
Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective

 ,
,  and
and
Abstract
:1. Introduction
1.1. Global CO2 Emission Status
1.2. Carbon Sequestration Technologies
2. Physiological Mechanism of Carbon Sequestration in Algae
2.1. Photoautotrophic Metabolism
2.2. Heterotrophic Metabolism
2.3. Mixotrophic Metabolism
3. Carbon Concentration Mechanism (CCM) in Algae
3.1. C4 Pathways
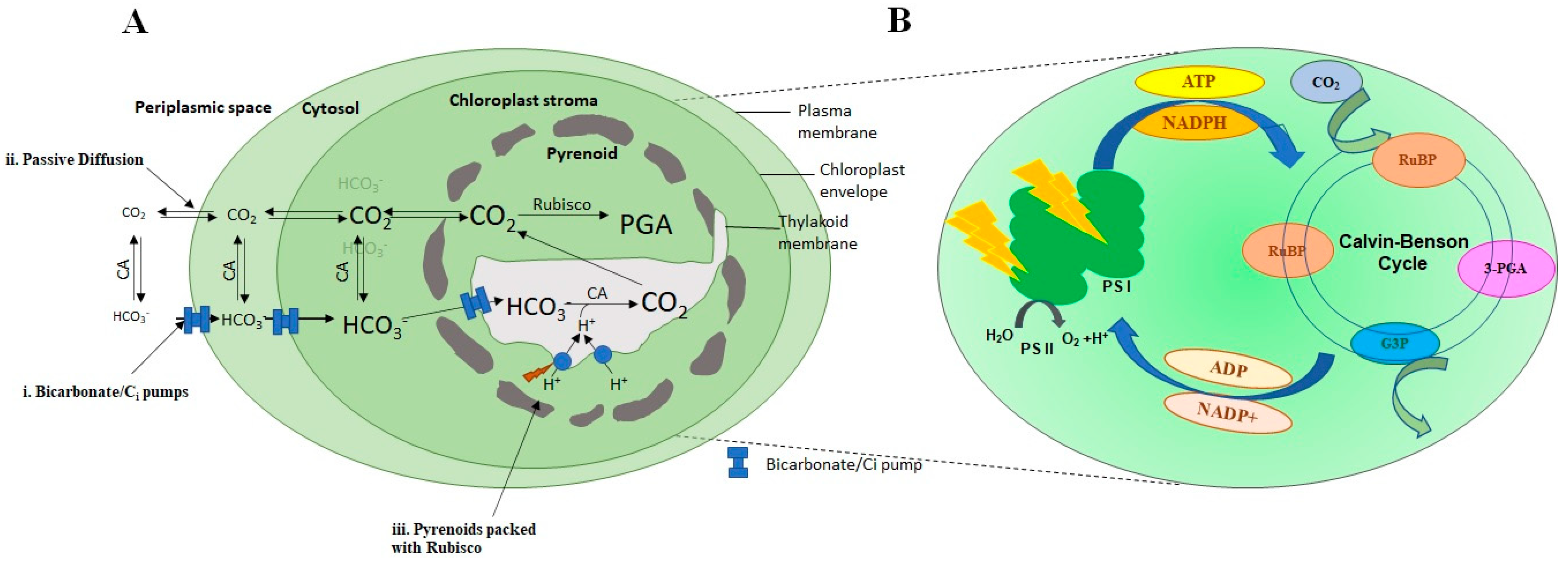
3.2. Inorganic Carbon (Ci) Transportation and Conversion Mechanism
3.3. Raise of CO2 Concentration around the Enzyme
4. Recent Development in Microalgae Carbon Sequestration
5. Factor Affecting Carbon Sequestration in Microalgae
5.1. CO2 Concentration
5.2. pH
5.3. Temperature
5.4. Irradiance
6. Techno-Economic Challenges with Microalgae
7. Future Prospective
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CCM | Carbon Concentration Mechanism |
| CCS | Carbon Capture and Storage |
| Sp. | Species |
| CO2 | Carbon Dioxide |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate Hydrogen |
| ATP | Adenosine triphosphate |
| CA | Carbonic Anhydrases |
References
- Hansen, J.; Sato, M.; Ruedy, R.; Lacis, A.; Oinas, V. Global warming in the twenty-first century: An alternative scenario. Proc. Natl. Acad. Sci. USA 2000, 97, 9875–9880. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Special Report on Emissions Scenarios: A Special Report of Working Group III of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2014; ISBN 92-9169-1135. [Google Scholar]
- Available online: https://www.epa.gov/ghgemissions (accessed on 20 November 2021).
- Lindsey, R. Climate Change: Atmospheric Carbon Dioxide; National Oceanic and Atmospheric Administration: Copenhagen, Denmark, 2020. [Google Scholar]
- Joint, I.; Doney, S.C.; Karl, D.M. Will ocean acidification affect marine microbes? ISME J. 2011, 5, 1–7. [Google Scholar] [CrossRef]
- Yang, X.; Liu, L.; Yin, Z.; Wang, X.; Wang, S.; Ye, Z. Quantifying photosynthetic performance of phytoplankton based on photosynthesis—Irradiance response models. Environ. Sci. Eur. 2020, 32, 24. [Google Scholar] [CrossRef] [Green Version]
- Shreyash, N.; Sonker, M.; Bajpai, S.; Tiwary, S.K.; Khan, M.A. The Review of Carbon Capture-Storage Technologies and Developing Fuel Cells for Enhancing Utilization. Energies 2021, 14, 4978. [Google Scholar] [CrossRef]
- Osman, A.I.; Hefny, M.; Maksoud, M.I.A.A.; Elgarahy, A.M.; Rooney, D.W. Recent advances in carbon capture storage and utilisation technologies: A review. Environ. Chem. Lett. 2021, 19, 797–849. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, Y.J.; Show Loke, P.; Suan, H.N.; Ching, J.J.; Chuan, T. Microalgae biorefinery: High-value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Faried, M.; Samer, M.; Abdelsalam, E.; Yousef, R.S.; Attia, Y.A.; Ali, A.S. Biodiesel production from microalgae: Processes, technologies and recent advancements. Renew. Sustain. Energy Rev. 2017, 79, 893–913. [Google Scholar] [CrossRef]
- Klinthong, W.; Yang, Y.; Huang, C.; Tan, C. A Review: Microalgae and Their Applications in CO2 Capture and Renewable Energy. Aerosol Air Qual. Res. 2015, 15, 712–742. [Google Scholar] [CrossRef]
- Camerini, F.; de Morais, M.G.; da Silva Vaz, B.; de Morais, E.G.; Costa, J.A.V. Biofixation of CO2 on a pilot scale: Scaling of the process for industrial application. Afr. J. Microbiol. Res. 2016, 10, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.M.; Mechery, J.; Paulose, S.V. Carbon dioxide capture strategies from flue gas using microalgae: A review. Environ. Sci. Pollut. Res. 2016, 16926–16940. [Google Scholar] [CrossRef]
- Zhu, X.; Rong, J.; Chen, H.; He, C.; Hu, W.; Wang, Q. An informatics-based analysis of developments to date and prospects for the application of microalgae in the biological sequestration of industrial flue gas. Appl. Microbiol. Biotechnol. 2016, 100, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-bermudez, S.P.; Garcia-perez, J.S.; Rittmann, B.E.; Parra-saldivar, R. Photosynthetic bioenergy utilizing CO2: An approach on flue gases utilization for third-generation biofuels. J. Clean. Prod. 2020, 98, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Lam, M.K.; Lee, K.T.; Mohamed, A.R. Current status and challenges on microalgae-based carbon capture. Int. J. Greenh. Gas Control 2012, 10, 456–469. [Google Scholar] [CrossRef]
- Khan, S.A.; Hussain, M.Z.; Prasad, S.; Banerjee, U.C. Prospects of biodiesel production from microalgae in India. Renew. Sustain. Energy Rev. 2009, 13, 2361–2372. [Google Scholar] [CrossRef]
- Barrett, J.; Girr, P.; Mackinder, L.C.M. Pyrenoids: CO2-fixing phase separated liquid organelles one phase two phase. BBA-Mol. Cell Res. 2021, 1868, 118949. [Google Scholar] [CrossRef]
- Wang, Y.; Stessman, D.J.; Spalding, M.H. The CO2 concentrating mechanism and photosynthetic carbon assimilation in limiting CO2: How Chlamydomonas works against the gradient. Plant J. 2015, 82, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.; Ho, S.; Chen, C.; Chang, J. CO2, NOx and SOx removal from flue gas via microalgae cultivation: A critical review. Biotechnol. J. 2015, 10, 829–839. [Google Scholar] [CrossRef]
- Larkum, A.W.D.; Ross, I.L.; Kruse, O.; Hankamer, B. Selection, breeding and engineering of microalgae for bioenergy and biofuel production. Trends Biotechnol. 2012, 30, 198–205. [Google Scholar] [CrossRef]
- Raheem, A.; Prinsen, P.; Vuppaladadiyam, A.K.; Zhao, M. A review on sustainable microalgae-based biofuel and bioenergy production: Recent developments. J. Clean. Prod. 2018, 181, 42–59. [Google Scholar] [CrossRef]
- Quiroz, C.E.; Peebles, C.; Bradley, T.H. Scalability of combining microalgae-based biofuels with wastewater facilities: A review. Algal Res. 2015, 9, 160–169. [Google Scholar] [CrossRef]
- Kothari, R.; Prasad, R.; Kumar, V.; Singh, D.P. Production of biodiesel from microalgae Chlamydomonas polypyrenoideum grown on dairy industry wastewater. Bioresour. Technol. 2013, 144, 499–503. [Google Scholar] [CrossRef]
- Bhola, V.; Swalaha, F.; Kumar, R.R.; Singh, M.; Bux, F. Overview of the potential of microalgae for CO2 sequestration. Int. J. Environ. Sci. Technol. 2014, 2103–2118. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, K.K.; Dutta, S.; Banerjee, I.; Pohrmen, C.B.; Kumar, V. Photosynthetic microalgae—Based carbon sequestration and generation of biomass in biorefinery approach for renewable biofuels for a cleaner environment. Biomass Conser. Biorefin. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Liu, Z. Near-real-time monitoring of global CO2 emissions reveals the effects of the COVID-19 pandemic. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Crippa, M.; Guizzardi, D.; Solazzo, E.; Muntean, M.; Schaaf, E.; Monforti-Ferrario, F.; Banja, M.; Olivier, J.G.J.; Grassi, G.; Rossi, S.; et al. GHG Emissions of All World Countries—2021 Report; EUR 30831 EN; Publications Office of the European Union: Luxembourg, 2021; ISBN 978-92-76-41546-6. [Google Scholar] [CrossRef]
- Crippa, M.; Solazzo, E.; Huang, G.; Guizzardi, D.; Koffi, E.; Muntean, M.; Schieberle, C.; Friedrich, R.; Janssens-Maenhout, G. High-resolution temporal profiles in the Emissions Database for Global Atmospheric Research. Sci. Data 2020, 7, 121. [Google Scholar] [CrossRef]
- IEA. World Energy Outlook; IEA: Paris, France, 2020; Available online: https://www.iea.org/reports/world-energy-outlook-2020 (accessed on 20 November 2021).
- Leung, D.Y.C.; Caramanna, G.; Maroto-valer, M.M. An overview of current status of carbon dioxide captures and storage technologies. Renew. Sustain. Energy Rev. 2014, 39, 426–443. [Google Scholar] [CrossRef] [Green Version]
- Jerndal, E.; Mattisson, T.; Lyngfelt, A.; Combustion, C.; After, O. Thermal analysis of chemical-looping combustion. Chem. Eng. Res. Des. 2006, 84, 795–806. [Google Scholar] [CrossRef]
- Pires, J.C.M.; Martins, F.G.; Simões, M. Carbon dioxide capture from flue gases using microalgae: Engineering aspects and biorefinery concept. Renew. Sustain. Energy Rev. 2012, 16, 3043–3053. [Google Scholar] [CrossRef]
- Suzuki, T.; Toriumi, M.; Sakemi, T.; Masui, N. Conceptual Design of CO2 Transportation System for CCS. Energy Procedia 2013, 37, 2989–2996. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Moroney, J.V.; Ynalvez, R.A. Proposed Carbon Dioxide Concentrating Mechanism in Chlamydomonas reinhardtii. Eukaryot. Cell 2007, 6, 1251–1259. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Shabnam, N.; Pardha-Saradhi, P. Immobilization on cotton cloth pieces is ideal for storage and conservation of microalgae. Algal Res. 2016, 20, 172–179. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://capitalenergy.biz/?p=16989 (accessed on 13 October 2021).
- Aci, G.; Matito-martos, I.; Sepúlveda, C.; Cintia, G.; Perez-carbajo, J.; Ania, C. Potential of CO2 capture from flue gases by physicochemical and biological methods: A comparative study. Chem. Eng. J. 2021, 417, 1–10. [Google Scholar]
- Vuppaladadiyam, A.K.; Yao, J.G.; Florin, N.; George, A.; Wang, X.; Labeeuw, L.; Jiang, Y.; Davis, R.W.; Abbas, A.; Ralph, P.; et al. Impact of Flue Gas Compounds on Microalgae and Mechanisms for Carbon Assimilation and Utilization. ChemSusChem 2018, 11, 334–355. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 Concentrating Mechanisms in Algae: Mechanisms, Environmental Modulation, and Evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spalding, M.H.; Biology, C. Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J. Exp. Bot. 2008, 59, 1463–1473. [Google Scholar] [CrossRef]
- Min, M.; Hu, B.; Zhou, W.; Li, Y.; Chen, P.; Ruan, R. Mutual influence of light and CO2 on carbon sequestration via cultivating mixotrophic alga Auxenochlorella protothecoides UMN280 in an organic carbon-rich wastewater. J. Appl. Phycol. 2012, 24, 1099–1105. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Silva, S.O.; Baptista, J.M.; Malcata, F.X. Light requirements in microalgal photobioreactors: An overview of biophotonic aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Bassaham, J.A.; Benson, A.A.; Kay, L.D.; Harris, A.Z.; Wilsson, A.T.C.M. The Path of Carbon in Photosynthesis. XXI. The Cyclic Regeneration of Carbon. J. Am. Chem. Soc. 1953, 76, 1760–1770. [Google Scholar] [CrossRef] [Green Version]
- Schloss, J.V. Oxygen toxicity from plants to people. Planta 2002, 216, 38–43. [Google Scholar] [CrossRef]
- Williams, P.J.B.; Laurens, L.M.L. Microalgae as biodiesel & biomass feedstocks: Review & analysis of the biochemistry, energetics & economics. Energy Environ. Sci. 2010, 3, 554–590. [Google Scholar] [CrossRef]
- Bowes, G.; Ogren, W.L.; Hageman, R.H. Phosphoclycol4te Production Catalyzed By Ribulose Diphosphate Carboxylase. Biochem. Biophys. Res. Commun. 1971, 45, 716–722. [Google Scholar] [CrossRef]
- Koning, R.E. Photorespiration. Plant Physiol. 1994. Available online: http://plantphys.info/plant_physiology/photoresp.shtml (accessed on 20 November 2021).
- Sayre, R. Microalgae: The Potential for Carbon Capture. Bioscience 2010, 60, 722–727. [Google Scholar] [CrossRef]
- Moroney, J.V.; Jungnick, N.; Dimario, R.J.; Longstreth, D.J. Photorespiration and carbon concentrating mechanisms: Two adaptations to high O2, low CO2 conditions. Photosynth. Res. 2013, 117, 121–131. [Google Scholar] [CrossRef]
- Tuchman, N.C.; Schollett, M.A.; Rier, S.T.; Geddes, P. Differential heterotrophic utilization of organic compounds by diatoms and bacteria under light and dark conditions. Hydrobiologia 2006, 561, 167–177. [Google Scholar] [CrossRef]
- Lowrey, J.; Armenta, R.E.; Brooks, M.S. Nutrient and media recycling in heterotrophic microalgae cultures. Appl. Microbiol. Biotechnol. 2016, 100, 1061–1075. [Google Scholar] [CrossRef]
- Morales-Sanchez, D.; Kyndt, J.; Martinez, A. Heterotrophic growth of microalgae: Metabolic aspects. World J. Microbiol. Biotechnol. 2015, 31, 1–9. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Chen, F. Biofuels from Algae, 1st ed.; Elsevier: San Diego, CA, USA, 2014; pp. 111–142. [Google Scholar]
- Ingram, L.O.; Baalen, C.; Van Calder, J.A. Role of Reduced Exogenous Organic Compounds in the Physiology of the Blue-Green Bacteria (Algae): Photoheterotrophic Growth of an “Autotrophic” Blue-Green Bacterium. J. Bacteriol. 1973, 114, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Lu, Y.; Chen, Y.; Wu, Q. Waste molasses alone displaces glucose-based medium for microalgal fermentation towards cost-saving biodiesel production. Bioresour. Technol. 2011, 102, 6487–6493. [Google Scholar] [CrossRef]
- Pleissner, D.; Chi, W.; Sun, Z.; Sze, C.; Lin, K. Biore source Technology Food waste as nutrient source in heterotrophic microalgae cultivation. Bioresour. Technol. 2013, 137, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K. Algal nutrition. Heterotrophic carbon nutrition. In Handbook of Microalgal Culture. Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Publishing: Oxford, UK, 2004; p. 116. [Google Scholar]
- Wang, J.; Yang, H.; Wang, F. Mixotrophic Cultivation of Microalgae for Biodiesel Production: Status and Prospects. Appl. Biochem. Biotechnol. 2014, 172, 3307–3329. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.V.; Devi, M.P. Bioresource Technology Salinity stress induced lipid synthesis to harness biodiesel during dual mode cultivation of mixotrophic microalgae. Bioresour. Technol. 2014, 165, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Min, M.; Zhou, W.; Li, Y.; Mohr, M.; Cheng, Y.; Lei, H.; Liu, Y.; Lin, X.; Chen, P.; et al. Influence of Exogenous CO2 on Biomass and Lipid Accumulation of Microalgae Auxenochlorella protothecoides Cultivated in Concentrated Municipal Wastewater. Appl. Biochem. Biotechnol. 2012, 166, 1661–1673. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hua, Q.; Shimizu, K. Energetics and carbon metabolism during growth of microalgal cells under photoautotrophic, mixotrophic and cyclic light-autotrophic/dark-heterotrophic conditions. Biochem. Eng. J. 2000, 6, 87–102. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-gonzález, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, A.; Chinnasamy, S.; Singh, M.; Das, K.C. Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl. Energy 2011, 88, 3425–3431. [Google Scholar] [CrossRef]
- Park, K.C.; Whitney, C.; Mcnichol, J.C.; Dickinson, K.E.; Macquarrie, S.; Skrupski, B.P.; Zou, J.; Wilson, K.E.; O’Leary, S.J.; McGinn, P.J. Mixotrophic and photoautotrophic cultivation of 14 microalgae isolates from Saskatchewan, Canada: Potential applications for wastewater remediation for biofuel production. J. Appl. Phycol. 2012, 24, 339–348. [Google Scholar] [CrossRef]
- Kong, W.; Song, H.; Hua, S.; Yang, H.; Yang, Q.; Xia, C. Enhancement of biomass and hydrocarbon productivities of Botryococcus braunii by mixotrophic cultivation and its application in brewery wastewater treatment. Afr. J. Microbiol. Res. 2012, 6, 1489–1496. [Google Scholar] [CrossRef]
- Patel, A.K.; Choi, Y.Y.; Sim, S.J. Emerging prospects of mixotrophic microalgae: Way forward to sustainable bioprocess for environmental remediation and cost-effective biofuels. Bioresour. Technol. 2020, 300, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-arita, C.; Sheehan, J.J.; Bradley, T.H. Life cycle net energy and greenhouse gas emissions of photosynthetic cyanobacterial biore fi neries: Challenges for industrial production of biofuels. Algal Res. 2017, 26, 445–452. [Google Scholar] [CrossRef]
- Durall, C.; Lindblad, P. Mechanisms of carbon fixation and engineering for increased carbon fixation in cyanobacteria. Algal Res. 2015, 11, 263–270. [Google Scholar] [CrossRef]
- Singh, S.K.; Sundaram, S.; Kishor, K. Carbon-Concentrating Mechanism. In Photosynthetic Microorganisms; SpringerBriefs in Materials; Springer: Cham, Switzerland, 2014. [Google Scholar] [CrossRef]
- Boatman, T.G.; Mangan, N.M.; Lawson, T.; Geider, R.J. Inorganic carbon and pH dependency of photosynthetic rates in Trichodesmium. J. Exp. Bot. 2018, 69, 3651–3660. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Hanson, D.; Price, G.D. Evolution and diversity of CO2 concentrating mechanisms in cyanobacteria. Funct. Plant Biol. 2002, 29, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Tabita, F.R.; Hanson, T.E.; Li, H.; Satagopan, S.; Singh, J.; Chan, S. Function, Structure, and Evolution of the RubisCO-Like Proteins and Their Rubisco Homologs. Microbiol. Mol. Biol. Rev. 2007, 71, 576–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, S.M.; Houtz, R.L.; Alonso, H. Advancing Our Understanding and Capacity to Engineer Nature’s CO2-Sequestering Enzyme, Rubisco. Plant Physiol. 2011, 155, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Sinetova, M.A.; Kupriyanova, E.V.; Markelova, A.G.; Allakhverdiev, S.I.; Pronina, N.A. Identification and functional role of the carbonic anhydrase Cah3 in thylakoid membranes of pyrenoid of Chlamydomonas reinhardtii. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. Inorganic carbon concentrating mechanisms in relation to the biology of algae. Photosynth. Res. 2003, 77, 155–171. [Google Scholar] [CrossRef]
- Reinfelder, J.R.; Kraepiel, A.M.L.; Morel, F.M.M. Unicellular C 4 photosynthesis in a marine diatom. Nature 2000, 407, 996–999. [Google Scholar] [CrossRef]
- Reinfelder, J.R.; Milligan, A.J.; Morel, M.M. The Role of the C4 Pathway in Carbon Accumulation and Fixation in a Marine Diatom. Plant Physiol. 2004, 135, 2106–2111. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.G.; Zwart, P.; Bagby, S.C.; Cai, F.; Chisholm, S.W.; Heinhorst, S.; Cannon, G.C.; Kerfeld, C.A. Identification and Structural Analysis of a Novel Carboxysome Shell Protein with Implications for Metabolite Transport. J. Mol. Biol. 2009, 392, 319–333. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wollman, A.J.M.; Huang, F.; Leake, M.C.; Liu, L. Single-Organelle Quanti fi cation Reveals Stoichiometric and Structural Variability of Carboxysomes Dependent on. Plant Cell 2019, 31, 1648–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turmo, A.; Gonzalez-Esquer, C.R.; Kerfeld, C.A. Carboxysomes: Metabolic modules for CO2 fixation. FEMS Microbiol. Lett. 2017, 364, fnx176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pronina, N.A.; Semenenko, V.E. Membrane-Bound Carbonic Anhydrase Takes Part in CO2 Concentration in Algae Cells. In Current Research in Photosynthesis; Baltscheffsky, M., Ed.; Springer: Dordrecht, The Netherlands, 1990. [Google Scholar] [CrossRef]
- Gutknecht, J.; Bisson, M.A.; Tosteson, F. Diffusion of Carbon Dioxide through Lipid Bilayer Membranes Effects of Carbonic Anhydrase, Bicarbonate, and Unstirred Layers. J. Gen. Physiol. 1977, 69, 779–794. [Google Scholar] [CrossRef] [Green Version]
- Yamano, T.; Fukuzawa, H. Carbon-concentrating mechanism in a green alga, Chlamydomonas reinhardtii, revealed by transcriptome analyses. J. Basic Microbiol. 2009, 49, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Bloom, A. Photorespiration: The Futile Cycle? Plants 2021, 10, 908. [Google Scholar] [CrossRef]
- Jungnick, N.; Ma, Y.; Mukherjee, B.; Cronan, J.C.; Moroney, J.V. The carbon concentrating mechanism in Chlamydomonas reinhardtii finding the missing pieces. Photosynth. Res. 2014, 121, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, K.; Cruz, J.A.; Kanazawa, A.; Kramer, D.M. The thylakoid proton motive force in vivo. Quantitative, non-invasive probes, energetics, and regulatory consequences of light-induced pmf. Biochim. Biophys. Acta 2007, 1767, 1233–1244. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Sing, C.; Walker, C.E.; Rai, A.K.; Prejean, C.I.; Yates, G. Thylakoid localized bestrophin-like proteins are essential for the CO2 concentrating mechanism of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 16915–16920. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.T.; Whittaker, C.; Griffiths, H. The algal pyrenoid: Key unanswered questions. J. Exp. 2017, 68, 3739–3749. [Google Scholar] [CrossRef] [Green Version]
- Rochaix, J. The Pyrenoid: An Overlooked Organelle Comes out of Age. Cell 2017, 171, 28–29. [Google Scholar] [CrossRef] [Green Version]
- Liberton, M.; Ii, J.R.A.; Berg, R.H.; Pakrasi, H.B. Insights into the complex 3-D architecture of thylakoid membranes in unicellular cyanobacterium Cyanothece sp. ATCC 51142. Plant Signl. Behav 2011, 6, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Jonikas, M.C. The pyrenoid. Curr. Biol. 2020, 30, R451–R520. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.; Leitao, N.; Orr, D.J.; Meyer, M.T.; Carmo-silva, E.; Griffiths, H.; Smith, A.M.; McCormick, A.J. Rubisco small subunits from the unicellular green alga Chlamydomonas complement Rubisco-deficient mutants of Arabidopsis. New Phytol. 2017, 214, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedroni, P.; Davision, J.; Beckert, H.; Bergman, P. A Proposal to Establish an International Network on Biofixation of CO2 and Greenhouse Gas Abatement with Microalgae. J. Energy Environ. Res. 2001, 1, 136–150. [Google Scholar]
- Engel, B.D.; Schaffer, M.; Cuellar, L.K.; Villa, E.; Plitzko, J.M.; Baumeister, W. Native architecture of the Chlamydomonas chloroplast revealed by in situ cryo-electron tomography. elife 2015, 4, 1–29. [Google Scholar] [CrossRef]
- Zhang, C.; Li, W.; Shi, Y.; Li, Y.; Huang, J.; Li, H. A new technology of CO2 supplementary for microalgae cultivation on large scale—A spraying absorption tower coupled with an outdoor open runway pond. Bioresour. Technol. 2016, 209, 351–359. [Google Scholar] [CrossRef]
- Lee, J.; Kim, D.; Lee, J.; Park, S.; Koh, J.; Cho, H.; Kim, S.W. Effects of SO2 and NO on growth of Chlorella sp. KR-1. Bioresour. Technol. 2002, 82, 1–4. [Google Scholar] [CrossRef]
- Vunjak-novakovic, G.; Kim, Y.; Wu, X.; Berzin, I. Air-Lift Bioreactors for Algal Growth on Flue Gas: Mathematical Modeling and Pilot-Plant Studies. Ind. Eng. Chem. Res. 2005, 44, 6154–6163. [Google Scholar] [CrossRef]
- Hoyos, B.S.; Deiver, A.G.; Miranda, A.M.; Alex, A.S.; Vargas, G.J. Improving microalgal biomass production with industrial CO2 for bio-oil obtention by hydrothermal liquefaction. Fuel 2021, 302, 1–6. [Google Scholar]
- Ho, S.; Chen, C.; Lee, D.; Chang, J. Perspectives on microalgal CO2-emission mitigation systems—A review. Biotechnol. Adv. 2020, 29, 189–198. [Google Scholar] [CrossRef]
- Kassim, M.A.; Meng, T.K. Carbon dioxide (CO2) biofixation by microalgae and its potential for biorefinery and biofuel production. Sci. Total Environ. 2017, 585, 1121–1129. [Google Scholar] [CrossRef]
- Sadeghizadeh, A.; Farhad, F.; Moghaddasi, L.; Rahimi, R. CO2 capture from air by Chlorella vulgaris microalgae in an airlift photobioreactor. Bioresour. Technol. 2017, 243, 441–447. [Google Scholar] [CrossRef]
- Kuo, C.; Jian, J.; Lin, T.; Chang, Y.; Wan, X.; Lai, J.; Chang, J.S.; Lin, C.S. Simultaneous microalgal biomass production and CO2 fixation by cultivating Chlorella sp. G.D. with aquaculture wastewater and boiler flue gas. Bioresour. Technol. 2016, 221, 241–250. [Google Scholar] [CrossRef]
- Zhao, B.; Su, Y. Process effect of microalgal-carbon dioxide fixation and biomass production: A review. Renew. Sustain. Energy Rev. 2014, 31, 121–132. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.; Chen, C.; Chen, C.N.; Liu, C.; Ng, I.; Jing, K.J.; Yang, S.C.; Chen, C.H.; Chang, J.S.; et al. Simultaneous enhancement of CO2 fixation and lutein production with thermo-tolerant Desmodesmus sp. F51 using a repeated fed-batch cultivation strategy. Biochem. Eng. J. 2014, 86, 33–40. [Google Scholar] [CrossRef]
- Anjos, M.; Fernandes, B.D.; Vicente, A.A.; Teixeira, J.A.; Dragone, G. Optimization of CO2 bio-mitigation by Chlorella vulgaris. Bioresour. Technol. 2013, 139, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Han, W.; Li, P.; Miao, X.; Zhong, J. CO2 biofixation and fatty acid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in response to different CO2 levels. Bioresour. Technol. 2011, 102, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Zhang, Y.; Xiong, K.; Zhang, Z. Effect of cultivation mode on microalgal growth and CO2. Chem. Eng. Res. Des. 2011, 89, 1758–1762. [Google Scholar] [CrossRef]
- Chiang, C.; Lee, C.; Chen, P. Utilization of the cyanobacteria Anabaena sp. CH1 in biological carbon dioxide mitigation processes. Bioresour. Technol. 2011, 102, 5400–5405. [Google Scholar] [CrossRef]
- Basu, S.; Sarma, A.; Mohanty, K.; Ghoshal, A.K. Enhanced CO2 sequestration by a novel microalga: Scenedesmus obliquus SA1 isolated from bio-diversity hotspot region of Assam, India. Bioresour. Technol. 2013, 143, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, Z.; Zeng, R.; Yang, G.; Chang, X.; Yan, J.; Hou, Y.L. Microalgae Capture of CO2 from Actual Flue Gas Discharged from a Combustion Chamber. Ind. Eng. Chem. Res. 2011, 50, 6496–6502. [Google Scholar] [CrossRef]
- Ramanan, R.; Kannan, K.; Deshkar, A.; Yadav, R.; Chakrabarti, T. Enhanced algal CO2 sequestration through calcite deposition by Chlorella sp. and Spirulina platensis in a mini-raceway pond. Bioresour. Technol. 2010, 101, 2616–2622. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.; Jun, S.; Lee, J.; Ahn, C.; Oh, H. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour. Technol. 2010, 101, S71–S74. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, A.L.; Dennis, J.S.; Howe, C.J.; Scott, S.A.; Smith, A.G. Influence of nitrogen-limitation regime on the production by Chlorella vulgaris of lipids for biodiesel feedstocks Influence of nitrogen-limitation regime on the production by Chlorella vulgaris of lipids for biodiesel feedstocks. Biofuels 2014, 1, 47–58. [Google Scholar] [CrossRef]
- Sydney, E.B.; Sturm, W.; de Carvalho, J.C.; Thomaz-soccol, V.; Larroche, C.; Pandey, A.; Soccol, C.R. Potential carbon dioxide fixation by industrially important microalgae. Bioresour. Technol. 2010, 101, 5892–5896. [Google Scholar] [CrossRef]
- Chiu, S.Y.; Tsai, M.T.; Kao, C.Y.; Ong, S.C.; Lin, C.S. The air-lift photobioreactors with flow patterning for high-density cultures of microalgae and carbon dioxide removal. Eng. Life Sci. 2009, 9, 254–260. [Google Scholar] [CrossRef]
- Ryu, H.J.; Oh, K.K.; Kim, Y.S. Optimization of the influential factors for the improvement of CO2 utilization efficiency and CO2 mass transfer rate. J. Ind. Eng. Chem. 2009, 15, 471–475. [Google Scholar] [CrossRef]
- Fan, L.; Zhang, Y.; Zhang, L.; Chen, H. Evaluation of a membrane-sparged helical tubular photobioreactor for carbon dioxide biofixation by Chlorella vulgaris. J. Membr. Sci. 2008, 325, 336–345. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, L.; Chen, H.; Gao, C. Carbon dioxide removal from air by microalgae cultured in a membrane-photobioreactor. Sep. Purif. Technol. 2006, 50, 324–329. [Google Scholar] [CrossRef]
- Jin, H.; Lim, B.; Lee, K. Influence of Nitrate Feeding on Carbon Dioxide Fixation by Microalgae. J. Environ. Sci. Health Part A Toxic/Hazard. Subst. Environ. Eng. 2007, 41, 37–41. [Google Scholar] [CrossRef]
- Yue, L.; Chen, W. Isolation and determination of cultural characteristics of a new highly CO2 tolerant fresh water microalgae. Energy Convers. Manag. 2005, 46, 1868–1876. [Google Scholar] [CrossRef]
- Yun, Y.; Lee, S.B.; Park, J.M.; Lee, C.; Yang, J. Carbon Dioxide Fixation by Algal Cultivation Using Wastewater Nutrients. J. Chem. Technol. Biotechnol. 1997, 69, 451–455. [Google Scholar] [CrossRef]
- Solovechenko, A.; Khoizin-Goldberg, I. High-CO2 tolerance in microalgae: Possible mechanisms and implications for biotechnology and bioremediation. Biotechnol. Lett. 2013, 35, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Singh, P. Effect of CO2 concentration on algal growth: A review. Renew. Sustain. Energy Rev. 2014, 38, 172–179. [Google Scholar] [CrossRef]
- Azov, Y. Effect of pH Inorganic Carbon Uptake in Algal Cultures. Appl. Environ. Microbiol. 1982, 43, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.; Wang, Z.; Wen, X.; Ding, Y.; Hou, X.; Geng, Y.; Li, Y. Effects of carbon concentration, pH, and bubbling depth on carbon dioxide absorption ratio in microalgae medium. Environ. Sci. Pollut. Res. 2019, 26, 32902–32910. [Google Scholar] [CrossRef]
- Al Jabri, H.; Taleb, A.; Touchard, R.; Saadaoui, I.; Goetz, V.; Pruvost, J. Cultivating Microalgae in Desert Conditions: Evaluation of the Effect of Light-Temperature Summer Conditions on the Growth and Metabolism of Nannochloropsis QU130. Appl. Sci. 2021, 11, 3799. [Google Scholar] [CrossRef]
- Ördög, V.; Stirk, W.A.; Bálint, P.; Aremu, A.O.; Okem, A.; Lovász, C.; Molnár, Z.; van Staden, J. Effect of temperature and nitrogen concentration on lipid productivity and fatty acid composition in three Chlorella strains. Algal Res. 2016, 16, 141–149. [Google Scholar] [CrossRef]
- Xu, N.J.; Zhang, X.C. Effect of temperature, light intensity and pH on the growth and fatty acid compositions of Ellipsoidion sp. J. Ocean Univ. Qingdao 2001, 31, 541–547. [Google Scholar]
- Pueyo, J.; Alfonso, M.; Andre, C.; Picorel, R. Increased tolerance to thermal inactivation of oxygen evolution in spinach Photosystem II membranes by substitution of the extrinsic 33-kDa protein by its homologue from a thermophilic cyanobacterium. Biochim. Biophys. Acta 2002, 1554, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Lindberg, P.; Lindblad, P. Sustainable Energy & Fuels enhanced growth and productivity. Sustain. Energy Fuels 2018, 2, 2583–2600. [Google Scholar] [CrossRef] [Green Version]
- Aro, E.; Virgin, I.; Andersson, B. Photoinhibition of Photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Pniewski, F.; Piasecka, I. Photoacclimation to constant and changing light conditions in a benthic diatom. Fron. Mar. Sci. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- De Oliveira, C.Y.B.; Viegas, T.L.; Fernanda, M.; Fracalossi, D.M.; Lopes, R.G.; Derner, R.B. Effect of trace metals on growth performance and accumulation of lipids, proteins, and carbohydrates on the green microalga Scenedesmus obliquus. Aquac. Int. 2020, 28, 1435–1444. [Google Scholar] [CrossRef]
- Garcia-Chamcho, F.; Sanchez-Miron, A.; Molina-Grima, E.; Chamcho-Ruvio, F.; Merchuck, J.C. A mechanistic model of photosynthesis in microalgae including photoacclimation dynamics. J. Theor. Biol. 2012, 304, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Shigeoka, S. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis 1. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Krzeminska, I.; Pawlik-Skowrońska, B.; Trzcinska, M.; Tys, J. Influence of photoperiods on the growth rate and biomass productivity of green microalgae. Bioprocess Biosyst. Eng. 2014, 37, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Ramanna, L.; Rawat, I.; Bux, F. Light enhancement strategies improve microalgal biomass productivity. Renew. Sustain. Energy Rev. 2017, 80, 765–773. [Google Scholar] [CrossRef]
- Slade, R.; Bauen, A. Micro-algae cultivation for biofuels: Cost, energy balance, environmental impacts and future prospects. Biomass Bioenergy 2013, 53, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Singh, U.B.; Ahluwalia, A.S. Microalgae: A promising tool for carbon sequestration. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 73–95. [Google Scholar] [CrossRef]
- Quiroz-arita, C.; Blaylock, M.L.; Gharagozloo, P.E.; Bradley, T.H.; Dempster, T.; Mcgowen, J.; Davis, R.W. Bioresource Technology Reports A dynamic thermal algal growth model for pilot-scale open-channel raceways. Bioresour. Technol. Rep. 2020, 10, 100405. [Google Scholar] [CrossRef]
- Jorquera, O.; Kiperstok, A.; Sales, E.A.; Embiruçu, M.; Ghirardi, M.L. Comparative energy life-cycle analyses of microalgal biomass production in open ponds and photobioreactors. Bioresour. Technol. 2010, 101, 1406–1413. [Google Scholar] [CrossRef]
- Yadav, G.; Dubey, B.K.; Sen, R. A comparative life cycle assessment of microalgae production by CO2 sequestration from flue gas in outdoor raceway ponds under batch and semi-continuous regime. J. Clean. Prod. 2020, 258, 120703. [Google Scholar] [CrossRef]
- Stewart, C.; Hessami, M. A study of methods of carbon dioxide capture and sequestration—The sustainability of a photosynthetic bioreactor approach. Energy Convers. Manag. 2005, 46, 403–420. [Google Scholar] [CrossRef]
- Stepan, D.J.; Shockey, R.E.; Moe, T.A.; Dorn, R. Carbon Dioxide Sequestering Using Microalgal Sustems; University of North Dakota: Grand Forks, ND, USA, 1998. [Google Scholar]
- Cheng, Y.W.; Lim, J.S.M.; Chong, C.C.; Lam, M.K.; Lim, J.W.; Tan, I.S.; Foo, H.C.Y.; Show, P.L.; Lim, S. Unravelling CO2 capture performance of microalgae cultivation and other technologies via comparative carbon balance analysis. J. Env. Chem. Eng. 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Picardo, M.C.; Luiz, J.; Medeiros, D.; Ofélia de Queiroz, F.A.; Chaloub, R.M. Effects of CO2 enrichment and nutrients supply intermittency on batch cultures of Isochrysis galbana. Bioresour. Technol. 2013, 143, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Quiroz, C.; Yilmaz, Ö.; Barlak, S.; Catton, K.B.; Quinn, J.C.; Bradley, T.H. Bioresource Technology A geographical assessment of vegetation carbon stocks and greenhouse gas emissions on potential microalgae-based biofuel facilities in the United States. Bioresour. Technol. 2016, 221, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Daneshvar, E.; Wicker, R.J.; Show, P.; Bhatnagar, A. Biologically-mediated carbon capture and utilization by microalgae towards sustainable CO2 biofixation and biomass valorization—A review. Chem. Eng. J. 2021, 427, 1–15. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, J.; Chen, P.; Ji, C.; Kang, Q.; Lu, B.; Li, K.; Liu, J.; Ruan, R. Bio-mitigation of carbon dioxide using microalgal systems: Advances and perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1163–1175. [Google Scholar] [CrossRef]
- Mondal, M.; Goswami, S.; Ghosh, A.; Oinam, G. Production of biodiesel from microalgae through biological carbon capture: A review. 3 Biotech 2017, 7, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.A.; Cockell, C.S.; De La Rocha, C.L. The evolution of inorganic carbon concentrating mechanisms in photosynthesis. Philos. Trans. R. Soc. B 2008, 363, 2641–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gayathri, R.; Mahboob, S.; Govindarajan, M.; Al-ghanim, K.A.; Ahmed, Z.; Al-mulhm, N.; Vodovnik, M.; Vijayalakshmi, S. A review on biological carbon sequestration: A sustainable solution for a cleaner air environment, less pollution and lower health risks. J. King Saud Univ.-Sci. 2021, 33, 101282. [Google Scholar] [CrossRef]
- Cheah, Y.W.; Show Loke, P.; Chang, J.; Ling, C.T.; Juan, C.J. Biosequestration of atmospheric CO2 and flue gas-containing CO2 by microalgae. Bioresour. Technol. 2015, 184, 190–201. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S NO. | Microalgae | CO2 Tolerance Capacity (%) | CO2 Assimilation Rate (g/L/d) | CO2 Assimilation Efficiency (g/L/d) | Cultivation Conditions | Cultivation System | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | T °C | CO2(%) | Light Intensity | Culture Medium | |||||||
| 1. | Chlorella sp. | 40 | 0.097 | - | 7.5–9 | 30 | 15 | 450 # | - | - | [104] |
| 2. | Chlorella vulgaris | 18 | - | 76 | 7.2 | 30 | 30 | 1800 * | f/2 | APBRa | [105] |
| 3. | Chlorella sp. | 40 | 2.33 | - | 6.3–9 | 26 | 26 | 100 $ | Modified freshwater medium | FPBR | [106] |
| 4. | Chlorella sp. | 40 | 0.510 | - | 8.2 | 18 | 10–20 | 84 # | - | BCPBR | [107] |
| 5. | Desmodesmus sp. | 100 | 1.58 | - | - | 30 | 30 | 60 # | 3N-BBM | FBC | [108] |
| 6. | Chlorella vulgaris | 18 | 2.22 | - | - | 30 | - | 70 # | BG11 | BCPB | [109] |
| 7. | Chlorella pyrenoidosa Scendesmus obliquus | 10 10 | 0.26 0.28 | - - | 7 | 25 | 10 | 180 # | BG11 | EF | [110] |
| 8. | Chlorella sp. | - | 0.25 1.7 | 8 | 18 | 0.03 | 6000 * | f/2 and AFW | BPR | [111] | |
| 9. | Anabaena sp. | 10 | 1.01 | 67–79 | 20–25 | 5–15 | 127–250 # | BG11 | BPR | [112] | |
| 10. | Scenedesmus obliquus | 18 | 0.252 | - | 7 | 25 | 13.8 | 5496 * | f/2 | EF | [113] |
| 11. | Scenedesmus obliquus | 18 | - | 67 | - | 26 | 26–28 | 12,000 * | Soil extract | APBR | [114] |
| 12. | Chlorella sp. | 40 | - | 46 | 10 | 30 | 10 | 30 # | - | LSF | [115] |
| 13. | Scenedesmus obliquus | 18 | - | 40.2 | - | 25 | 10 | 12,000 * | - | APBR | [114] |
| 14. | Botryococcus braunii | 10 | - | - | - | 25 | 5.5 | 150 # | Chu 13 | - | [116] |
| 15. | Chlorella vulgaris | 18 | 0.522 | - | 7.2 | 22 | 22 | 165 # | 3N-BBM | CF | [117] |
| 16. | Chlorella vulgaris | 18 | 0.251 | - | 6.0 | 30 | 30 | 3500 * | FM | [118] | |
| 17. | Chlorella sp. | 10 | - | - | - | 26 | 10 | 300 # | AFW | BCPBR | [119] |
| 18. | Chlorella sp. | 5 | 0.35 | - | 7.18 | - | 5 | 100 # | BG11 | VTPBR | [120] |
| 19. | Chlorella vulgaris | 18 | 2.664 | - | 7.02–8.2 | 25 | 25 | 3600 * | Synthetic Sea Salt | PCPB | [121] |
| 20. | Chlorella vulgaris | 1 | 6.24 | - | 8.5 | 27 | 0.2 | 75 $ | - | MPBR | [122] |
| 21. | Chlorella | 15 | 0.46 | - | 8 | 27 | 0.2 | 200 # | MA | CF | [123] |
| 22. | Chlorella | 10 | - | 0.57 | 6 | 25 | 10 | MBM | BCPBR | [124] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13061. https://doi.org/10.3390/su132313061
Prasad R, Gupta SK, Shabnam N, Oliveira CYB, Nema AK, Ansari FA, Bux F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability. 2021; 13(23):13061. https://doi.org/10.3390/su132313061
Chicago/Turabian StylePrasad, Ravindra, Sanjay Kumar Gupta, Nisha Shabnam, Carlos Yure B. Oliveira, Arvind Kumar Nema, Faiz Ahmad Ansari, and Faizal Bux. 2021. "Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective" Sustainability 13, no. 23: 13061. https://doi.org/10.3390/su132313061
APA StylePrasad, R., Gupta, S. K., Shabnam, N., Oliveira, C. Y. B., Nema, A. K., Ansari, F. A., & Bux, F. (2021). Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability, 13(23), 13061. https://doi.org/10.3390/su132313061