
Environmental Drivers and Social Structure Features behind the Low Reproductive Success of Dusky Groupers Epinephelus marginatus (Lowe, 1834) in a Mediterranean Marine Protected Area



Abstract
:1. Introduction
2. Materials and Methods
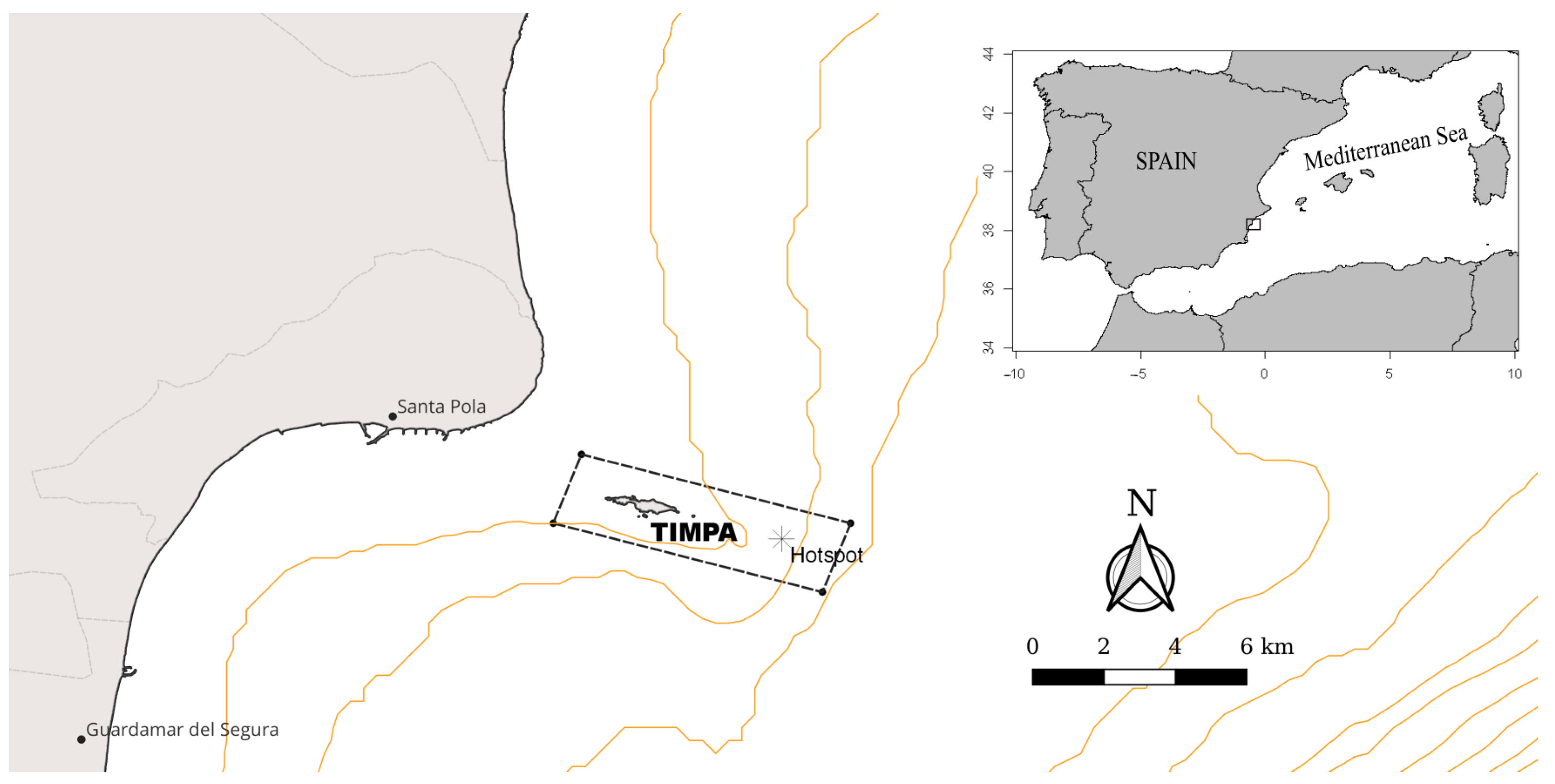
2.1. Sampling Area and Experimental Design
- Environmental drivers:
- The seabed temperature per day (°C). Temperature data were recorded each hour by a HOBO Pendant UA 002-64 data logger placed at 70 cm up the seabed and averaged per day;
- The thermocline, estimated by sea surface–bed temperature difference. Large differences were linked to a stable thermocline, whereas small differences to a weak or nonexistent thermocline;
- The visibility was established qualitatively by the sampler as bad (<10 m), moderate (from 10 to 20 m) or good visibility (>30 m);
- The stream force over the seabed was classified the same way as the previous variable: (1) no stream, (2) moderate, (3) strong and (4) very strong stream;
- The photoperiod or day length in hours at the hotspot position each survey-day. These data were consulted from the National Meteorological Agency (AEMET);
- The number of recreational divers who visited the hotspot per survey-day which was obtained from the MPA surveillance enterprise.
- Social structure and behavior variables:
- The density or specimen number per survey;
- The size class. Groupers were classified into three size classes, smalls (30 to 50 cm), mediums (50 to 80 cm) and larges (>80 cm). The size classification is strongly linked to sexual maturation [24];
- The group size. We classified the size group where the sighted grouper was included in (1) an alone specimen, (2) small (2 to 5 specimens), (3) medium (6 to 15 specimens) and (4) large (16 to 30 specimens) groups;
- The behavior. Its classification was summarized, as the pattern color, from those reported in the literature cited. It consisted of (1) non-sexual behavior, (2) pursuit or escape, (3) territory patrol, (4) female showing reproductive behavior and (5) male in courtship;
- The reproductive status. This qualitative variable indicated the sexual state of the specimen when it was sighted and only admitted two states: (1) reproductive or (2) non-reproductive status. It was determined by the behavior and the color pattern;
- The position. Specimen position was determined as (1) swimming, (2) motionless, resting at the water column, (3) hidden or (4) perched on the seabed;
- The belly. A swollen belly was representative of a female with matured eggs; we noticed if it was (1) swollen or (2) flat.
2.2. Statistical Analysis
3. Results
3.1. Temporal Pattern of Dusky Grouper Reproductive Behavior
3.2. Relationship of Reproductive Behavior with Environmental Drivers
3.3. Effect of Social Structure on Reproductive Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Rodríguez, D. Marine Protected Areas: Attempting the sustainability of the seas. In World Seas: An Environmental Evaluation; Sheppard, C., Ed.; Volume III: Ecological Issues and Environmental Impacts; Academic Press: Cambridge, MA, USA, 2019; pp. 475–489. [Google Scholar]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Salm, R.V.; Clark, J.; Siirila, E. Marine and Coastal Protected Areas: A Guide for Planners and Managers; IUCN: Washington, DC, USA, 2000; 371p. [Google Scholar]
- Cheng, B.S.; Altieri, A.H.; Torchin, M.E.; Ruiz, G.M. Can marine reserves restore lost ecosystem functioning? Glob. Synth. Ecol. 2019, 100, e02617. [Google Scholar]
- Blyth-Skyrme, R.; Kaiser, M.J.; Hiddink, J.G.; Edwards-Jones, G.; Hart, P.J.B. Conservation benefits of temperate marine protected areas: Variation among fish species. Conserv. Biol. 2006, 20, 811–820. [Google Scholar] [CrossRef]
- Chauvet, C. Statut d’ Epinephelus guaza (Linnaeus, 1758) et éléments de dynamique des populations méditerranée et atlantique. In Les Espèces Marines à Protéger en Méditerranée; Boudouresque, C.F., Avon, M., Graves, V., Eds.; GIS Posidonie: Marseille, France, 1991; pp. 255–275. [Google Scholar]
- Sadovy de Mitcheson, Y.; Craig, M.T.; Bertoncini, A.A.; Carpenter, K.E.; Cheung, W.W.L.; Choat, J.H.; Cornish, A.S.; Fennessy, S.T.; Ferreira, B.P.; Heemstra, P.C.; et al. Fishing groupers towards extinction: A global assessment of threats and extinction risks in a billion dollar fishery. Fish Fish. 2013, 14, 119–136. [Google Scholar] [CrossRef]
- Bohnsack, J.A. Marine reserves: They enhance fisheries, reduce conflicts, and protect resources. Oceanus 1993, 36, 63–70. [Google Scholar]
- Bohnsack, J.A.; Ault, J.S. Management Strategies to Conserve Marine Biodiversity. Oceanography, 1996, Volume 9, No. 1, Marine Biological Diversity: A Special Issue Commemorating 25 Years of Science and Service; National Oceanic and Atmospheric Administration (United States Department of Commerce): Washington, DC, USA, 1996; pp. 73–82.
- Harmelin-Vivien, M.L.; Le Direach, J.T.; Bayle-Sempere, E.; Charbonnel, J.A.; García-Charton, D.; Ody, A.; Pérez-Rusafa, O.; Reñones, P.; Sánchez-Jerez, C.V. Gradients of abundance and biomass in six Mediterranean marine protected areas: Evidence of fish spillover? Biol. Conserv. 2008, 141, 1829–1839. [Google Scholar] [CrossRef]
- Sánchez-Lizaso, J.L.; Goñi, R.; Reñones, O.; García-Charton, J.A.; Galzin, R.; Bayle-Sempere, T.; Sánchez-Jerez, P.; Pérez-Ruzafa, A.; Ramos, A. Density dependence in marine protected populations: A review. Environ. Conserv. 2000, 27, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, M.; Guidetti, P.; Franco, A.D.; Calo‘, A.; Claudet, J. Assessing spillover from marine protected areas and its drivers: A meta-analytical approach. Fish Fish. 2020, 21, 906–915. [Google Scholar] [CrossRef]
- Planes, S.; Galzin, R.; Rubies, A.; Goñi, R.; Harmelin, J.G.; LeDiréach, L.; Lenfant, P.; Quetglas, A. Effects of marine protected areas on recruitment processes with special reference to Mediterranean littoral ecosystems. Environ. Conserv. 2000, 27, 126–143. [Google Scholar] [CrossRef]
- Schunter, C.; Carreras-Carbonell, J.; Planes, S.; Sala, E.; Ballesteros, E.; Zabala, M.; Harmelin, J.G.; Harmelin-Vivien, M.; Macpherson, E.; Pascual, M. Genetic connectivity patterns in an endangered species: The dusky grouper (Epinephelus marginatus). J. Exp. Mar. Biol. Ecol. 2011, 401, 126–133. [Google Scholar] [CrossRef]
- Heemstra, P.C.; Randall, J.E. FAO species catalog. Groupers of the world (Family Serranidae, Subfamily Epinephelinae). An annotated and illustrated catalog of the grouper, rock cod, hind, coral grouper and lyretail species known to date. FAO Fish. Synop. 1993, 16, 382. [Google Scholar]
- Condini, M.C.; García-Charton, J.A.; García, A.M. A review of the biology, ecology, behavior and conservation status of the dusky grouper, Epinephelus marginatus (Lowe, 1834). Rev. Fish. Biol. Fish. 2018, 28, 301–330. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. 2022. Available online: https://www.iucnredlist.org (accessed on 3 May 2022).
- Zabala, M.; García-Rubies, A.; Louisy, P.; Sala, E. Spawning behavior of the Mediterranean dusky grouper Epinephelus marginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean, Spain). Sci. Mar. 1997, 61, 65–77. [Google Scholar]
- Zabala, M.; Louisy, P.; García-Rubiés, A.; Gracia, V. Socio-behavioral context of reproduction in the Mediterranean dusky grouper Epinephelus marginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean, Spain). Sci. Mar. 1997, 61, 79–89. [Google Scholar]
- Louisy, P.; Culioli, J.-M. Review of present knowledge on the reproductive activity of the dusky grouper Epinephelus marginatus (Lowe, 1834) in the North-Western Mediterranean. Mar. Life 1999, 9, 47–57. [Google Scholar]
- Bertucci, F.; Lejeune, P.; Payrot, J.; Parmentier, E. Sound production by dusky grouper Epinephelus marginatus at spawning aggregation sites. J. Fish Biol. 2015, 87, 400–421. [Google Scholar] [CrossRef]
- Pelaprat, C. Observations on the spawning behabiour of the dusky grouper Epinephelus marginatus (Lowe, 1834) in the north of Corsica (France). Mar. Life 1999, 9, 59–65. [Google Scholar]
- Marinaro, J.Y.; Roussel, E.; Lawson, J.; Crec’hriou, R.; Planes, S. Premier signalement d’une reproduction effective de mérou brun, Epinephelus marginatus, dans la Réserve marine de Cerbére-Banyuls (France). Cybium 2005, 29, 198–200. [Google Scholar]
- Reñones, O.; Grau, A.; Mas, X.; Riera, F.; Saborido-Rey, F. Reproductive pattern of an exploited dusky grouper Epinephelus marginatus (Lowe 1834) (Pisces: Serranidae) population in the western Mediterranean. Sci. Mar. 2010, 74, 523–537. [Google Scholar] [CrossRef] [Green Version]
- Marino, G.; Azzurro, E.; Massari, A.; Finoia, M.G.; Mandich, A. Reproduction in the dusky grouper from the southern Mediterranean. J. Fish Biol. 2001, 58, 909–927. [Google Scholar] [CrossRef]
- Özen, M.R.; Balci, B.A. Reproductive characteristics of dusky goruper (Epinephelus guaza, Linnaeus 1758) in Antalya Bay of Eastern Mediterranean. Pak. Vet. J. 2011, 31, 215–218. [Google Scholar]
- Koeck, B.; Pastor, J.; Saragoni, G.; Dalias, N.; Payrot, J.; Lenfant, P. Diel and seasonal movement pattern of the dusky grouper Epinephelus marginatus inside a marine reserve. Mar. Environ. Res. 2014, 94, 38–47. [Google Scholar] [CrossRef]
- Desiderà, E.; Trainito, E.; Navone, A.; Blandin, R.; Magnani, L.; Panzalis, P.; Mazzoldi, C.; Guidetti, P. Using complementary visual approaches to investigate residency, site fidelity and movement patterns of the dusky grouper (Epinephelus marginatus) in a Mediterranean marine protected area. Mar. Biol. 2021, 168, 111. [Google Scholar] [CrossRef]
- Hereu, B.; Diaz, D.; Paqüal, J.; Zabala, M.; Sala, E. Temporal patterns of spawning of the dusky grouper Epinephelus marginatus in relation to environmental factors. Mar. Ecol. Prog. Ser. 2006, 325, 187–194. [Google Scholar] [CrossRef]
- Félix-Hackradt, F.C. Chapter 1.2 Early life development. In Biology and Ecology of Groupers; Félix-Hackradt, F.C., Hackradt, C.W., García-Charton, J.A., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 2022; pp. 85–110. [Google Scholar]
- Revenga, S.; Badalamenti, F. Management of marine protected areas for fisheries in the Mediterranean. In The Mediterranean Fisheries Sector. A Reference Publication for the VII Meeting of Ministers of Agriculture and fisheries of CIHEAM Member Countries (Zaragoza, Spain, 4 February 2008); Options Méditerranéennes: Série B. Etudes et Recherches, n. 62; Basurco, B., Ed.; CIHEAM: Paris, France; FAO: Rome, Italy; GFCM: Zaragoza, Spain, 2008; pp. 107–111. [Google Scholar]
- Forcada, A.; Bayle-Sempere, J.T.; Valle, C.; Sánchez-Jerez, P. Habitat continuity effects on gradients of fish biomass across marine protected area boundaries. Mar. Environ. Res. 2008, 66, 536–547. [Google Scholar] [CrossRef] [Green Version]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Chauvet, C.; Duval, C.; Galzin, R.; Lejeune, P.; Barnabé, G.; Blanc, F.; Chevalier, R.; Duclerc, J.; et al. Evaluation des peuplements et populations de poissons. Méthodes et problèmes. Rev. Ecol. (Terre Vie) 1985, 40, 467–539. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology; R. Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Fournier, D.A.; Skaug, H.J.; Ancheta, J.; Ianelli, J.; Magnusson, A.; Maunder, M.N.; Nielsen, A.; Sibert, J. AD model builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim Methods Softw. 2012, 27, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Skaug, H.; Fournier, D.; Nielsen, A.; Magnusson, A.; Bolker, B. Generalized Linear Mixed Models Using ‘AD Model Builder’. R Package Version 0.8.3.3. 2016. Available online: https://rdrr.io/rforge/glmmADMB/ (accessed on 18 October 2021).
- R Core and Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://cran.r-project.org/ (accessed on 15 October 2021).
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Crawley, M.J. Statistical Computing: An Introduction to Data Analysis Using S-Plus; John Wiley & Sons: Chichester, UK, 2003; 772p. [Google Scholar]
- Bolker, B.M. Ecological Models and Data in R; Princeton University Press: Princeton, NJ, USA, 2008; 408p. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons Inc.: Chichester, UK, 2012; 1080p. [Google Scholar]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef]
- Pastor, F.; Valiente, J.A.; Palau, J.L. Sea surface temperature in the Mediterranean: Trends and spatial patterns (1982–2016). Pure Appl. Geophys. 2018, 175, 4017–4029. [Google Scholar] [CrossRef] [Green Version]
- Andrade, A.B.; Machado, L.F.; Hostim-Silva, M.; Barreiros, J.P. Reproductive biology of the dusky grouper Epinephelus marginatus (Lowe, 1834). Braz. Arch. Biol. Technol. 2003, 46, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Colin, P.L. Reproduction of the Nassau grouper, Epinephelus striatus (Pisces: Serranidae) and its relationship to environmental conditions. Environ. Biol. Fishes 1992, 34, 357–377. [Google Scholar] [CrossRef]
- Watanabe, W.O.; Lee, C.S.; Ellis, S.C.; Ellis, E.P. Hatchery study of the effects of temperature on eggs and yolk sac larvae of the Nassau grouper Epinephelus striatus. Aquaculture 1995, 136, 141–147. [Google Scholar] [CrossRef]
- Das, S.K.; Xiang, T.W.; Noor, N.M.; De, M.; Mazumder, S.K.; Goutham-Bharathi, M.P. Temperature physiology in grouper (Epinephelinae: Serranidae) aquaculture: A brief review. Aquac. Rep. 2021, 20, 1000682. [Google Scholar] [CrossRef]
- Bruton, M.N. The effects of suspensoids on fish. Hydrobiologia 1985, 125, 221–241. [Google Scholar] [CrossRef]
- Hardie, S.A. Hydrological manipulation to assist spawning of a threatened galaxiid fish in a highland lake system. Mar. Freshw. Res. 2013, 64, 887–899. [Google Scholar] [CrossRef]
- Skaramuca, B.; Musin, D.; Onofri, V.; Caric, M. A contribution to the knowledge on the spawning time of the dusky grouper (Epinephelus guaza L.). Acta Biol. Iugosl. E. Ichthyol. 1989, 21, 79–85. [Google Scholar]
- Pérez-Ruzafa, A.; Martín, E.; Marcos, C.; Zamarro, J.M.; Stobart, B.; Harmelin-Vivien, M.; Polti, S.; Planes, S.; García-Charton, J.A.; González-Wangüemert, M. Modelling spatial and temporal scales for spill-over and biomass exportation from MPAs and their potential for fisheries enhancement. J. Nat. Conserv. 2008, 16, 234–255. [Google Scholar] [CrossRef]
- Ohayon, S.; Granot, I.; Belmaker, J. A meta-analysis reveals edge effects within marine protected areas. Nat. Ecol. Evol. 2021, 5, 1301–1308. [Google Scholar] [CrossRef]
- Salas, S.; Chuenpagdee, R.; Barragán-Paladines, M.J. Drivers and Prospects For The Sustainability And Viability Of Small-Scale Fisheries in Latin America and the Caribbean. In Viability and Sustainability of Small-Scale Fisheries in Latin America and the Caribbean; Salas, S., Barragán-Paladines, M.J., Chenpagdee, R., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 543–559. [Google Scholar]
- Grafton, R.Q.; Kompas, T. Uncertainty and the active adaptative management of marine reserves. Mar. Policy 2005, 29, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Nickols, K.J.; White, J.W.; Malone, D.; Carr, M.H.; Starr, R.M.; Baskett, M.L.; Hastings, A.; Botsford, L.W. Setting ecological expectations for adaptive management of marine protected areas. J. Appl. Ecol. 2019, 56, 2376–2385. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Estimate | Std. Error | Exp. Estimate | Lower 95 | Upper 95 | p-Value |
|---|---|---|---|---|---|---|
| Intercept | −0.158 | 0.305 | 0.853 | 0.470 | 1.550 | 0.603 |
| S(T) | 0.794 | 0.159 | 2.213 | 1.622 | 3.021 | <0.001 |
| S(The) | 1.045 | 0.170 | 2.844 | 2.037 | 3.971 | <0.001 |
| Moderate V | −0.127 | 0.206 | 0.881 | 0.589 | 1.319 | 0.538 |
| Good V | 0.190 | 0.0289 | 1.209 | 0.687 | 2.130 | 0.510 |
| S(Ph) | 0.214 | 0.117 | 1.239 | 0.985 | 1.558 | 0.067 |
| S(T):S(The) | 0.236 | 0.151 | 1.267 | 0.942 | 1.703 | 0.118 |
| S(T):Moderate V | −1.338 | 0.271 | 0.262 | 0.154 | 0.446 | <0.001 |
| S(T):Good V | 1.072 | 0.408 | 2.920 | 1.313 | 6.492 | 0.009 |
| S(The):Moderate V | −1.049 | 0.244 | 0.350 | 0.217 | 0.566 | <0.001 |
| S(The):Good V | −0.435 | 0.521 | 0.647 | 0.233 | 1.798 | 0.404 |
| S(T):Scale(The):Moderate V | −0.672 | 0.310 | 0.511 | 0.278 | 0.938 | 0.03 |
| S(T):Scale(The):Good V | 1.351 | 0.534 | 3.861 | 1.356 | 10.995 | 0.011 |
| AIC | 1205.394 | |||||
| Sd. sites (Intercept) | 0.725 |
| Variables | Estimate | Std. Error | Exp. Estimate | Lower 95 | Upper 95 | p-Value |
|---|---|---|---|---|---|---|
| Intercept | −0.938 | 0.277 | 0.391 | 0.288 | 0.673 | 0.001 |
| Medium SC | 0.524 | 0.358 | 1.690 | 0.837 | 3.410 | 0.143 |
| Large SC | 1.360 | 0.240 | 3.897 | 2.433 | 6.243 | <0.001 |
| MotionlessP | −0.276 | 0.170 | 0.759 | 0.543 | 1.060 | 0.106 |
| HiddenP | −2.659 | 0.611 | 0.070 | 0.021 | 0.232 | <0.001 |
| PerchedP | −3.143 | 0.471 | 0.043 | 0.017 | 0.109 | <0.001 |
| Small SG | −1.078 | 0.213 | 0.340 | 0.224 | 0.517 | <0.001 |
| Medium SG | −2.011 | 0.498 | 0.134 | 0.050 | 0.355 | <0.001 |
| Large SG | −13.805 | 479.98 | 0 | 0 | ∞ | 0.977 |
| AIC | 1131.382 | |||||
| Sd. sites (Intercept) | 0.404 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano-Quijada, F.; González-Correa, J.M.; Bayle-Sempere, J.T. Environmental Drivers and Social Structure Features behind the Low Reproductive Success of Dusky Groupers Epinephelus marginatus (Lowe, 1834) in a Mediterranean Marine Protected Area. Sustainability 2022, 14, 6169. https://doi.org/10.3390/su14106169
Lozano-Quijada F, González-Correa JM, Bayle-Sempere JT. Environmental Drivers and Social Structure Features behind the Low Reproductive Success of Dusky Groupers Epinephelus marginatus (Lowe, 1834) in a Mediterranean Marine Protected Area. Sustainability. 2022; 14(10):6169. https://doi.org/10.3390/su14106169
Chicago/Turabian StyleLozano-Quijada, Felio, José Miguel González-Correa, and Just Tomàs Bayle-Sempere. 2022. "Environmental Drivers and Social Structure Features behind the Low Reproductive Success of Dusky Groupers Epinephelus marginatus (Lowe, 1834) in a Mediterranean Marine Protected Area" Sustainability 14, no. 10: 6169. https://doi.org/10.3390/su14106169
APA StyleLozano-Quijada, F., González-Correa, J. M., & Bayle-Sempere, J. T. (2022). Environmental Drivers and Social Structure Features behind the Low Reproductive Success of Dusky Groupers Epinephelus marginatus (Lowe, 1834) in a Mediterranean Marine Protected Area. Sustainability, 14(10), 6169. https://doi.org/10.3390/su14106169