
Utility of Human Footprint Pressure Mapping for Large Carnivore Conservation: The Kafue-Zambezi Interface


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generating Human Footprint Pressure Maps
- Settlement data was derived from Bonafilia et. al. [43] at 30 m resolution. All pixels overlapping settlement areas were given a pressure score of 10 representing the highest level of direct pressure (implying settled area were unsuitable for wildlife), with all other pixels given a score of 0.
- Human Population Density data was unavailable at sufficiently fine scale for the landscape to include as a stand-alone data layer. Given the largely homogenous nature of settlement throughout the area (an absence of large multi-story buildings and dense conurbations versus ubiquitous single-story concrete block and tin buildings with scattered adobe and grass huts throughout rural area (Lines, pers obs)), we calculated average population density for the study area from the district scale data using 2019 population projected data [40]. Assuming that the total population of a district exists within the area of settlements, the total population was divided with the total area occupied by settlements, to provide their average population density. We then applied the log formula employed by Venter et al. [17] for scarcely populated areas (Table 1). The calculated score was applied to all pixels that constitute part of a settlement.
- Roads, acquired by the Peace Parks Foundation (PPF), constitute both a direct and an indirect human pressure. They reduce the extent of suitable habitats and the degree of habitat fragmentation while at the same time they are associated with increased traffic-induced mortality [45]. The indirect impacts are associated with the increased accessibility to wild areas ensured by a dense road network.
- The same approach adopted by Venter et al. [17] for scoring the human pressure associated with roads was adopted in the current study. A pressure score of 8 was assigned to all pixels in a distance of 0.5 km either side of roads, indicating high direct human pressure. A pressure score of 4, exponentially decaying out to 15 km, was assigned to pixels in a distance longer from 0.5 km either side of the roads up to 15 km away, indicating lower indirect pressures as distance from the roads increases. The threshold of 15 km was set as it represents the approximate distance a person might reasonably access on foot within a day. A vector roads layer, provided by Peace Parks Foundation (unpublished data), including major tar and secondary dirt roads linking settlements, formed the baseline for the generation of the roads raster layer at a spatial resolution of 10 m. Tertiary dirt tracks were omitted from the analysis due to their dynamic nature and inconsistent mapping. The range of pressure scores varied between 0.25 at a distance of 15 km from roads to 8 for pixels next to roads.
- Railways, acquired by the PPF, represent direct drivers of habitat conversion and conduits of human access into wildlife areas similar to roads. Since passengers cannot commonly disembark at will, indirect effects away from the railway line are considered minimal. Following Venter et al. [17], we gave railways a direct pressure score of 8 for a distance of 0.5 km either side of the railway using the same method as for roads.
- Navigable Waterways, acquired by the PPF, like roads, provide direct access to wildlife habitats along the waterway, and indirect access in periphery areas. The Zambezi River is the only permanent navigable waterway in our study area, and following Venter et al. [17], we assigned a pressure score of 4 to pixels adjacent to the river, exponentially decaying out to 15 km.
- Arable land throughout the Kafue–Zambezi interface is characterized by majority maize and pulses cultivated using the traditional Chitemene low input, rain fed, slash and burn farming method [46]. Arable land cover classifications are considered by Venter et al. [17] to provide intermediate disturbance to wildlife though direct reduction of wildlife habitat.
- The arable land was extracted by a land cover map produced by Lines et al. [37] using a mosaic of 24, geometrically and atmospherically corrected, Sentinel 2 images in an Object Oriented Image Analysis environment. The land cover map had a spatial resolution of 10 m and an overall classification accuracy of 91.6% [37]. A pressure score of 7 was assigned to all pixels covered by arable land and 0 to all other pixels [17].
- Night-time light infrastructure, while sparse and of low intensity throughout much of our study area, is considered a direct human pressure limiting wildlife through a range of negative impacts [47].
- The “vcm-orm-ntl” (VIIRS Cloud Mask—Outlier Removed—Night-time Lights) annual average layer was used [44], for generating the respective pressure layer. Pixels with a value of 0 (no light) were assigned the value of 0 in the generated layer. For all other pixels, following Venter et al. [17], we applied the same log formula used for pop density (Table 1) resulting in pixels with scores ranging from 0 to 8.971.
- Aggregating the layers: All generated layers were added to generate an aggregate layer indicating for each pixel the total Human Footprint Pressure. Before aggregation all layers generated at a spatial resolution coarser than 10 m were resampled to a spatial resolution of 10 m. The resolution of 10 m was adopted for this analysis, because it corresponds to the resolution of the land cover data and it is the finest among all data. While the resampling to a finer resolution does not affect the quality of data provided at coarser resolutions, a resampling of fine resolution data to a coarser resolution would probably result to information loss. The aggregated Human Footprint Pressure layer was used as the single explanatory variable in a habitat suitability modelling analysis.
2.2. MaxEnt Habitat Suitability Modelling
3. Results
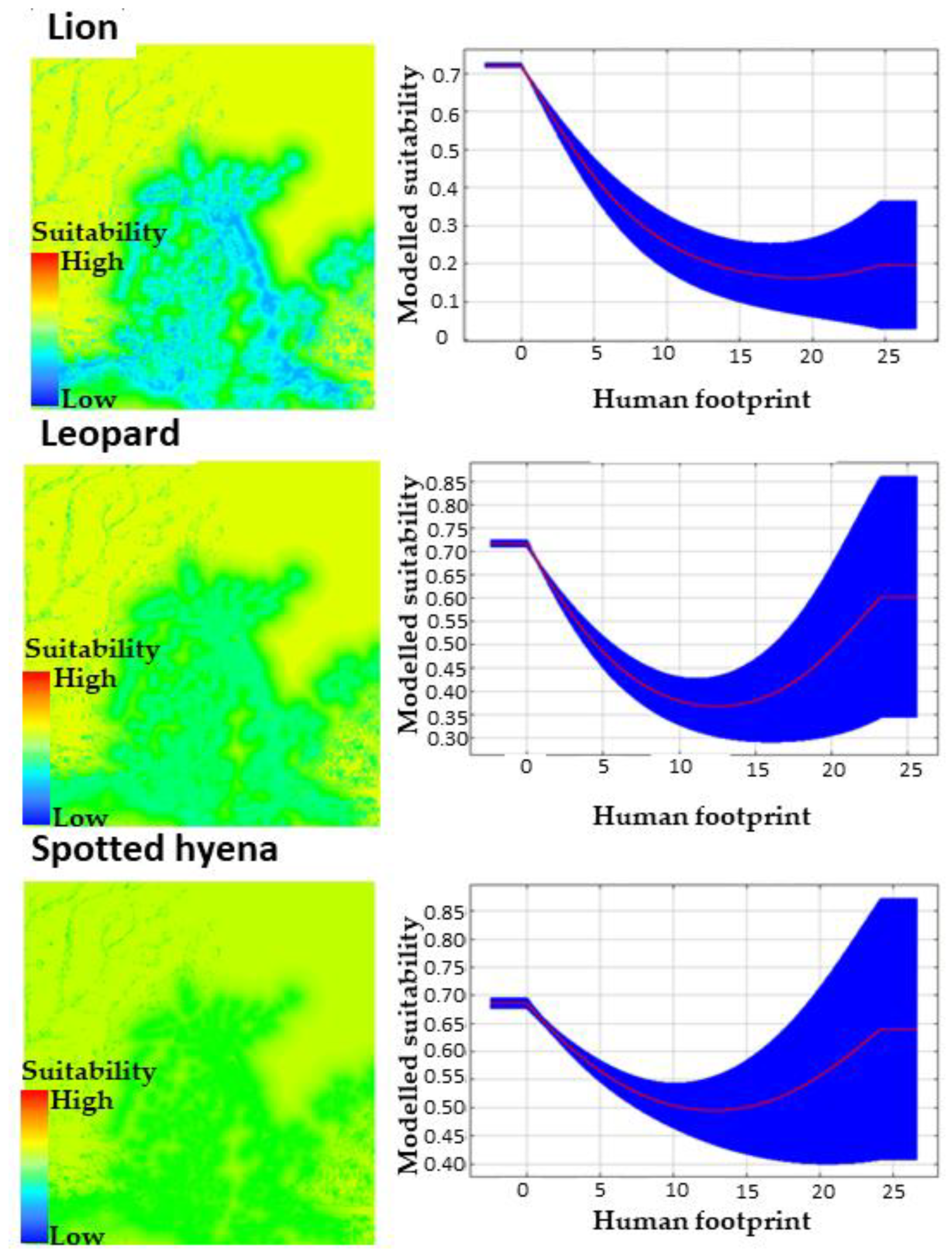
3.1. MaxEnt Habitat Suitability Modelling Outputs
3.2. Human Footprint Pressure Thresholds and Species Persistence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.; Mooney, H.; Lubchenco, J.; Melillo, J. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Wackernagel, M.; Schulz, N.B.; Deumling, D.; Linares, C.A.; Jenkins, M.; Kapos, V.; Monfreda, C.; Loh, J.; Myers, N.; Norgaard, R.; et al. Tracking the ecological overshoot of the human economy. Proc. Natl. Acad. Sci. USA 2002, 99, 9266–9271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riitters, K.; Wickham, J.; O’Neill, R.; Jones, B.; Smith, E. Global-scale patterns of forest fragmentation. Conserv. Ecol. 2000, 4, 3. Available online: http://www.consecol.org/vol4/iss2/art3/ (accessed on 20 October 2021). [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; Carpenter, S.R.; Chapin, F.S.; et al. Global consequences of land use. Science 2005, 333, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.A.; Fagan, W.F.; Fryxell, J.M.; Van Bram, M.; Susan, C.A.; Abdullahi, H.A.; Andrew, M.A.; Attias, N.; Tal, A.; Brooks, B.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E. Biodiversity: The ravages of guns, nets and bulldozers. Nat. News 2016, 546, 143. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R. Mammal population losses and the extinction crisis. Science 2002, 296, 904–907. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; Garcia, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [Green Version]
- Navarro, L.M.; Pereira, H.M. Rewilding abandoned landscapes in Europe. In Rewilding European Landscapes; Springer: Berlin/Heidelberg, Germany, 2015; pp. 3–23. [Google Scholar]
- Brooks, T.M.; Mittermeier, R.A.; Fonseca, G.A.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newmark, W.D. Isolation of African protected areas. Front. Ecol. Environ. 2008, 6, 321–328. [Google Scholar] [CrossRef]
- Geldmann, J.; Joppa, L.N.; Burgess, N.D. Mapping change in human pressure globally on land and within protected areas. Conserv. Biol. 2014, 28, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Duffy, R. Global governance and environmental management: The politics of transfrontier conservation areas in Southern Africa. Political Geogr. 2006, 25, 89–112. [Google Scholar] [CrossRef]
- Gren, M.; Svensson, T.; Elofsson, K.; Engelmann, M. Economics of wildlife management—An overview. Eur. J. Wildl. Res. 2018, 64, 22. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The human footprint and the last of the wild: The human footprint is a global map of human influence on the land surface, which suggests that human beings are stewards of nature, whether we like it or not. Bioscience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Walker, B.; Salt, D. Resilience Thinking: Sustaining Ecosystems and People in a Changing World; Island Press: Washington, DC, USA, 2006; 192p, ISBN 978-1-59726-622-2. [Google Scholar]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Global terrestrial Human Footprint maps for 1993 and 2009. Sci. Data 2016, 3, 160067. [Google Scholar] [CrossRef] [Green Version]
- Ayram, C.A.C.; Mendoza, M.E.; Etter, A.; Salicrup, D.R.P. Anthropogenic impact on habitat connectivity: A multidimensional human footprint index evaluated in a highly biodiverse landscape of Mexico. Ecol. Indic. 2017, 895–909. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Shanahan, D.F.; Di Marco, M.; Allan, J.; Laurance, W.F.; Sanderson, E.W.; Mackey, B.; Venter, O. Catastrophic declines in wilderness areas undermine global environment targets. Curr. Biol. 2016, 26, 2929–2934. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.R.; Venter, O.; Fuller, R.A.; Allan, J.; Maxwell, S.L.; Negret, P.J.; Watson, J.E. One-third of global protected land is under intense human pressure. Science 2018, 360, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Gaynor, K.M.; Hojnowski, C.E.; Brashares, J.S. The influence of human disturbance on wildlife nocturnality. Science 2018, 360, 1232–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marco, M.; Venter, O.; Possingham, H.P.; Watson, J.E. Changes in human footprint drive changes in species extinction risk. Nat. Commun. 2018, 9, 4621. [Google Scholar] [CrossRef]
- Di Marco, M.; Santini, L. Human pressures predict species’ geographic range size better than biological traits. Glob. Chang. Biol. 2015, 21, 2169–2178. [Google Scholar] [CrossRef]
- Trombulak, S.C.; Baldwin, R.F.; Woolmer, G. The Human Footprint as a Conservation Planning Tool in Landscape-Scale Conservation Planning; Trombulak, S.C., Baldwin, R.F., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 281–302. [Google Scholar]
- Sutherland, W.J.; Pullin, A.S.; Dolman, P.M.; Knight, T.M. The need for evidence-based conservation. Trends Ecol. Evol. 2004, 19, 305–308. [Google Scholar] [CrossRef]
- Woolmer, G.; Trombulak, S.C.; Ray, J.C.; Doran, P.J.; Anderson, M.G.; Baldwin, R.F.; Morgan, A.; Sanderson, E.W. Rescaling the human footprint: A tool for conservation planning at an ecoregional scale. Landsc. Urban Plan. 2008, 87, 42–53. [Google Scholar] [CrossRef]
- Cumming, D.H. Constraints to conservation and development success at the wildlife-livestock-human interface in southern African transfrontier conservation areas: A preliminary review. Wildl. Conserv. Soc. 2011. Available online: https://www.povertyandconservation.info/en/biblio/B1785 (accessed on 20 October 2021).
- Wittemyer, G.; Elsen, P.; Bean, W.T.; Burton, A.C.; Brashares, J.S. Accelerated human population growth at protected area edges. Science 2008, 321, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 124–148. [Google Scholar] [CrossRef] [Green Version]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, B.C.J.; et al. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riggio, J.; Kija, H.; Masenga, E.; Mbwilo, F.; Van de Perre, F.; Caro, T. Sensitivity of Africa’s larger mammals to humans. J. Nat. Conserv. 2018, 43, 136–145. [Google Scholar] [CrossRef]
- Funston, P.J.; Groom, R.J.; Lindsey, P.A. Insights into the management of large carnivores for profitable wildlife-based land uses in African savannas. PLoS ONE 2013, 8, e59044. [Google Scholar] [CrossRef]
- KAZA. The Kavango-Zambezi Transfrontier Conservation Area Carnivore Conservation Strategy; KAZA: Lusaka, Zambia, 2018. [Google Scholar]
- KAZA. Master Integrated Development Plan; KAZA: Lusaka, Zambia, 2014. [Google Scholar]
- UNEP-WCMC. United Nations Environment Programmes World Conservation Monitoring Centre. Protected Planet Database. Available online: https://www.protectedplanet.net/ (accessed on 1 May 2015).
- Lindsey, P.; Nyirenda, V.; Barnes, J.; Becker, M.S.; McRoob, R. Underperformance of the Zambian protected area network: Steps needed to improve functionality and effectiveness. PLoS ONE 2014, 9, e94109. [Google Scholar]
- Lines, R.; Bormpoudakis, D.; Xofis, P.; Tzanopoulos, J. Modelling multi-species connectivity at the Kafue-Zambezi Interface: Implications for Transboundary Carnivore Conservation. Sustainability 2021, 13, 12886. [Google Scholar] [CrossRef]
- Lines, R.; Tzanopoulos, J.; MacMillan, D. Status of terrestrial mammals at the Kafue–Zambezi interface: Implications for transboundary connectivity. Oryx 2018, 53, 764–773. [Google Scholar] [CrossRef]
- KAZA. Treaty Between the Governments of Angola, Botswana, Namibia, Zambia and Zimbabwe on the Establishment of the Kavango-Zambezi Transfrontier Conservation Area. J. Environ. Dev. 2008, 17, 99–117. [Google Scholar]
- CSO. Central Statistics Office of Zambia. Available online: http://www.zamstats.gov.zm/ (accessed on 23 September 2019).
- Calvert, G.M. Sitimela: A History of the Zambesi Sawmills Logging Railway, 1911–1972; Barotse Development Trust: Mongu, Zambia, 2005. [Google Scholar]
- Rees, W.; Wackernagel, M. Our Ecological Footprint: Reducing Human Impact on the Earth; New Society Publishers: Gabriola Island, BC, Canada, 1996. [Google Scholar]
- Bonafilia, D.; Gill, J.; Basu, S.; Yang, D. Building High Resolution Maps for Humanitarian Aid and Development with Weakly-and Semi-Supervised Learning. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, CVPR Workshops 2019, Long Beach, CA, USA, 16–20 June 2019; pp. 1–9. [Google Scholar]
- NOAA. National Oceanic and Atmospheric Administration. National Geophysical Data Center. DMSP Data Collected by US Air Force Weather Agency; NOAA: Washington, DC, USA, 2019.
- Woodroffe, R.; Ginsberg, J.R. Edge Effects and the Extinction of Populations Inside Protected Areas. Science 1998, 280, 2126–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musgrave, M. Scale, Governance and Change in Zambezi Teak Forests: Sustainable Development for Commodity and Community; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2016. [Google Scholar]
- Rich, C.; Longcore, T. Ecological Consequences of Artificial Night Lighting; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modelling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Jackson, C.R.; Marnewick, K.; Lindsey, P.; Røskaft, E.; Robertson, M.P. Evaluating habitat connectivity methodologies: A case study with endangered African wild dogs in South Africa. Landsc. Ecol. 2016, 312, 1433–1447. [Google Scholar] [CrossRef]
- Di Minin, E.; Macmillan, D.C.; Goodman, P.S.; Escott, B.; Slotow, R.; Moilanen, A. Conservation businesses and conservation planning in a biological diversity hotspot. Conserv. Biol. 2013, 27, 808–820. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, M.; Nezami Balouchi, B.; Jowkar, H.; Hemami, M.; Fadakar, D.; Malakouti-Khah, S.; Ostrowski, S. Combining landscape suitability and habitat connectivity to conserve the last surviving population of cheetah in Asia. Divers. Distrib. 2017, 23, 592–603. [Google Scholar] [CrossRef] [Green Version]
- Angelieri, C.C.S.; Adams-Hosking, C.; de Barros Ferraz, K.M.P.M.; de Souza, M.P.; McAlpine, C.A. Using species distribution models to predict potential landscape restoration effects on puma conservation. PLoS ONE 2016, 11, e0145232. [Google Scholar]
- Di Marco, M.; Rondinini, C.; Boitani, L.; Murray, K. Comparing multiple species distribution proxies and different quantifications of the human footprint map, implications for conservation. Biol. Conserv. 2013, 165, 203–211. [Google Scholar] [CrossRef]
- Leu, M.; Hanser, S.E.; Knick, S.T. The human footprint in the west: A large-scale analysis of anthropogenic impacts. Ecol. Appl. 2008, 18, 1119–1139. [Google Scholar] [CrossRef]
- Everatt, K.T.; Moore, J.; Kerley, G.I. Africa’s apex predator, the lion, is limited by interference and exploitive competition with humans. Glob. Ecol. Conserv. 2019, 20, e00758. [Google Scholar] [CrossRef]
- Brodie, J.F. Synergistic effects of climate change and agricultural land use on mammals. Front. Ecol. Environ. 2016, 14, 20–26. [Google Scholar] [CrossRef]
- Halpern, B.S.; Fujita, R. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Toews, M. Managing Human Footprint with Respect to its Effects on Large Mammals: Implications of Spatial Scale, Divergent Responses and Ecological Thresholds; University of Victoria: Victoria, BC, Canada, 2016. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Variable | Pressure Score | Source | Spatial Resolution | Details |
|---|---|---|---|---|
| Settlement | 0, 10 | [43] | 30 m | All settled areas mapped given score of 10 |
| Population Density | 0–10 Continuous | [43] | 30 m | Pressure score = 3.333 × log (population density + 1) |
| Roads | 8 Direct impacts 0–4 Indirect impacts | PPF | 10 m | Direct pressure score of 8 for 500 m either side of road, exponentially decaying out to 4 at 15 km |
| Railways | 0–8 | PPF | 10 m | Direct pressure score of 8 for 500 m either side |
| Navigable Water | 0–4 | PPF | 10 m | Pressure score of 4 exponentially decaying out to 15 km |
| Arable | 0, 7 | [37] | 10 m | All areas mapped as crops given score of 7 |
| Night Lights/NTL | 0–9 | [44] | 100 m | Pressure score = 3.333 × log (NTL + 1) |
| Area | IUCN | Area Ha | Mean HFP | Occurrence | ||
|---|---|---|---|---|---|---|
| Leopard | Lion | Hyena | ||||
| Kafue National Park | II | 206,314 | 1.1 | Yes | Yes | Yes |
| Mulobezi GMA | VI | 347,481 | 1.2 | Yes | Yes | Yes |
| Sichifulo GMA | VI | 133,734 | 2.2 | Yes | Yes | Yes |
| Nachitwe Forest Reserve | unreported | 71,075 | 2.3 | Yes | Yes | Yes |
| Martin Forest Reserve | unreported | 62,948 | 2.4 | Yes | Yes | Yes |
| Nyawa West | unreported | 57,439 | 3.7 | Yes | No | Yes |
| Simalaha Conservancy | unreported | 181,936 | 4.6 | No | No | No |
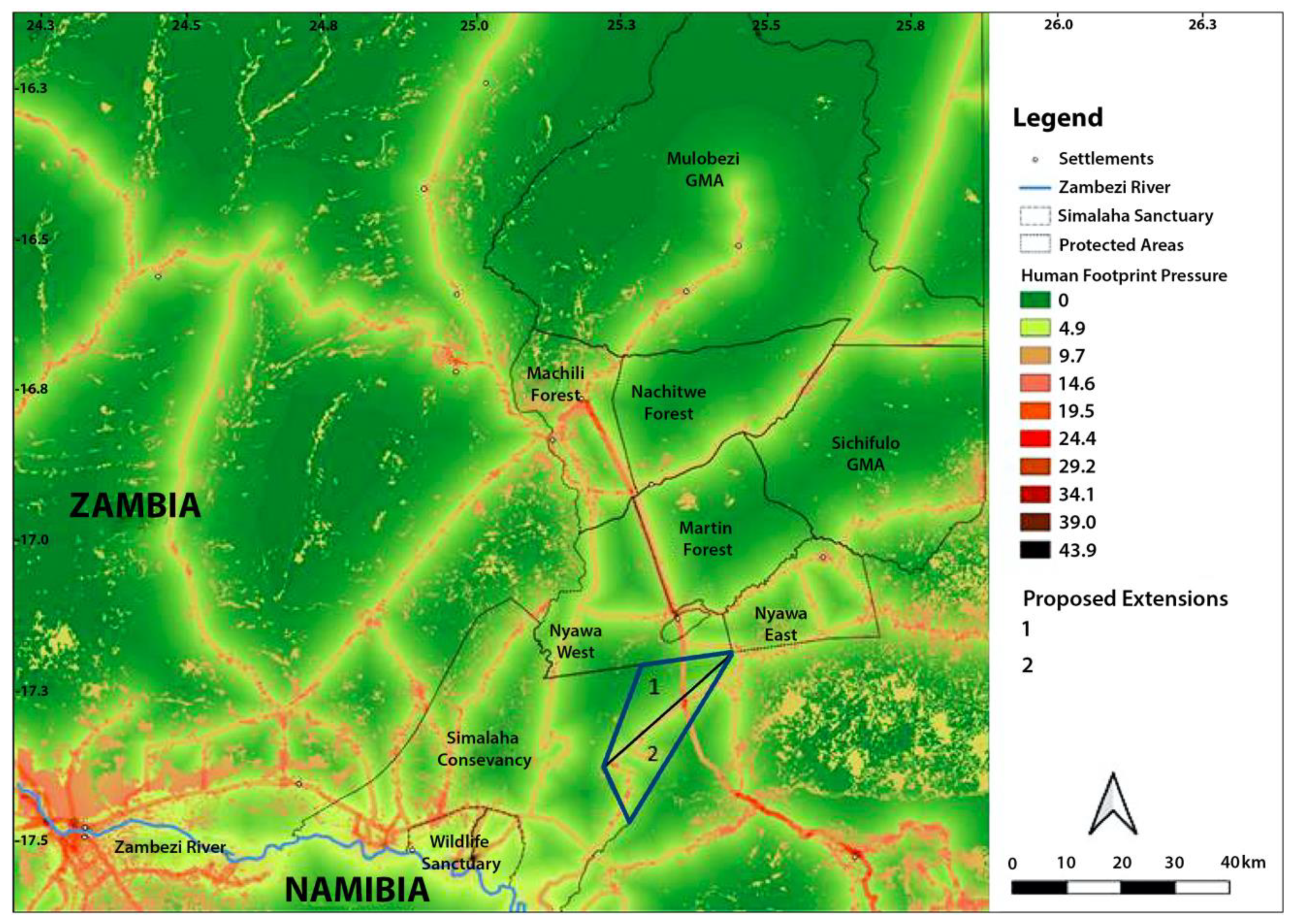
| Open Area extension 1 * | unreported | 32,712 | 3.9 | Yes | No | Yes |
| Open Area extension 1 + 2 * | unreported | 73,660 | 3.9 | Yes | No | Yes |
| Nyawa East | unreported | 39,565 | 4.6 | No | No | No |
| Machili Forest Reserve | unreported | 49,269 | 5.6 | Yes | No | Yes |
| Simalaha Sanctuary E | unreported | 11,020 | 5.8 | No | No | No |
| Simalaha Sanctuary W | unreported | 11,282 | 6.8 | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lines, R.; Bormpoudakis, D.; Xofis, P.; MacMillan, D.C.; Pieterse, L.; Tzanopoulos, J. Utility of Human Footprint Pressure Mapping for Large Carnivore Conservation: The Kafue-Zambezi Interface. Sustainability 2022, 14, 116. https://doi.org/10.3390/su14010116
Lines R, Bormpoudakis D, Xofis P, MacMillan DC, Pieterse L, Tzanopoulos J. Utility of Human Footprint Pressure Mapping for Large Carnivore Conservation: The Kafue-Zambezi Interface. Sustainability. 2022; 14(1):116. https://doi.org/10.3390/su14010116
Chicago/Turabian StyleLines, Robin, Dimitrios Bormpoudakis, Panteleimon Xofis, Douglas C. MacMillan, Lucy Pieterse, and Joseph Tzanopoulos. 2022. "Utility of Human Footprint Pressure Mapping for Large Carnivore Conservation: The Kafue-Zambezi Interface" Sustainability 14, no. 1: 116. https://doi.org/10.3390/su14010116