2008/2011 | [34] |
| IT-BCi | Italy | 40.52 | 14.95 | 18.0 | 600 | 2005–2007 | [34] | |
| US-Ne1 | USA | 41.16 | −96.47 | 10.1 | 790 | 2005–2007 | [37] | |
| Winter wheat | CH-Oe2 | Switzerland | 47.28 | 7.73 | 9.8 | 1155 | 2006–2007/2008–2009/2010–2011 | [38] |
| BE-Lon | Belgium | 50.55 | 4.64 | 10.0 | 800 | 2004–2005/2006–2007/2008–2009 | [39] | |
| FR-Gri | France | 48.84 | 1.95 | 12.0 | 650 | 2005–2006/2009–2010/2011–2012 | [34] |
| Crop Type | GPP (g Cm−2 d−1) | ELUE (g C MJ−1) |
|---|---|---|
| Paddy rice | 7.72 ± 2.10 | 0.77 ± 0.24 |
| Soybean | 7.80 ± 1.80 | 0.80 ± 0.16 |
| Summer maize | 9.76 ± 0.80 | 0.92 ± 0.06 |
| Winter wheat | 4.74 ± 0.48 | 0.72 ± 0.06 |
| Crop Types | NDVI | gc | Ta | Rn | PAR | VPD | EF |
|---|---|---|---|---|---|---|---|
| Paddy rice | 0.85 ** | 0.24 | 0.47 ** | 0.39 ** | 0.28 * | −0.06 | −0.07 |
| Soybean | 0.91 ** | 0.68 ** | 0.27 | 0.37 * | 0.17 | −0.20 | 0.82 ** |
| Maize | 0.81 ** | 0.54 ** | 0.56 ** | 0.62 ** | 0.40 ** | −0.14 | 0.67 ** |
| Winter wheat | 0.76 ** | 0.40 ** | 0.44 ** | 0.59 ** | 0.62 ** | −0.28 * | 0.42 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Cui, N.; Huang, Y.; Hu, X.; Gong, D.; Wang, Y.; Lv, M.; Jiang, S. Investigating the Patterns and Controls of Ecosystem Light Use Efficiency with the Data from the Global Farmland Fluxdata Network. Sustainability 2021, 13, 12673. https://doi.org/10.3390/su132212673
Chen F, Cui N, Huang Y, Hu X, Gong D, Wang Y, Lv M, Jiang S. Investigating the Patterns and Controls of Ecosystem Light Use Efficiency with the Data from the Global Farmland Fluxdata Network. Sustainability. 2021; 13(22):12673. https://doi.org/10.3390/su132212673
Chicago/Turabian StyleChen, Fei, Ningbo Cui, Yaowei Huang, Xiaotao Hu, Daozhi Gong, Yaosheng Wang, Min Lv, and Shouzheng Jiang. 2021. "Investigating the Patterns and Controls of Ecosystem Light Use Efficiency with the Data from the Global Farmland Fluxdata Network" Sustainability 13, no. 22: 12673. https://doi.org/10.3390/su132212673
APA StyleChen, F., Cui, N., Huang, Y., Hu, X., Gong, D., Wang, Y., Lv, M., & Jiang, S. (2021). Investigating the Patterns and Controls of Ecosystem Light Use Efficiency with the Data from the Global Farmland Fluxdata Network. Sustainability, 13(22), 12673. https://doi.org/10.3390/su132212673