
Slow-Release Urea as a Sustainable Alternative to Soybean Meal in Ruminant Nutrition

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1: In Vitro Evaluation to Determine the Effect of Slow-Release Urea on Rumen Fermentation
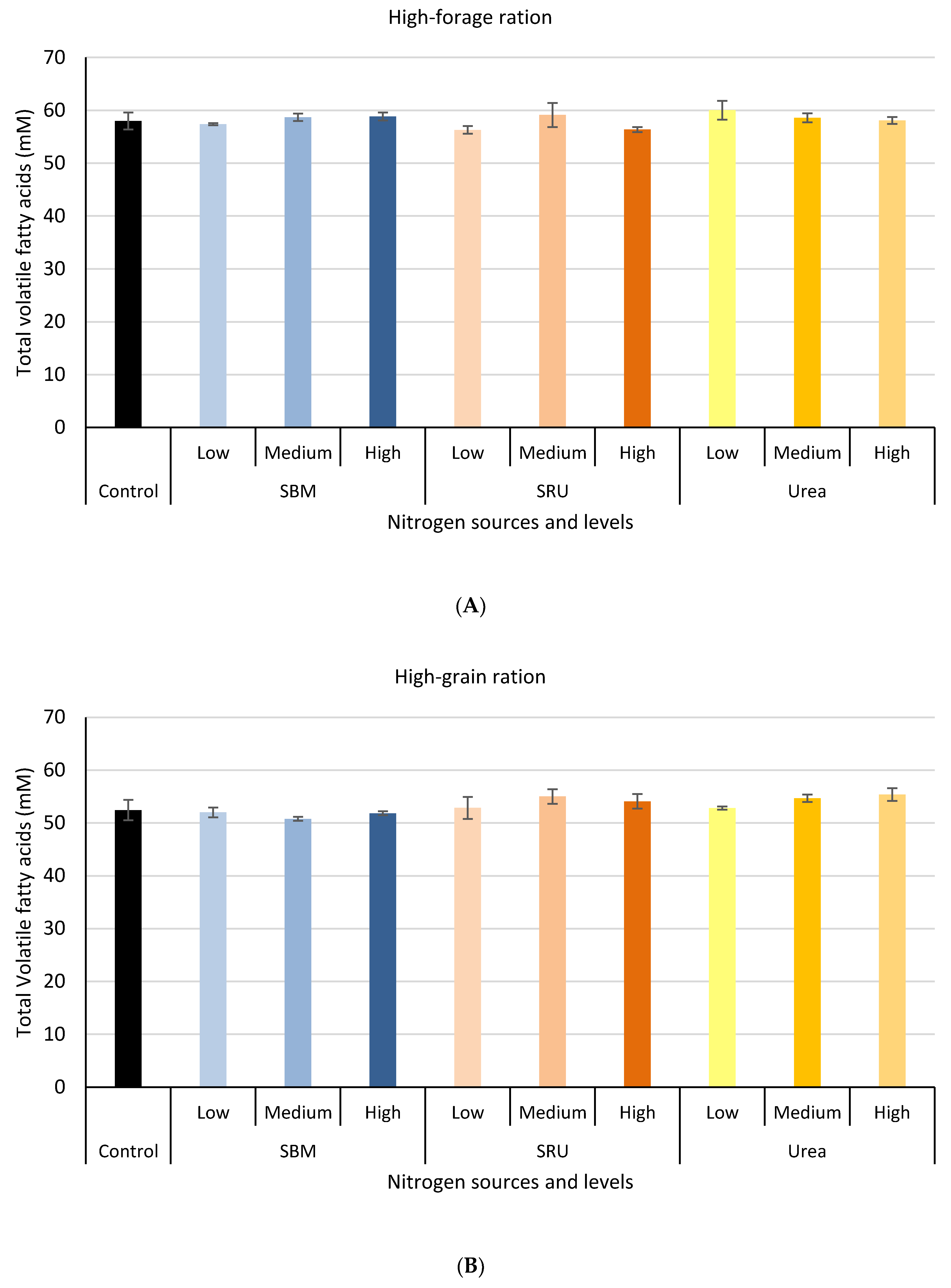
2.1.1. Diets, Treatments, and Doses
2.1.2. Rumen Fermentation Simulation
2.1.3. Analytical Procedures
2.2. Experiment 2: In Situ Evaluation to Determine the Effect of Slow-Release Urea on Feed Degradability in the Rumen
2.2.1. Diets, Treatments and In Situ Incubation
2.2.2. Analysis of Dry Matter and Nitrogen Degradation
2.3. Experiment 3: In Vivo Trial to Evaluate Feed Carbon Footprint and Toxicity Potential of Slow-Release Urea in Cattle
2.3.1. Animals, Diets and Experimental Design
2.3.2. Chemical Analysis
2.3.3. Calculations
2.4. Statistical Analysis
3. Results
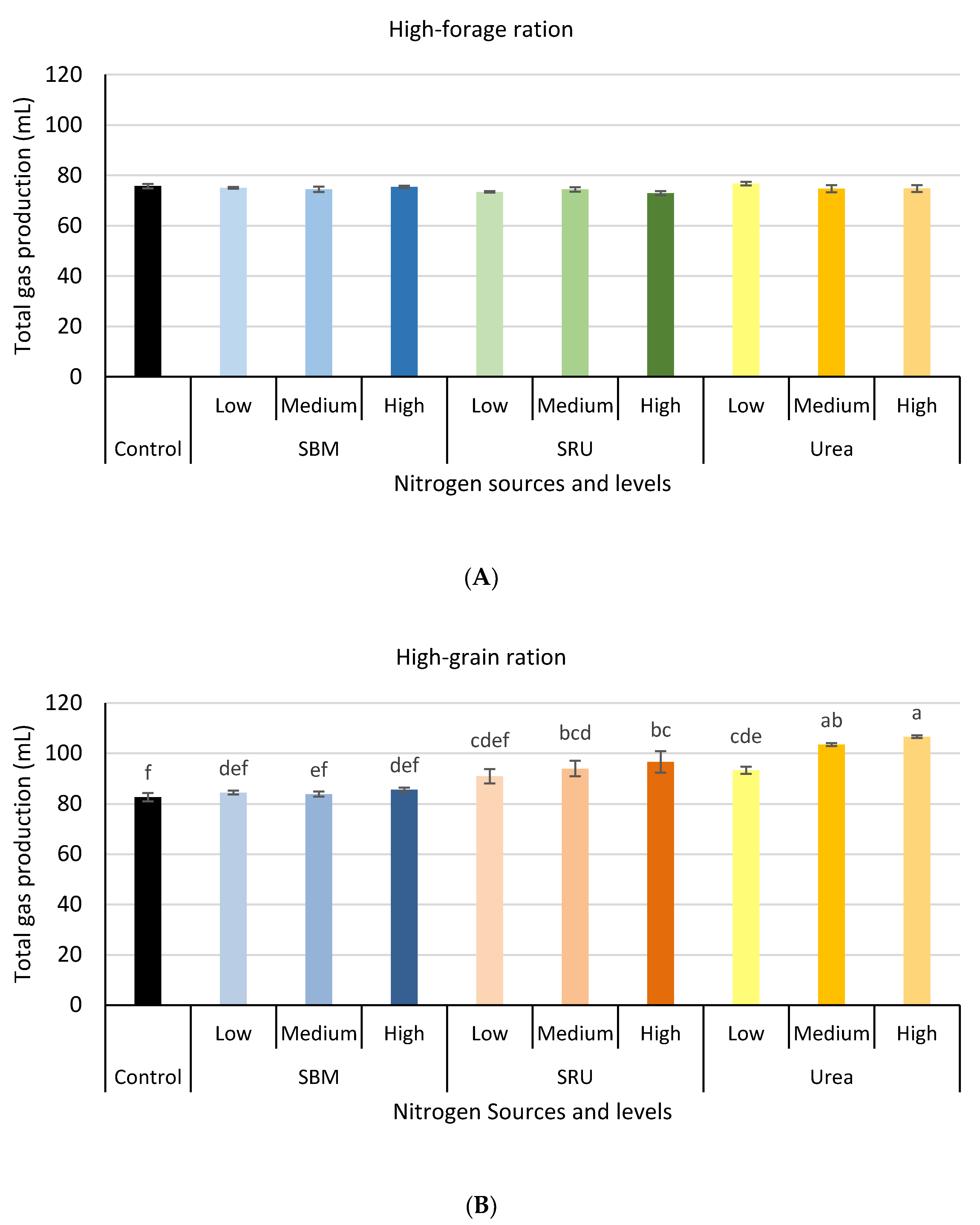
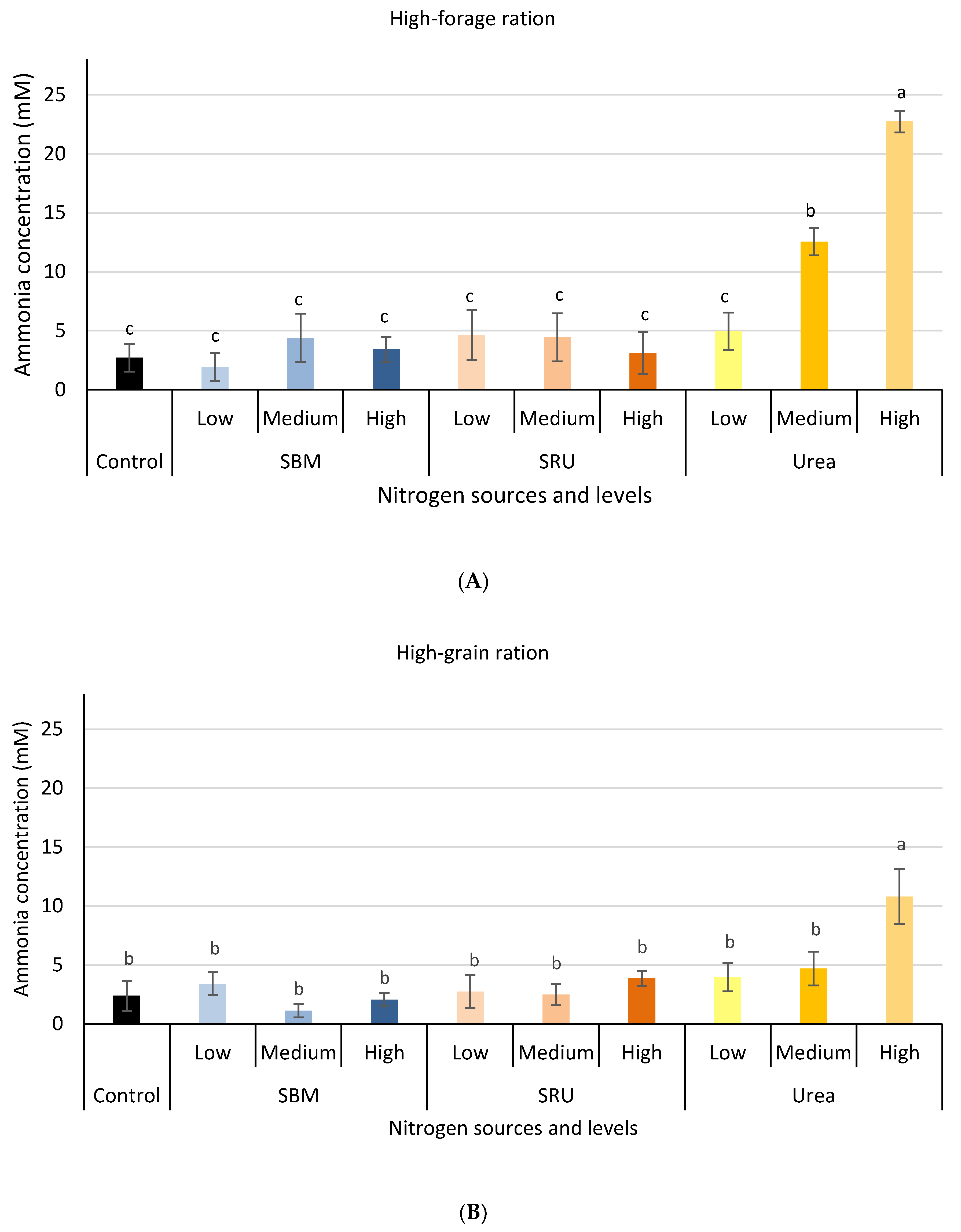
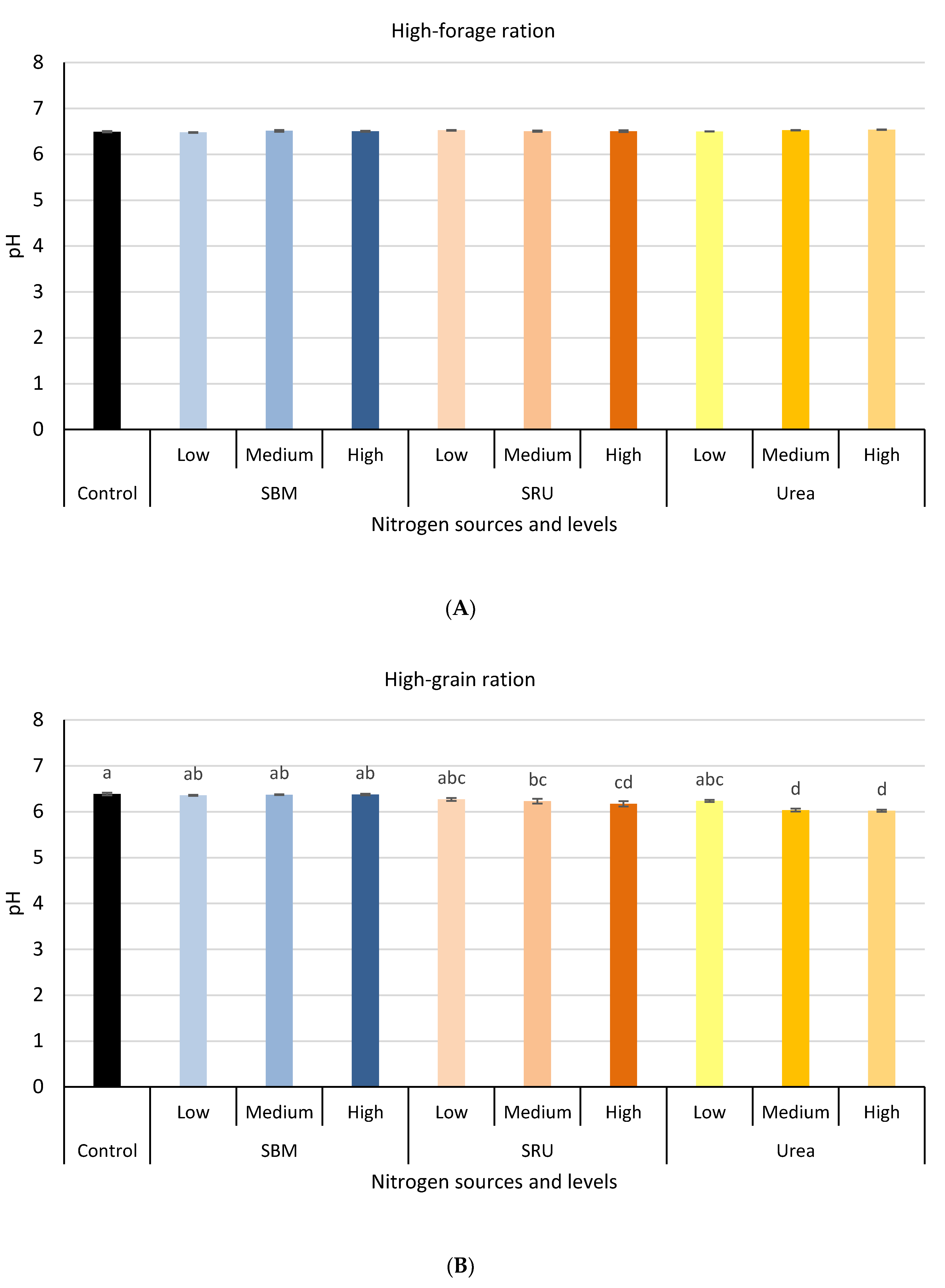
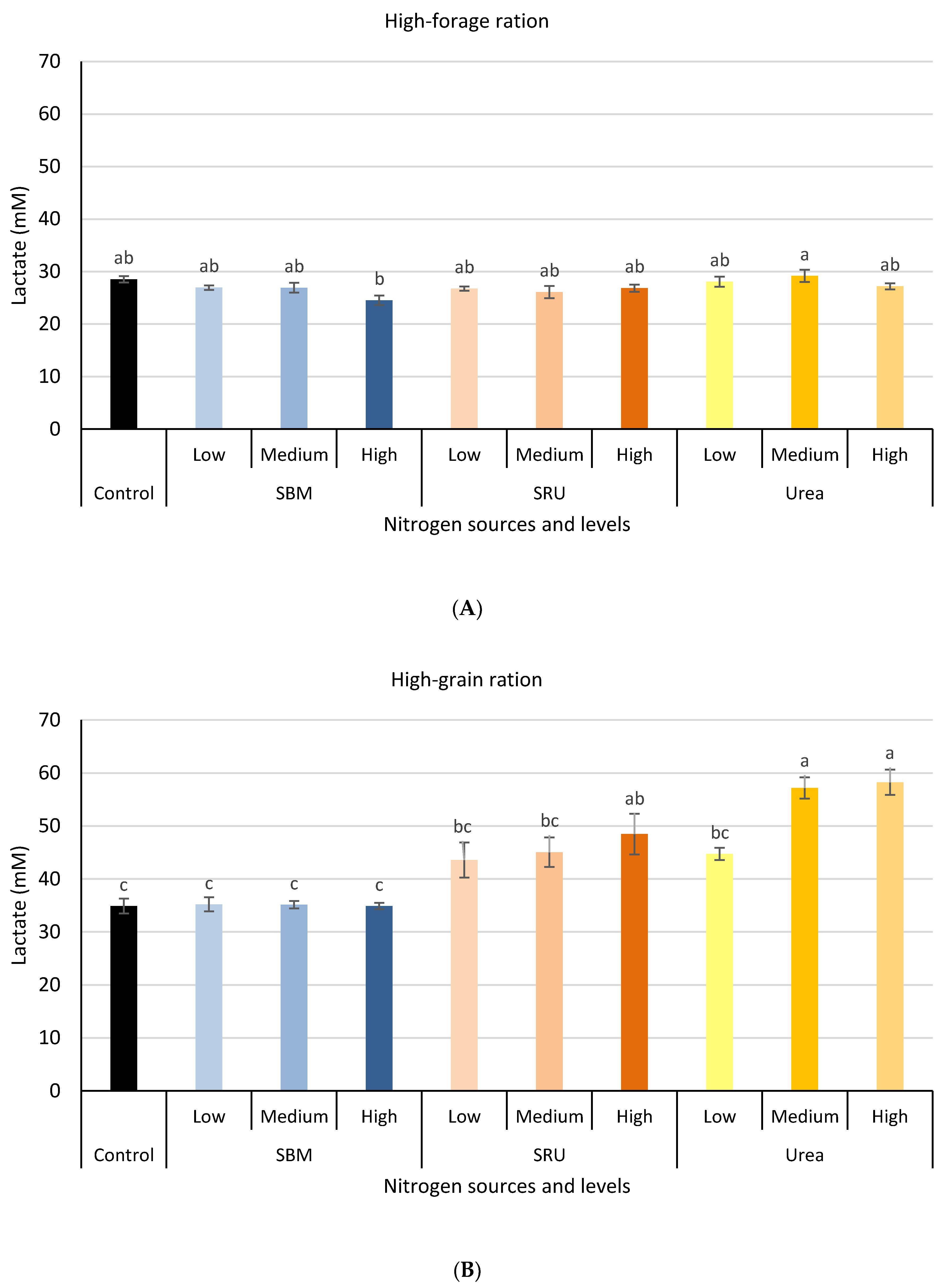
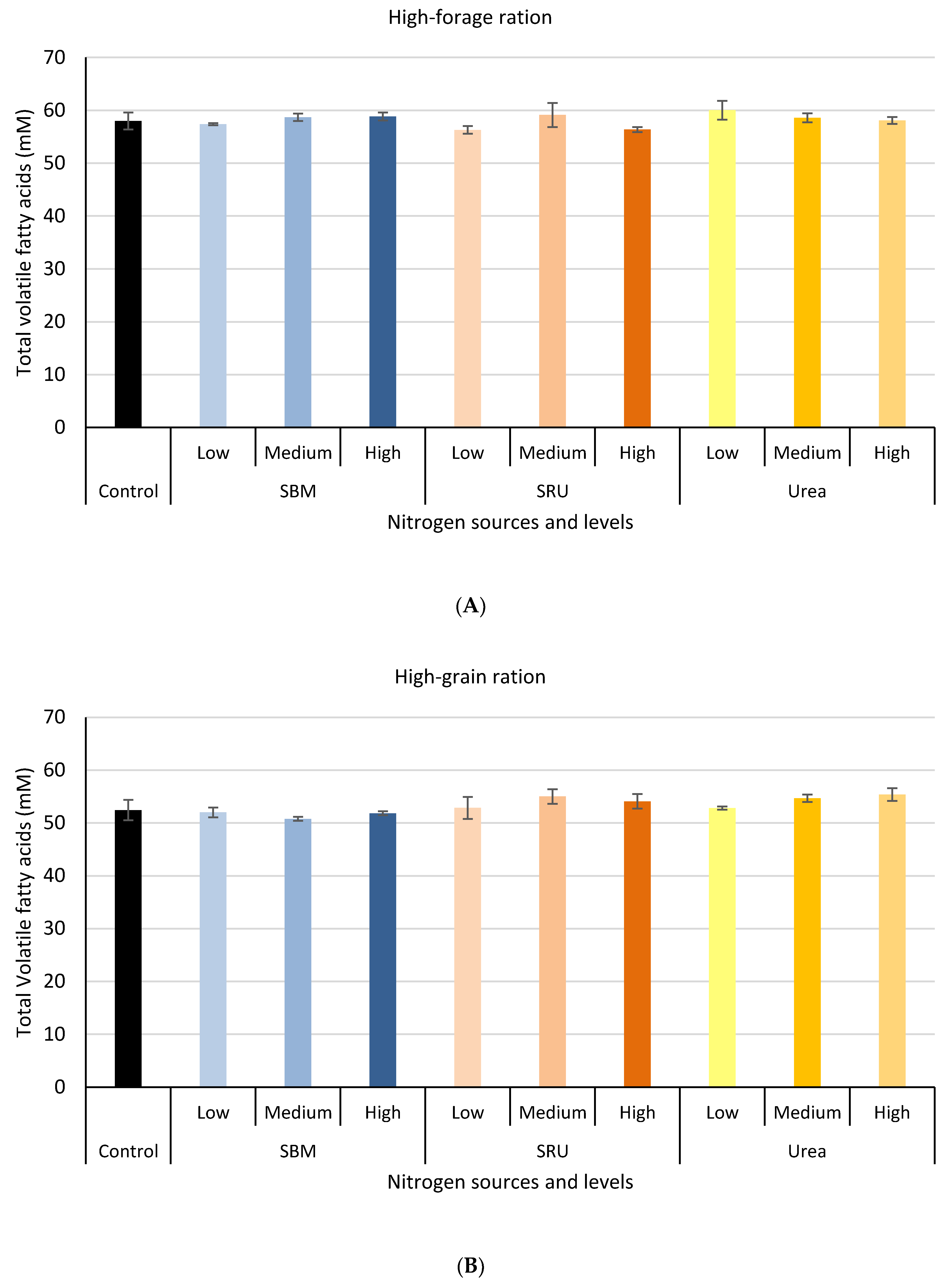
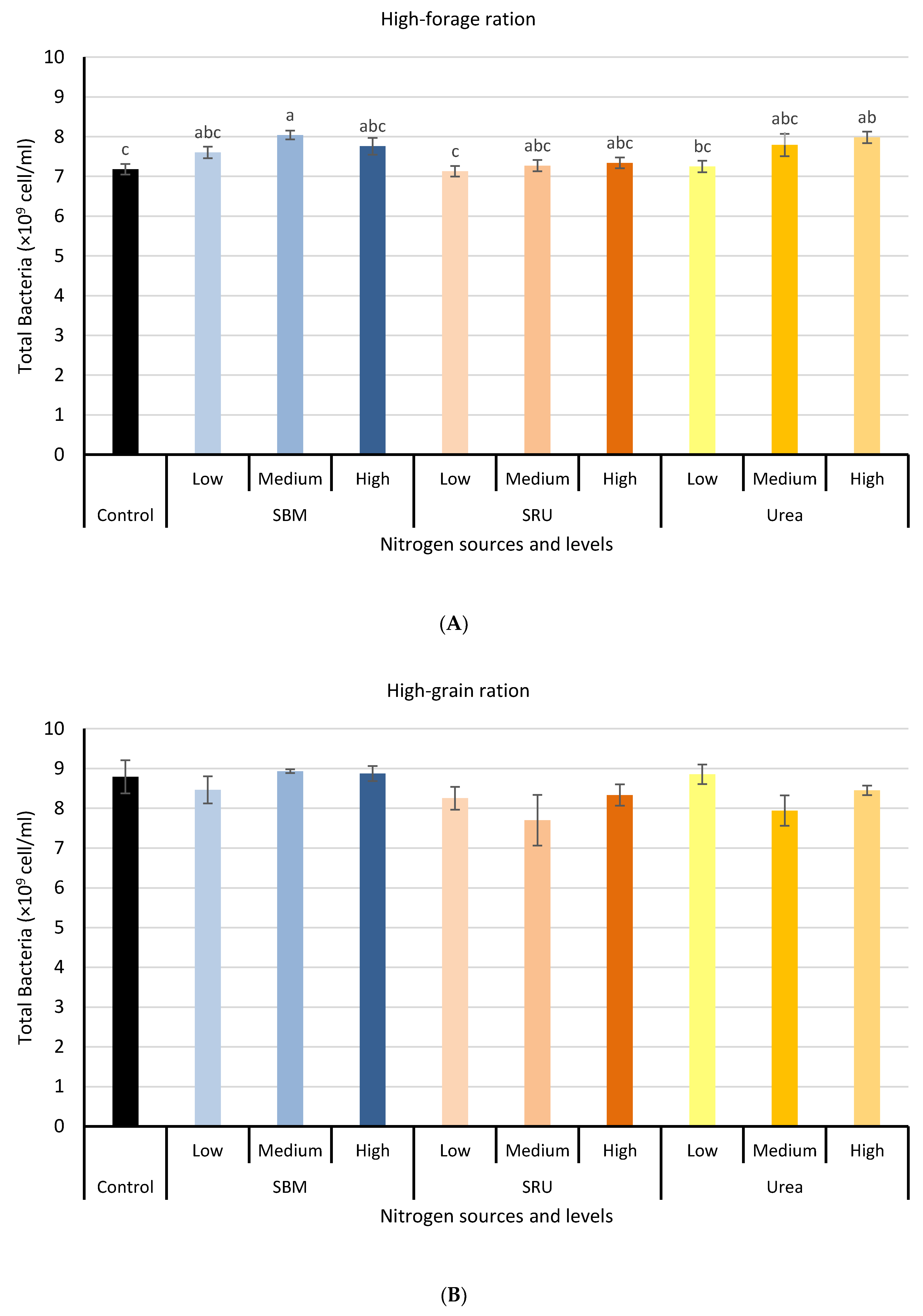
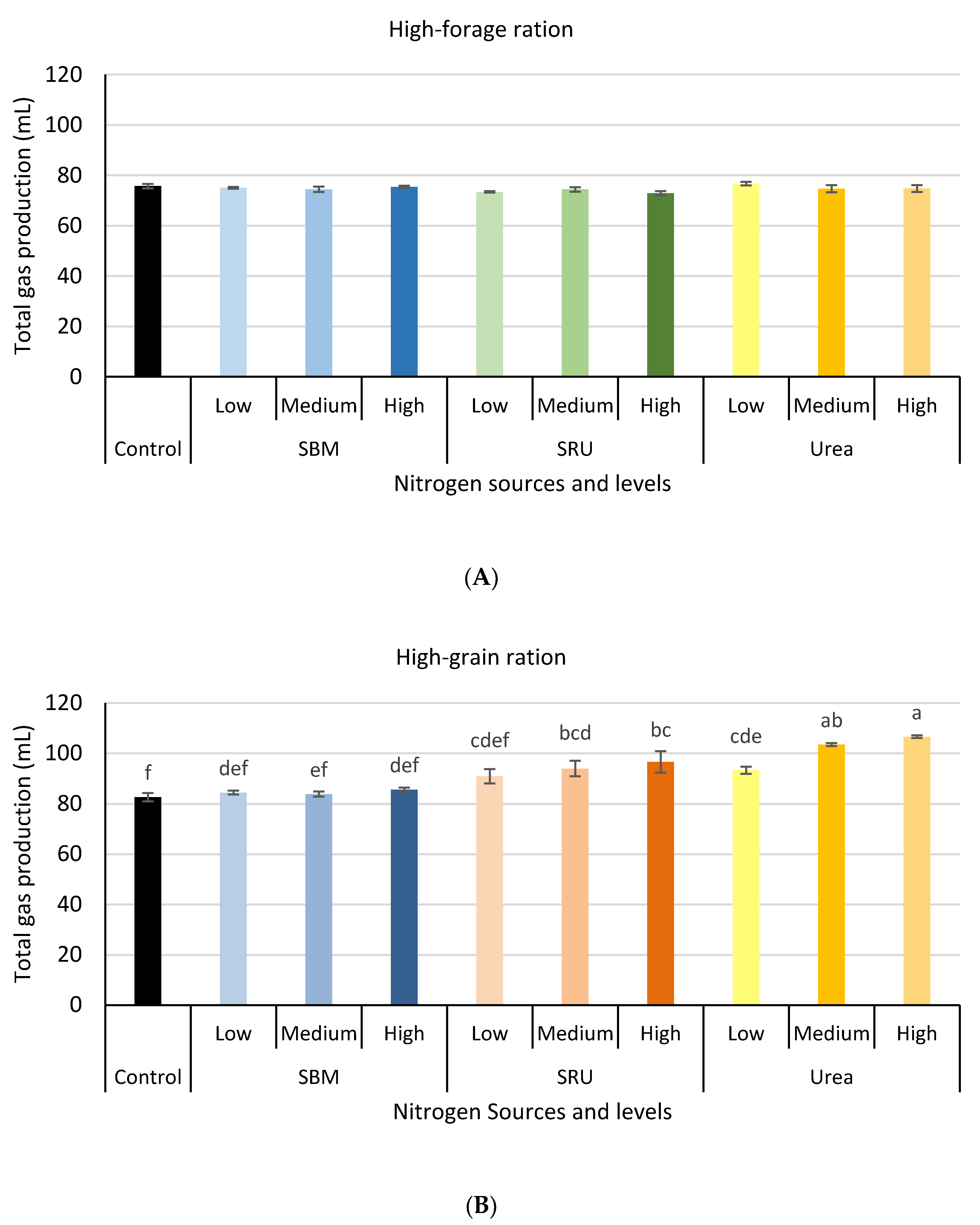
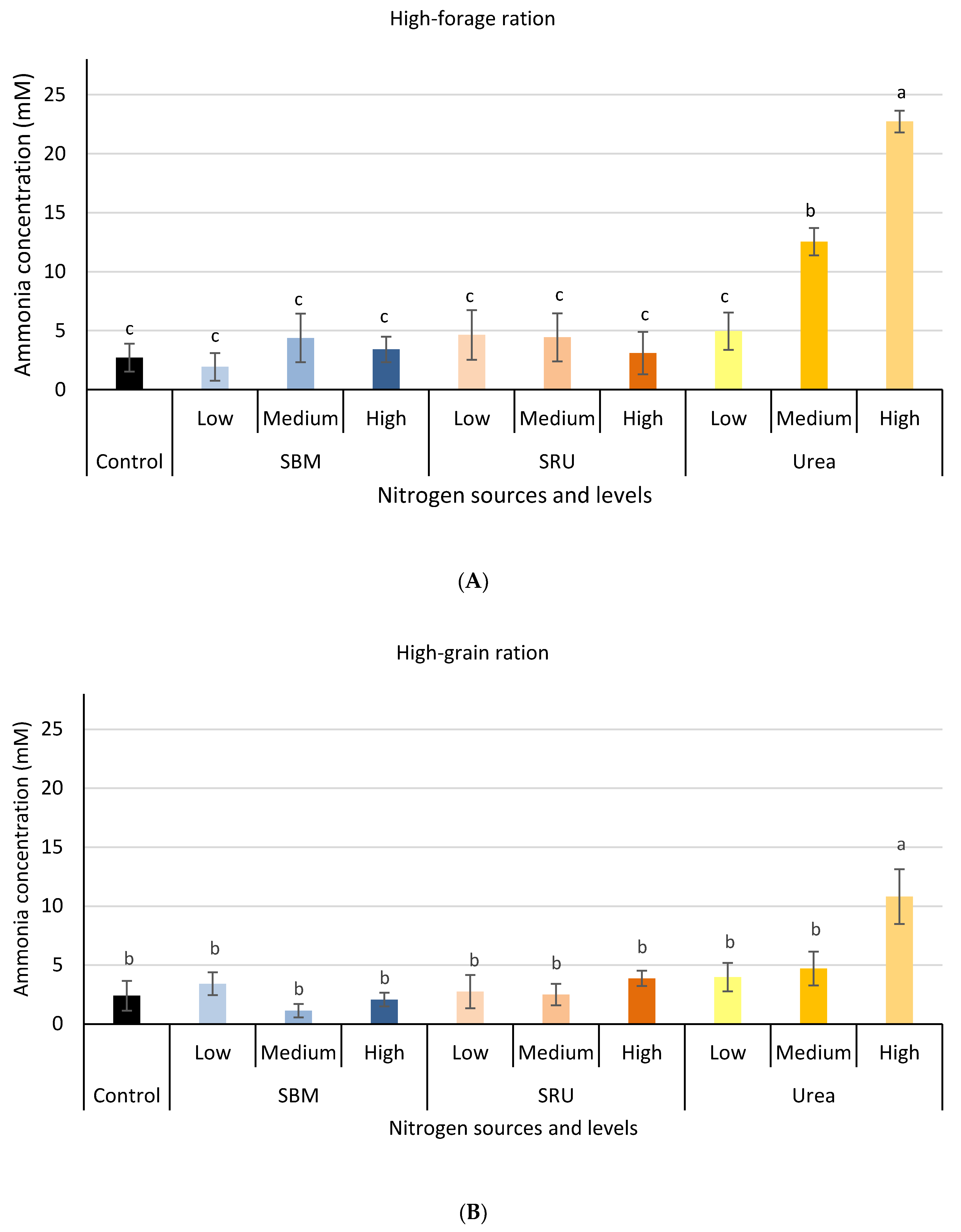
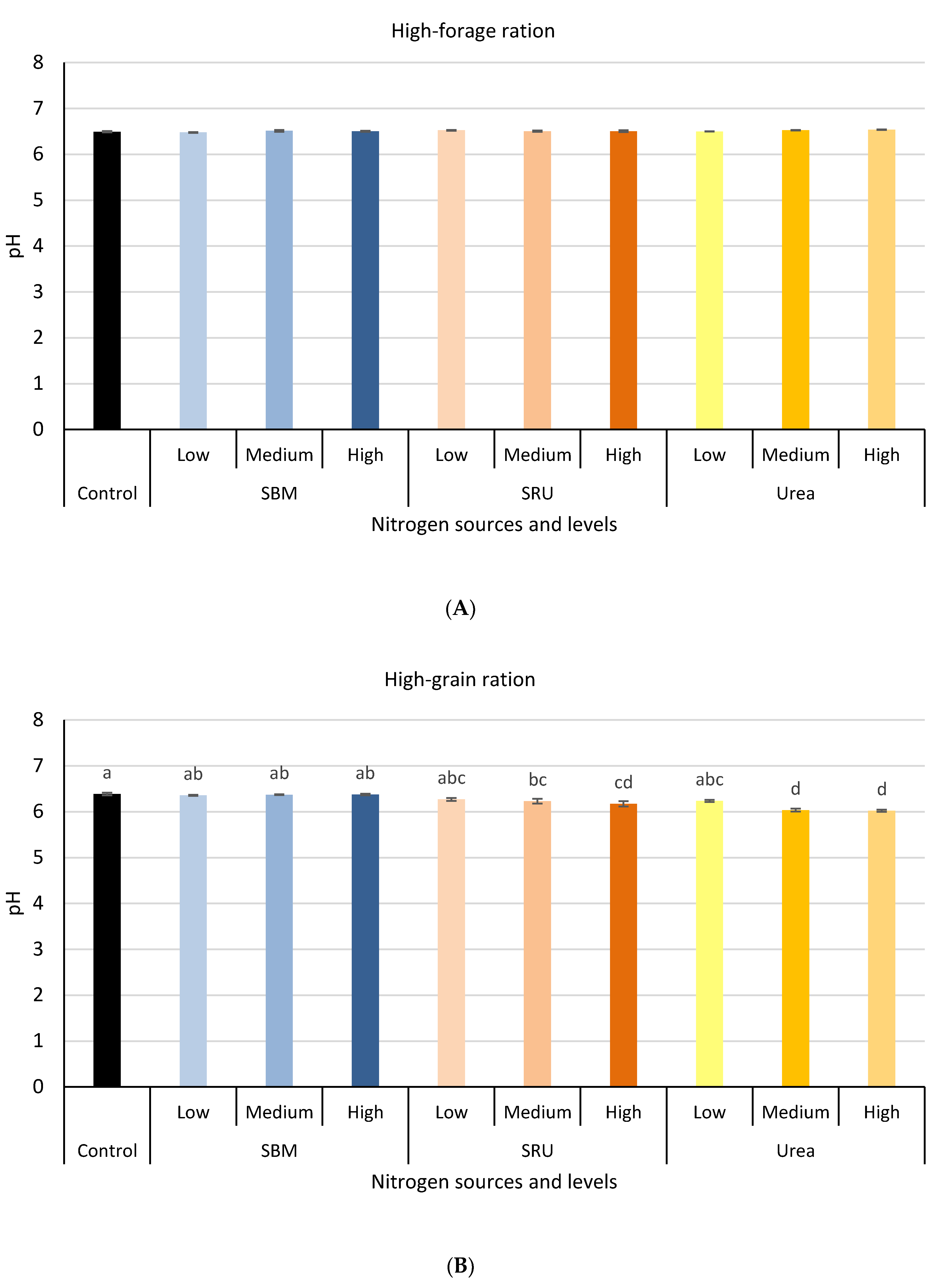
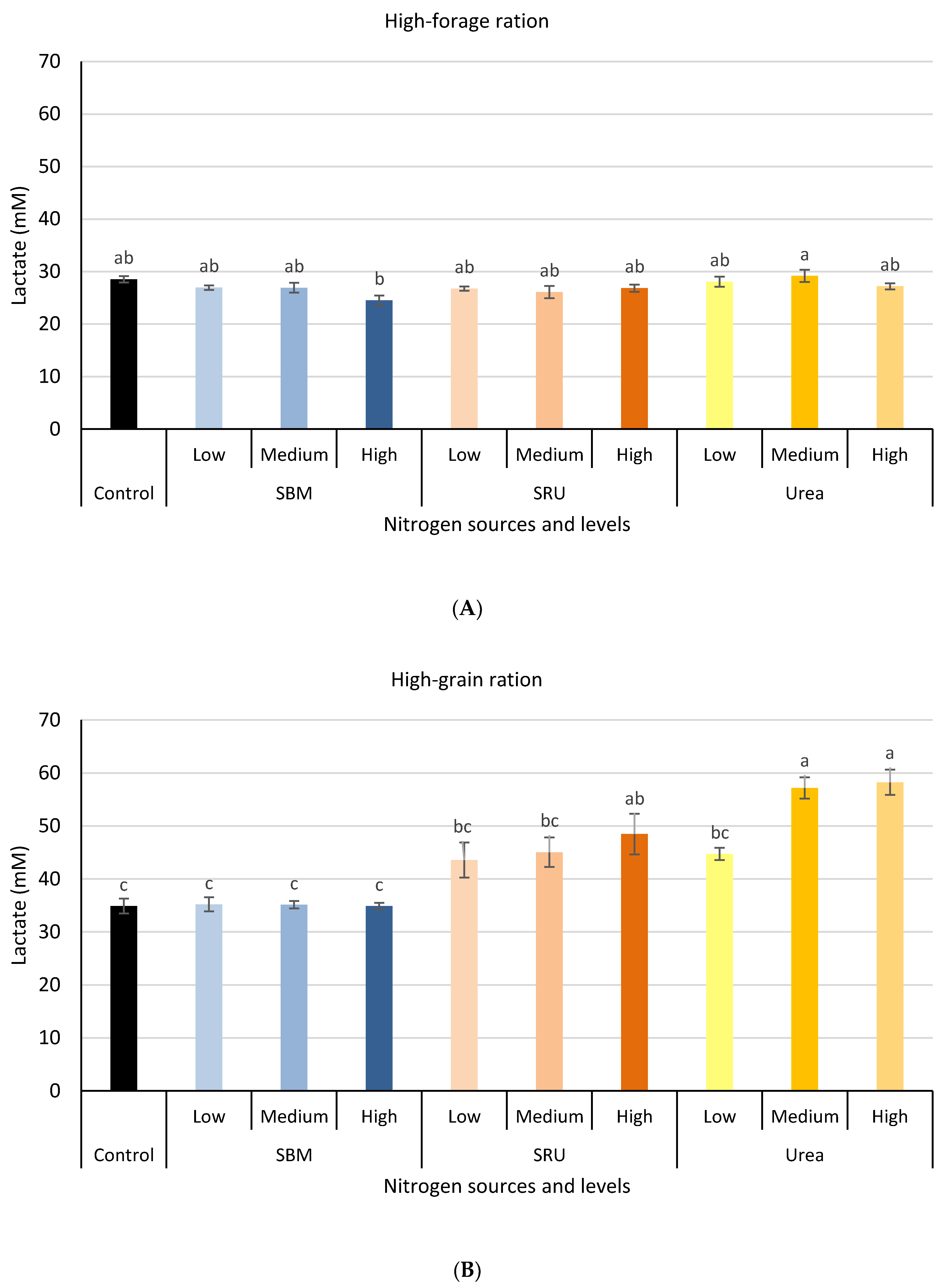
3.1. Experiment 1: In Vitro Rumen Fermentation
3.2. Experiment 2: In Situ Degradability
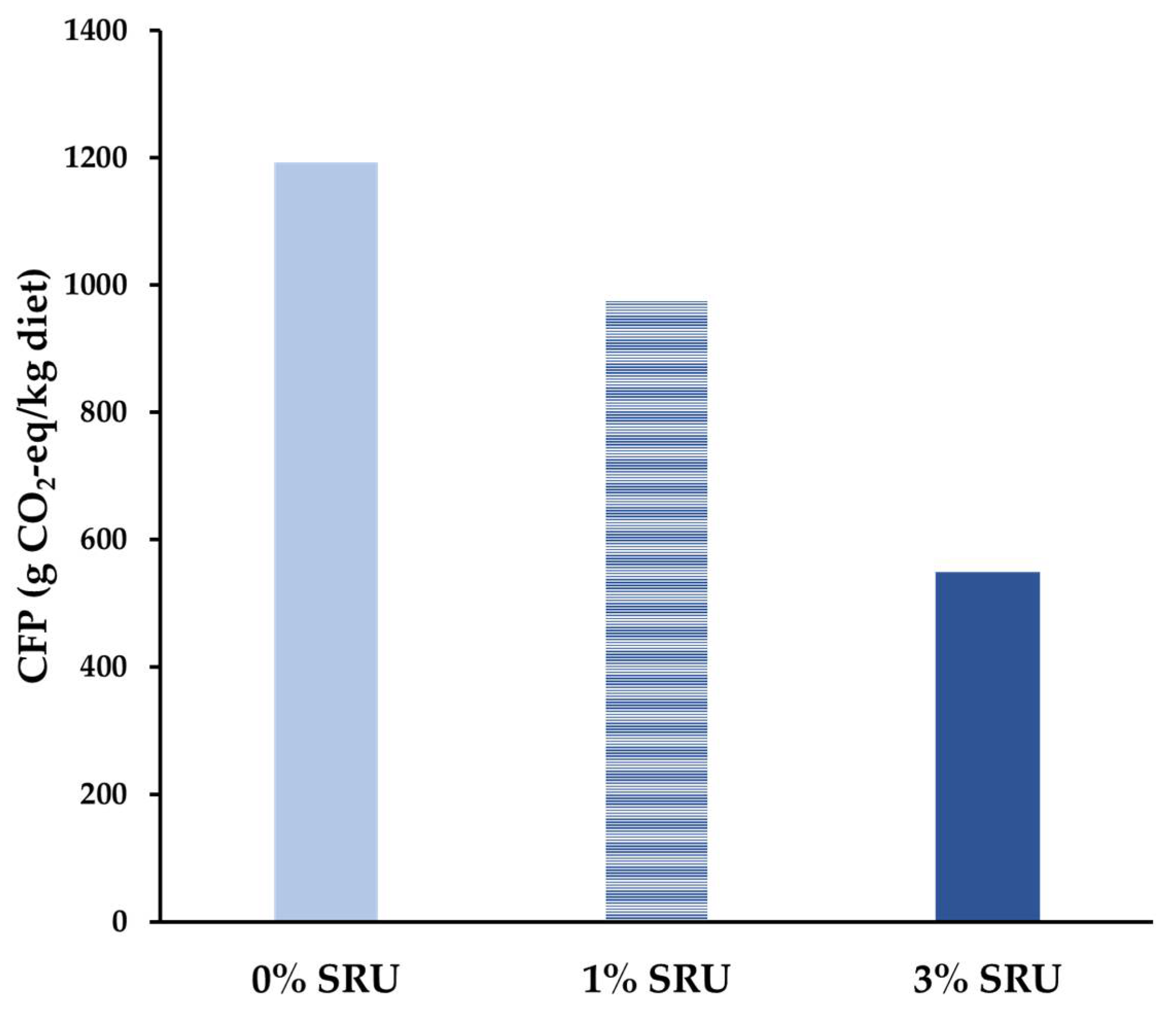
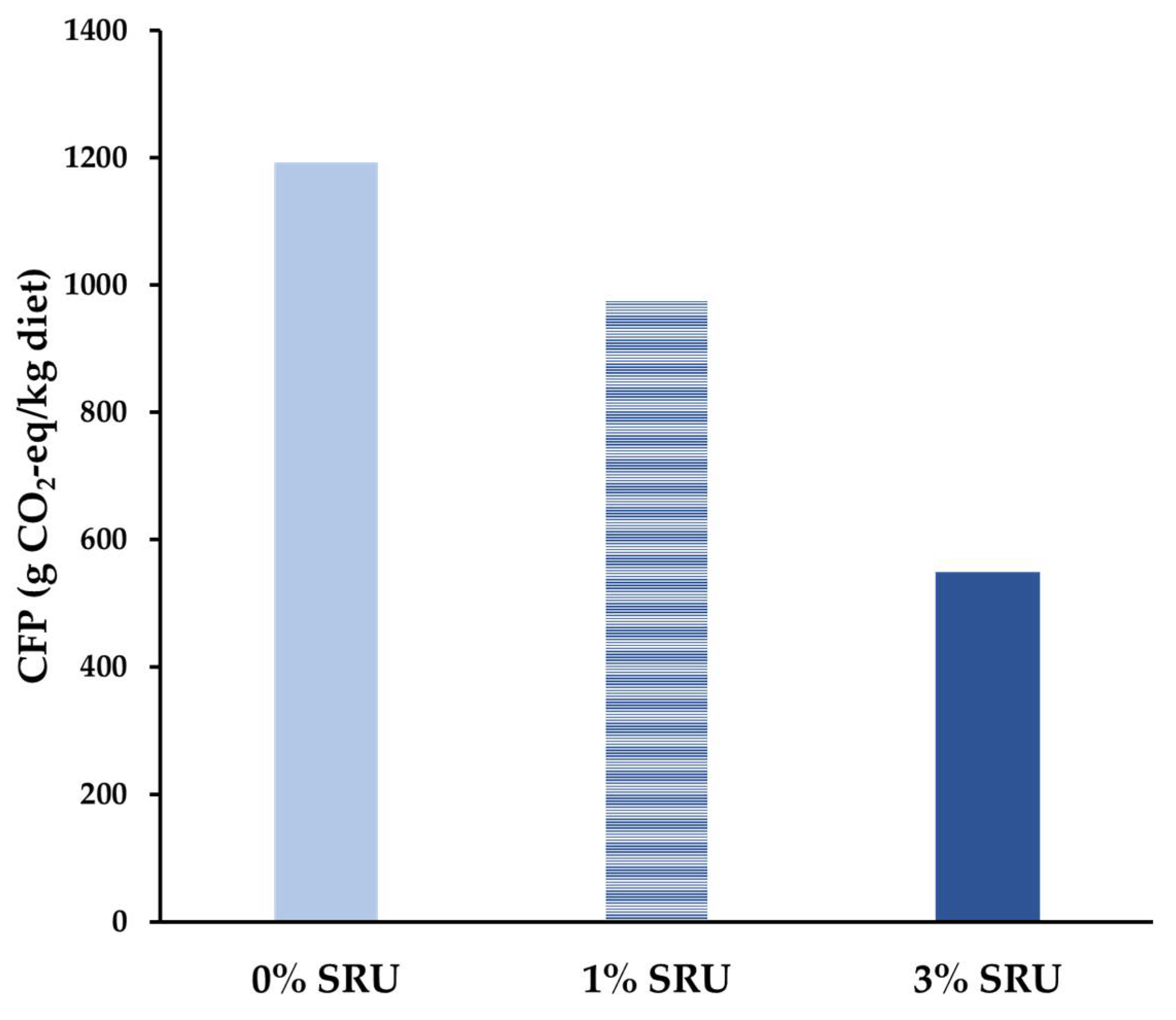
3.3. Experiment 3: Effect on Feed Intake and Blood Indices in Growing Cattle
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adesogan, A.T.; Havelaar, A.H.; McKune, S.L.; Eilittä, M.; Dahl, G.E. Animal source foods: Sustainability problem or malnutrition and sustainability solution? Perspective matters. Glob. Food Secur. 2020, 25, 100325. [Google Scholar] [CrossRef]
- Herrero, M.; Thornton, P.K. Livestock and global change: Emerging issues for sustainable food systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20878–20881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013.
- Wattiaux, M.; Uddin, M.; Letelier, P.; Jackson, R.; Larson, R. Invited Review: Emission and mitigation of greenhouse gases from dairy farms: The cow, the manure, and the field. Appl. Anim. Sci. 2019, 35, 238–254. [Google Scholar] [CrossRef]
- Wilkinson, J.; Garnsworthy, P. Impact of diet and fertility on greenhouse gas emissions and nitrogen efficiency of milk production. Livestock 2017, 22, 140–144. [Google Scholar] [CrossRef]
- Opio, C.; Gerber, P.; Mottet, A.; Falcucci, A.; Tempio, G.; MacLeod, M.; Vellinga, T.; Henderson, B.; Steinfeld, H. Greenhouse Gas Emissions from Ruminant Supply Chains–A Global Life Cycle Assessment; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013.
- Wilkinson, J.; Garnsworthy, P. Dietary options to reduce the environmental impact of milk production. J. Agric. Sci. 2017, 155, 334–347. [Google Scholar] [CrossRef]
- Salami, S.; Luciano, G.; O’Grady, M.; Biondi, L.; Newbold, C.; Kerry, J.; Priolo, A. Sustainability of feeding plant by-products: A review of the implications for ruminant meat production. Anim. Feed Sci. Technol. 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Reddy, P.R.K.; Kumar, D.S.; Rao, E.R.; Seshiah, C.V.; Sateesh, K.; Rao, K.A.; Reddy, Y.P.K.; Hyder, I. Environmental sustainability assessment of tropical dairy buffalo farming vis-a-vis sustainable feed replacement strategy. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Reddy, P.R.K.; Kumar, D.S.; Rao, E.R.; Seshiah, C.V.; Sateesh, K.; Reddy, Y.P.K.; Hyder, I. Assessment of eco-sustainability vis-à-vis zoo-technical attributes of soybean meal (SBM) replacement with varying levels of coated urea in Nellore sheep (Ovis aries). PLoS ONE 2019, 14, e0220252. [Google Scholar]
- Cherdthong, A.; Wanapat, M. Development of urea products as rumen slow-release feed for ruminant production: A review. Aust. J. Basic Appl. Sci. 2010, 4, 2232–2241. [Google Scholar]
- Calsamiglia, S.; Ferret, A.; Reynolds, C.; Kristensen, N.B.; Van Vuuren, A. Strategies for optimizing nitrogen use by ruminants. Animal 2010, 4, 1184–1196. [Google Scholar] [CrossRef] [PubMed]
- Cope, R.B. Nonprotein nitrogen (urea) and hyperammonemia. In Veterinary Toxicology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1093–1097. [Google Scholar]
- EFSA. EFSA Panel on Additives Products or Substances used in Animal Feed. Scientific Opinion on the safety and efficacy of Urea for ruminants. EFSA J. 2012, 10, 2624. [Google Scholar]
- Owens, F.; Lusby, K.; Mizwicki, K.; Forero, O. Slow ammonia release from urea: Rumen and metabolism studies. J. Anim. Sci. 1980, 50, 527–531. [Google Scholar] [CrossRef]
- Tedeschi, L.; Baker, M.; Ketchen, D.; Fox, D. Performance of growing and finishing cattle supplemented with a slow-rlease urea product and urea. Can. J. Anim. Sci. 2002, 82, 567–573. [Google Scholar] [CrossRef]
- Taylor-Edwards, C.; Hibbard, G.; Kitts, S.; McLeod, K.; Axe, D.; Vanzant, E.; Kristensen, N.; Harmon, D. Effects of slow-release urea on ruminal digesta characteristics and growth performance in beef steers. J. Anim. Sci. 2009, 87, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, L.; Blake, C.; Griffin, P.; Jones, G. The partial replacement of soyabean meal and rapeseed meal with feed grade urea or a slow-release urea and its effect on the performance, metabolism and digestibility in dairy cows. Animal 2012, 6, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gonzalez, R.; Tricarico, J.; Harrison, G.; Meyer, M.; McLeod, K.; Harmon, D.; Dawson, K. Optigen® is a sustained release source of non-protein nitrogen in the rumen. J. Anim. Sci. 2007, 85, 98. [Google Scholar]
- Ribeiro, S.; Vasconcelos, J.; Morais, M.; Ítavo, C.; Franco, G. Effects of ruminal infusion of a slow-release polymer-coated urea or conventional urea on apparent nutrient digestibility, in situ degradability, and rumen parameters in cattle fed low-quality hay. Anim. Feed Sci. Technol. 2011, 164, 53–61. [Google Scholar] [CrossRef]
- Vellinga, T.V.; Blonk, H.; Marinussen, M.; Van Zeist, W.; Starmans, D. Methodology Used in Feedprint: A Tool Quantifying Greenhouse Gas Emissions of Feed Production and Utilization; 1570-8616; Wageningen UR Livestock Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Apajalahti, J.; Vienola, K.; Raatikainen, K.; Holder, V.; Moran, C.A. Conversion of branched-chain amino acids to corresponding isoacids-an in vitro tool for estimating ruminal protein degradability. Front. Vet. Sci. 2019, 6, 311. [Google Scholar] [CrossRef] [PubMed]
- Meissner, H.; Henning, P.; Leeuw, K.-J.; Hagg, F.; Horn, C.; Kettunen, A.; Apajalahti, J. Efficacy and mode of action of selected non-ionophore antibiotics and direct-fed microbials in relation to Megasphaera elsdenii NCIMB 41125 during in vitro fermentation of an acidosis-causing substrate. Livest. Sci. 2014, 162, 115–125. [Google Scholar] [CrossRef]
- Salami, S.A.; Moran, C.A.; Warren, H.E.; Taylor-Pickard, J. A Meta-Analysis of the Effects of Slow-Release Urea Supplementation on the Performance of Beef Cattle. Animals 2020, 10, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apajalahti, J.H.; Kettunen, H.; Kettunen, A.; Holben, W.E.; Nurminen, P.H.; Rautonen, N.; Mutanen, M. Culture-independent microbial community analysis reveals that inulin in the diet primarily affects previously unknown bacteria in the mouse cecum. Appl. Environ. Microbiol. 2002, 68, 4986–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weatherburn, M. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Berthelot, M. Violet d’aniline. Rep. Chim. Appl. 1859, 1, 284. [Google Scholar]
- AOAC. Association of Official Analytical Chemists, 16th ed.; Official Methods of Analysis: Gaithersburg, MD, USA, 1996. [Google Scholar]
- Ørskov, E.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef] [Green Version]
- SAS. Stat User’s Guide; Cary, N.C., Ed.; Statistical Analysis Systems Inst. Inc.: Cary, NC, USA, 2001. [Google Scholar]
- NRC. Nutrient Requirements of Beef Cattle: Update 2000; National Academies Press: Washington, DC, USA, 2000. [Google Scholar]
- Van Soest, P.v.; Robertson, J.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Lumsden, J.; Mullen, K.; Rowe, R. Hematology and biochemistry reference values for female Holstein cattle. Can. J. Comp. Med. 1980, 44, 24. [Google Scholar] [PubMed]
- Martin, S.; Lumsden, J. The relationship of hematology and serum chemistry parameters to treatment for respiratory disease and weight gain in Ontario feedlot calves. Can. J. Vet. Res. 1987, 51, 499. [Google Scholar] [PubMed]
- Mohri, M.; Sharifi, K.; Eidi, S. Hematology and serum biochemistry of Holstein dairy calves: Age related changes and comparison with blood composition in adults. Res. Vet. Sci. 2007, 83, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; Moran, C.A.; Warren, H.E.; Taylor-Pickard, J. Meta-analysis and sustainability of feeding slow-release urea in dairy production. PLoS ONE 2021, 16, e0246922. [Google Scholar] [CrossRef] [PubMed]
- Kertz, A. Urea feeding to dairy cattle: A historical perspective and review. Prof. Anim. Sci. 2010, 26, 257–272. [Google Scholar] [CrossRef]
- Loor, J.; Elolimy, A.; McCann, J. Dietary impacts on rumen microbiota in beef and dairy production. Anim. Front. 2016, 6, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Getachew, G.; Robinson, P.; DePeters, E.; Taylor, S. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 2004, 111, 57–71. [Google Scholar] [CrossRef]
- Menke, K.H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Akay, V.; Tikofsky, J.; Holtz, C.; Dawson, K. Optigen® 1200: Controlled release of non-protein nitrogen in the rumen. In Proceedings of the 20th Alltech Symposium, Lexington, KY, USA, 23–24 May 2004; Alltech Inc.: Nicholasville, KY, USA, 2004; pp. 179–185. [Google Scholar]
- Bartley, E.; Davidovich, A.; Barr, G.; Griffel, G.; Dayton, A.; Deyoe, C.; Bechtle, R. Ammonia toxicity in cattle. I. Rumen and blood changes associated with toxicity and treatment methods. J. Anim. Sci. 1976, 43, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Chegeni, A.; Li, Y.; Deng, K.; Jiang, C.; Diao, Q. Effect of dietary polymer-coated urea and sodium bentonite on digestibility, rumen fermentation, and microbial protein yield in sheep fed high levels of corn stalk. Livest. Sci. 2013, 157, 141–150. [Google Scholar] [CrossRef]
- Kertz, A.; Davidson, L.; Cords, B.; Puch, H. Ruminal infusion of ammonium chloride in lactating cows to determine effect of pH on ammonia trapping. J. Dairy Sci. 1983, 66, 2597–2601. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Blanch, M.; Ferret, A.; Moya, D. Is subacute ruminal acidosis a pH related problem? Causes and tools for its control. Anim. Feed Sci. Technol. 2012, 172, 42–50. [Google Scholar] [CrossRef]
- Kenny, D.; Fitzsimons, C.; Waters, S.; McGee, M. Invited review: Improving feed efficiency of beef cattle–the current state of the art and future challenges. Animal 2018, 12, 1815–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, J.; Duval, S.M.; Tamassia, L.F.; Kindermann, M.; Stemmler, R.T.; de Gouvea, V.N.; Acedo, T.S.; Immig, I.; Williams, S.N.; Celi, P. Nutritional strategies in ruminants: A lifetime approach. Res. Vet. Sci. 2018, 116, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Bourg, B.; Tedeschi, L.; Wickersham, T.; Tricarico, J. Effects of a slow-release urea product on performance, carcass characteristics, and nitrogen balance of steers fed steam-flaked corn. J. Anim. Sci. 2012, 90, 3914–3923. [Google Scholar] [CrossRef] [PubMed]
- Inostroza, J.; Shaver, R.; Cabrera, V.; Tricárico, J. Effect of diets containing a controlled-release urea product on milk yield, milk composition, and milk component yields in commercial Wisconsin dairy herds and economic implications. Prof. Anim. Sci. 2010, 26, 175–180. [Google Scholar] [CrossRef]
- Radkowska, I.; Herbut, E. Hematological and biochemical blood parameters in dairy cows depending on the management system. Anim. Sci. Papers Rep. 2014, 32, 317–325. [Google Scholar]
- Roland, L.; Drillich, M.; Iwersen, M. Hematology as a diagnostic tool in bovine medicine. J. Vet. Diagn. Investig. 2014, 26, 592–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobley, G.; Connell, A.; Lomax, M.A.; Brown, D.S.; Milne, E.; Calder, A.G.; Farningham, D. Hepatic detoxification of ammonia in the ovine liver: Possible consequences for amino acid catabolism. Br. J. Nutr. 1995, 73, 667–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonds, H.; Mather, D.L.; Collis, K. The maximum capacity of the liver of the adult dairy cow to metabolize ammonia. Br. J. Nutr. 1981, 46, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Word, J.; Martin, L.; Williams, D.; Williams, E.; Panciera, R.; Nelson, T.; Tillman, A. Urea toxicity studies in the bovine. J. Anim. Sci. 1969, 29, 786–791. [Google Scholar] [CrossRef]
- Holder, V.B.; El-Kadi, S.W.; Tricarico, J.M.; Vanzant, E.S.; McLeod, K.R.; Harmon, D.L. The effects of crude protein concentration and slow release urea on nitrogen metabolism in Holstein steers. Arch. Anim. Nutr. 2013, 67, 93–103. [Google Scholar] [CrossRef]
- Ribeiro, M.N.; Ribeiro, N.L.; Bozzi, R.; Costa, R.G. Physiological and biochemical blood variables of goats subjected to heat stress–a review. J. Appl. Anim. Res. 2018, 46, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Huntington, G.; Archibeque, S. Practical aspects of urea and ammonia metabolism in ruminants. Proc. Am. Soc. Anim. Sci. 2000, 77, 1–11. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Number | Ration 1 | Nitrogen Supplement | Dose, mg (Nitrogen Levels) 2 | Replicate Fermentations |
|---|---|---|---|---|
| 1 | High-forage | None | - | 5 |
| 2 | SBM | 20 (low) | 5 | |
| 3 | 60 (medium) | 5 | ||
| 4 | 100 (high) | 5 | ||
| 5 | SRU | 2.6 (low) | 5 | |
| 6 | 7.8 (medium) | 5 | ||
| 7 | 13.1 (high) | 5 | ||
| 8 | Urea | 2.3 (low) | 5 | |
| 9 | 6.9 (medium) | 5 | ||
| 10 | 11.5 (high) | 5 | ||
| 11 | High-grain | None | - | 5 |
| 12 | SBM | 20 (low) | 5 | |
| 13 | 60 (medium) | 5 | ||
| 14 | 100 (high) | 5 | ||
| 15 | SRU | 2.6 (low) | 5 | |
| 16 | 7.8 (medium) | 5 | ||
| 17 | 13.1 (high) | 5 | ||
| 18 | Urea | 2.3 (low) | 5 | |
| 19 | 6.9 (medium) | 5 | ||
| 20 | 11.5 (high) | 5 | ||
| Total | 100 |
| Diet Component | Nitrogen Content (g/kg DM) | Crude Protein (g/kg DM) | DM (%) | % of Diet DM | |
|---|---|---|---|---|---|
| Control | SRU | ||||
| Soybean meal | 82.3 | 514 | 88.9 | 15 | 10 |
| SRU | 410 | 2563 | 100 | 0 | 0.75 |
| Compound feed 1 | 19.9 | 124 | 90 | 44 | 45.25 |
| Grass haylage | 26.2 | 164 | 72.1 | 41 | 44 |
| Ingredient (%) | 0% SRU | 1% SRU | 3% SRU |
|---|---|---|---|
| Corn grain meal | 38.19 | 40.96 | 48.00 |
| Wheat middlings | 3.02 | 3.02 | 2.00 |
| Barley grain meal | 17.02 | 19.02 | 22.61 |
| Wheat | 2.51 | 2.51 | 2.00 |
| Beet pulp | 7.99 | 7.99 | 9.99 |
| Dried alfalfa | 8.33 | 8.33 | 6.52 |
| Palm oil | 2.46 | 2.85 | 3.27 |
| Soybean meal | 18.38 | 12.16 | 0.00 |
| Calcium carbonate | 0.80 | 0.86 | 0.80 |
| Sodium bicarbonate | 0.60 | 0.60 | 0.60 |
| Salt | 0.40 | 0.40 | 0.40 |
| Bicalcium phosphate | 0.00 | 0.00 | 0.51 |
| Vitamin/mineral premix 809 1 | 0.30 | 0.30 | 0.30 |
| Slow Release Urea | 0.00 | 1.00 | 3.00 |
| Analyzed nutrient content (% DM) | |||
| DM | 87.46 | 87.81 | 87.51 |
| Crude protein | 18.22 | 18.03 | 18.28 |
| Ether extract | 4.28 | 5.39 | 6.15 |
| Neutral detergent fibre | 26.17 | 25.56 | 26.14 |
| Ash | 5.99 | 5.62 | 4.86 |
| Urea | - | <0.1 | 0.88 |
| Calculated NSC | 53.2 | 53.1 | 53.3 |
| Parameter | Control | SRU | SEM | p-Value |
|---|---|---|---|---|
| DM degradation | ||||
| b | 68.30 | 62.31 | 0.029 | 0.004 |
| c | 29.05 | 31.27 | 0.686 | 0.263 |
| Kd | 0.15 | 0.16 | 0.006 | 0.640 |
| ERD | 51.86 | 51.71 | 0.219 | 0.716 |
| Nitrogen degradation | ||||
| b | 88.40 | 85.91 | 0.509 | 0.179 |
| c | 11.60 | 14.09 | 0.509 | 0.179 |
| Kd | 0.10 | 0.10 | 0.002 | 0.252 |
| ERD | 54.40 | 53.98 | 0.024 | 0.052 |
| Parameter | 0% SRU | 1% SRU | 3% SRU | SEM | p-Value | Reference Values 1 |
|---|---|---|---|---|---|---|
| Red cell count (×106 cells/µL) | 9.6 | 9.5 | 9.7 | 0.15 | 0.649 | 6.5–11.9 |
| Hemoglobin (g/dL) | 13.0 a | 12.3 b | 12.3 b | 0.19 | 0.028 | 8–14.1 |
| Packed cell volume (%) | 33.1 a | 31.1 b | 31.2 b | 0.44 | 0.007 | 23–42 |
| Mean corpuscular volume (fL) | 34.2 a | 33.1 ab | 32.6 b | 0.44 | 0.090 | 26.6–44.3 |
| Mean corpuscular hemoglobin (pg) | 13.5 a | 13.1 ab | 12.8 b | 0.18 | 0.061 | 9.1–15.6 |
| White cell count (×103 cells/µL) | 11.6 | 10.9 | 11.9 | 0.92 | 0.609 | 5.6–13.7 |
| Neutrophils (×103 cells/µL) | 4.6 | 3.7 | 4.5 | 0.67 | 0.428 | 0.6–6.1 |
| Lymphocytes (×103 cells/µL) | 6.1 | 6.4 | 6.8 | 0.46 | 0.527 | 2.2–8.7 |
| Monocytes (×103 cells/µL) | 0.5 | 0.5 | 0.5 | 0.06 | 0.809 | 0.1–0.2 |
| Eosinophils (×103 cells/µL) | 0.2 | 0.2 | 0.2 | 0.08 | 0.902 | 0–0.3 |
| Basophils (×103 cells/µL) | 0.3 | 0.3 | 0.4 | 0.03 | 0.519 | 0–0.1 |
| Platelets (×103 cells/µL) | 580 | 591 | 524 | 48 | 0.752 | 220–950 |
| Parameter | 0% SRU | 1% SRU | 3% SRU | SEM | p-Value | Reference Values 1 |
|---|---|---|---|---|---|---|
| Glucose (mg/dL) | 101.2 | 103.0 | 102.9 | 2.91 | 0.879 | 45–135 |
| Aspartate aminotransferase (IU/L) | 66.1 a | 56.9 b | 62.5 ab | 3.03 | 0.067 | <60 |
| Alanine aminotransferase (IU/L) | 17.2 | 14.9 | 16.6 | 0.75 | 0.114 | 3–18 |
| Gamma-glutamyl transferase (IU/L) | 13.4 | 12.1 | 11.7 | 1.13 | 0.534 | <16 |
| Urea (mg/dL) | 27.5 | 28.2 | 24.4 | 1.44 | 0.180 | 21–54 |
| Uric acid (mg/dL) | 0.4 | 0.4 | 0.4 | - | 0.679 | - |
| Albumin (g/dL) | 3.7 a | 3.4 c | 3.6 b | 0.03 | 0.001 | 2.5–3.8 |
| Ammonia (mg/dL) | 0.7 | 0.7 | 0.9 | 0.09 | 0.105 | <5 |
| Total protein (g/dL) | 6.8 a | 6.6 b | 6.5 b | 0.09 | 0.031 | 6.8–7.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salami, S.A.; Devant, M.; Apajalahti, J.; Holder, V.; Salomaa, S.; Keegan, J.D.; Moran, C.A. Slow-Release Urea as a Sustainable Alternative to Soybean Meal in Ruminant Nutrition. Sustainability 2021, 13, 2464. https://doi.org/10.3390/su13052464
Salami SA, Devant M, Apajalahti J, Holder V, Salomaa S, Keegan JD, Moran CA. Slow-Release Urea as a Sustainable Alternative to Soybean Meal in Ruminant Nutrition. Sustainability. 2021; 13(5):2464. https://doi.org/10.3390/su13052464
Chicago/Turabian StyleSalami, Saheed A., Maria Devant, Juha Apajalahti, Vaughn Holder, Sini Salomaa, Jason D. Keegan, and Colm A. Moran. 2021. "Slow-Release Urea as a Sustainable Alternative to Soybean Meal in Ruminant Nutrition" Sustainability 13, no. 5: 2464. https://doi.org/10.3390/su13052464
APA StyleSalami, S. A., Devant, M., Apajalahti, J., Holder, V., Salomaa, S., Keegan, J. D., & Moran, C. A. (2021). Slow-Release Urea as a Sustainable Alternative to Soybean Meal in Ruminant Nutrition. Sustainability, 13(5), 2464. https://doi.org/10.3390/su13052464