
Responses of Vegetation Autumn Phenology to Climatic Factors in Northern China

Abstract
:1. Introduction
2. Materials and Methods
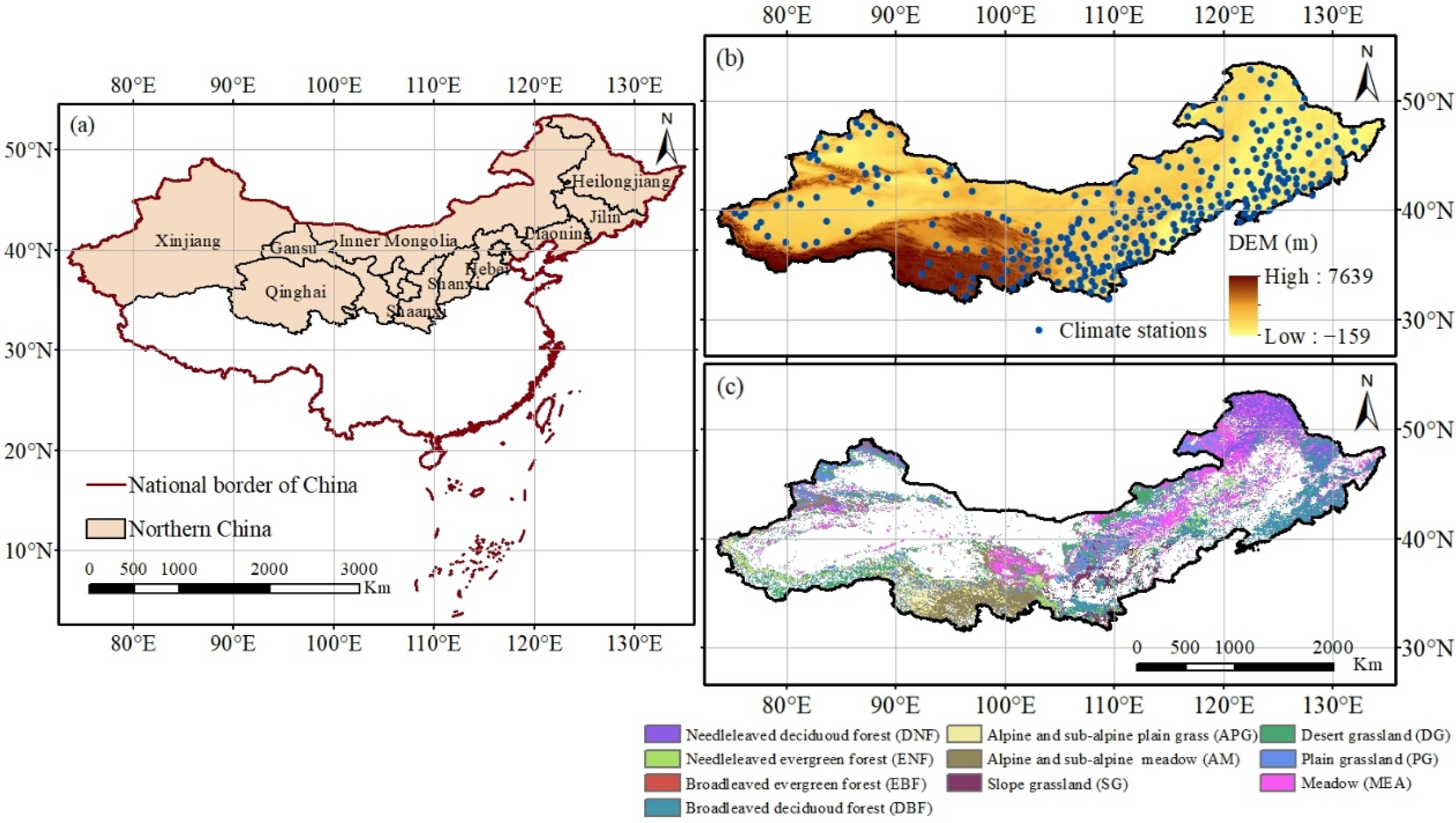
2.1. Study Region
2.2. Data
2.3. Methods
2.3.1. Retrieval of the EOS from EVI Data
2.3.2. Interannual Variation of EOS
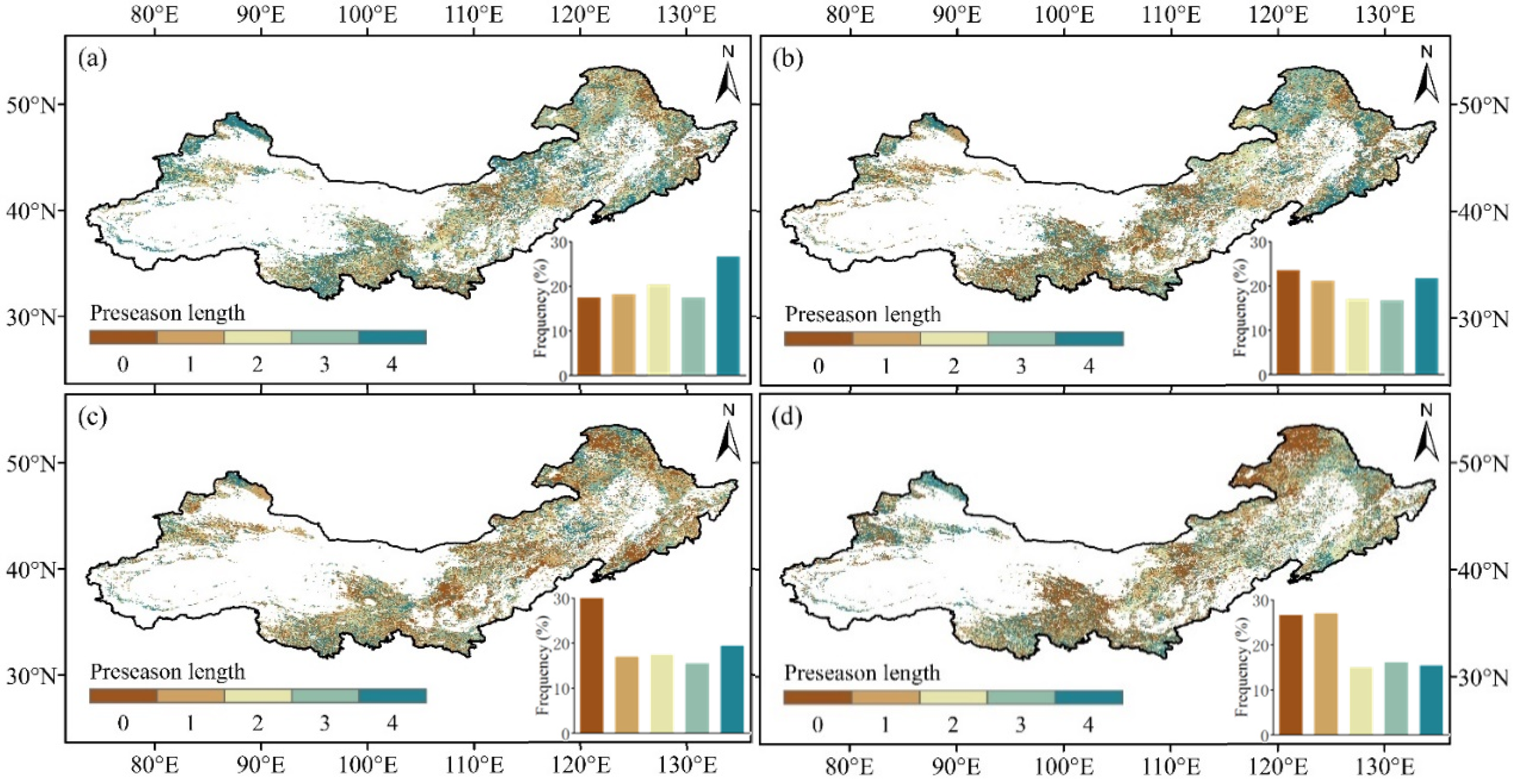
2.3.3. Determination of the Length of Preseason Climatic Factors
2.3.4. Multivariate Regression Analysis
3. Results
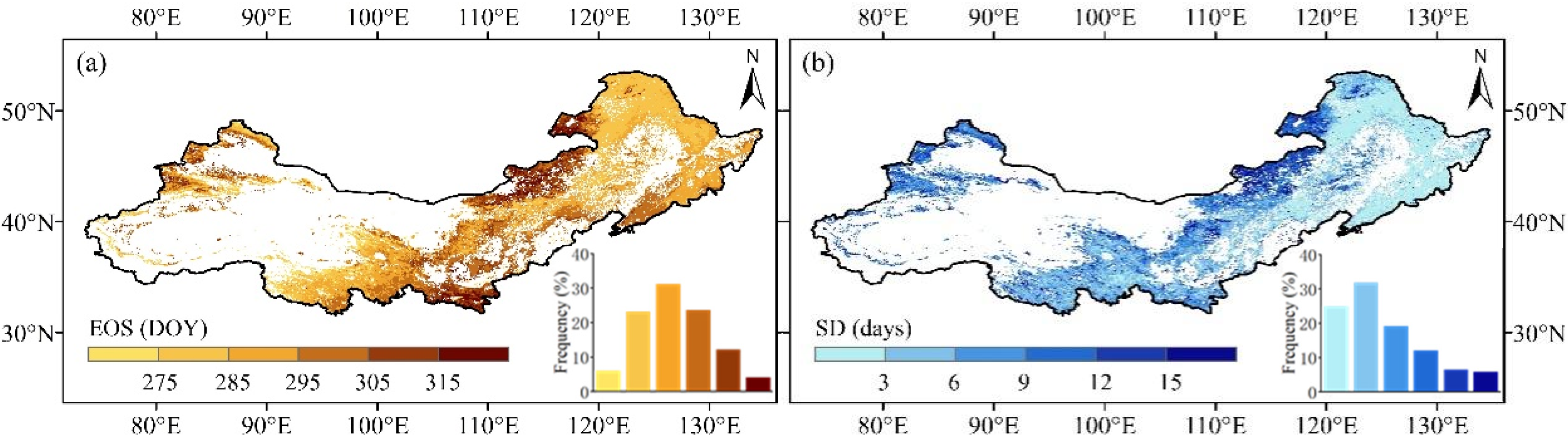
3.1. Spatial Distributions of EOS in Northern China
3.2. Inter-Annual Variations of EOS in Northern China
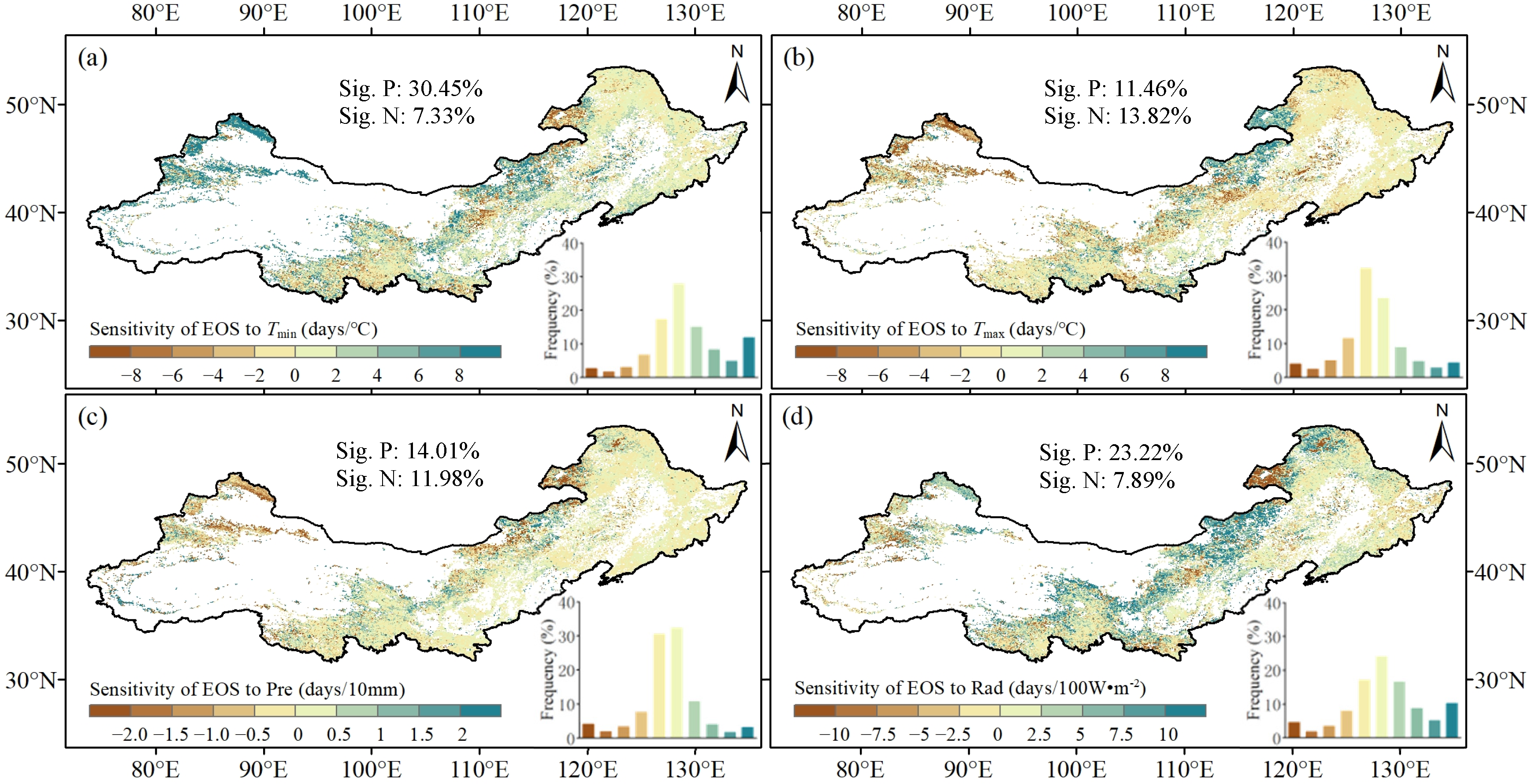
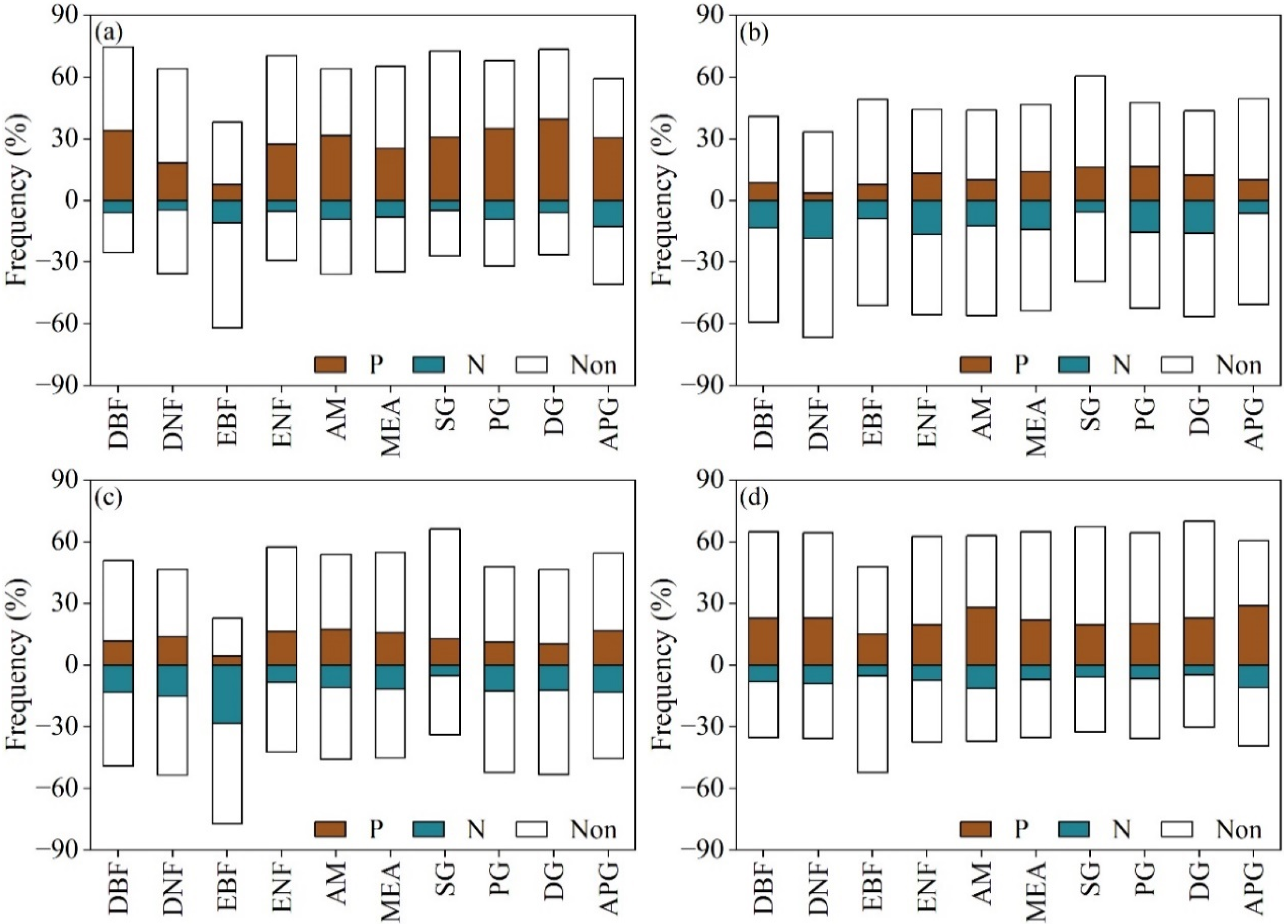
3.3. The Sensitivity of EOS to Climatic Factors in Northern China
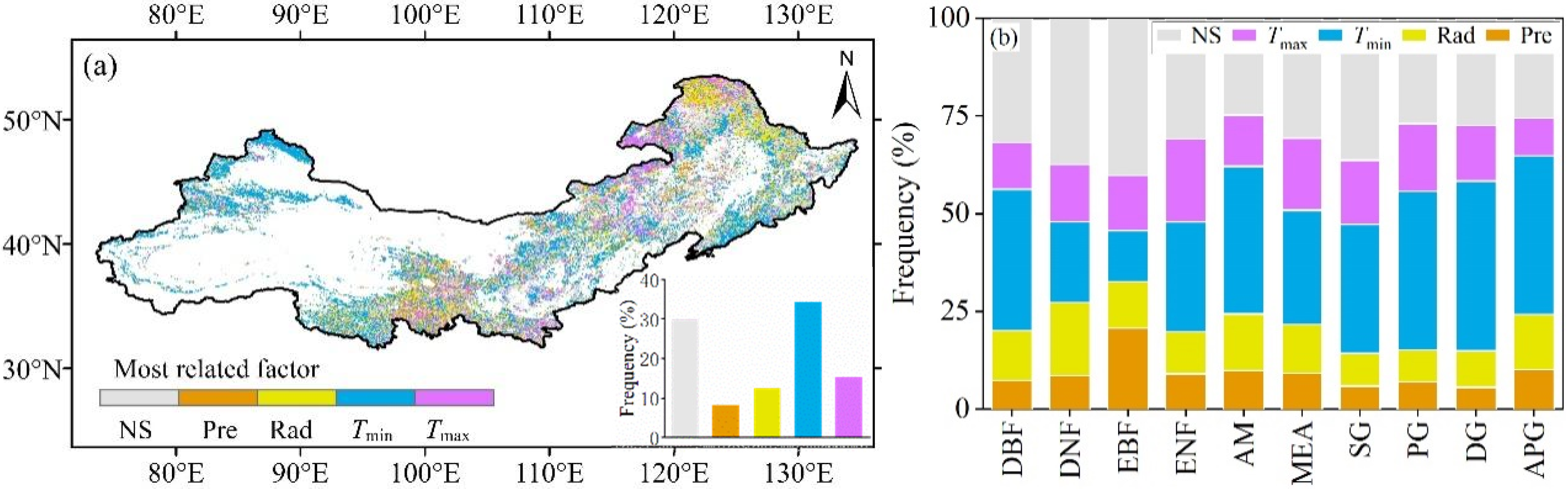
3.4. Determining the Key Climatic Factor driving EOS in Northern China
4. Discussion
4.1. Comparisons of EOS Trends in Different Regions
4.2. Comparison between the Impacts of Different Climatic Factors on EOS
4.3. Limitations and Future Work
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Aubrecht, D.M.; Furze, M.E.; Seyednasrollah, B.; Krassovski, M.B.; Latimer, J.M.; Nettles, W.R.; Heiderman, R.R.; et al. Ecosystem Warming Extends Vegetation Activity but Heightens Vulnerability to Cold Temperatures. Nature 2018, 560, 368–371. [Google Scholar] [CrossRef]
- Xia, J.; Niu, S.; Ciais, P.; Janssens, I.A.; Chen, J.; Ammann, C.; Arain, A.; Blanken, P.D.; Cescatti, A.; Bonal, D.; et al. Joint Control of Terrestrial Gross Primary Productivity by Plant Phenology and Physiology. Proc. Natl. Acad. Sci. USA 2015, 112, 2788–2793. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X. Legacy Effect of Spring Phenology on Vegetation Growth in Temperate China. Agric. For. Meteorol. 2020, 281, 107845. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Thompson, J.R. Climate Change Imposes Phenological Trade-Offs on Forest Net Primary Productivity. J. Geophys. Res. Biogeosci. 2017, 122, 2298–2313. [Google Scholar] [CrossRef]
- Zeng, Z.; Piao, S.; Li, L.Z.X.; Zhou, L.; Ciais, P.; Wang, T.; Li, Y.; Lian, X.; Wood, E.F.; Friedlingstein, P.; et al. Climate Mitigation from Vegetation Biophysical Feedbacks during the Past Three Decades. Nat. Clim. Chang. 2017, 7, 432–436. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hänninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger Temperature Response of Autumn Leaf Senescence than Spring Leaf-out Phenology. Glob. Chang. Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef]
- Peng, J.; Wu, C.; Wang, X.; Lu, L. Spring Phenology Outweighed Climate Change in Determining Autumn Phenology on the Tibetan Plateau. Int. J. Climatol. 2021, 41, 3725–3742. [Google Scholar] [CrossRef]
- Dannenberg, M.P.; Wise, E.K.; Janko, M.; Hwang, T.; Smith, W.K. Atmospheric Teleconnection Influence on North American Land Surface Phenology. Environ. Res. Lett. 2018, 13, 034029. [Google Scholar] [CrossRef]
- Li, C.; Wang, R.; Cui, X.; Wu, F.; Yan, Y.; Peng, Q.; Qian, Z.; Xu, Y. Responses of Vegetation Spring Phenology to Climatic Factors in Xinjiang, China. Ecol. Indic. 2021, 124, 107286. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring Vegetation Phenology Using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Rozenstein, O.; Adamowski, J. Linking Spaceborne and Ground Observations of Autumn Foliage Senescence in Southern Québec, Canada. Remote Sens. 2017, 9, 630. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gao, Q.; Wang, C.; Yu, M. Spatiotemporal Patterns of Vegetation Phenology Change and Relationships with Climate in the Two Transects of East China. Glob. Ecol. Conserv. 2017, 10, 206–219. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Piao, S. Spatial Variations in Responses of Vegetation Autumn Phenology to Climate Change on the Tibetan Plateau. J. Plant Ecol. 2016, 10, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Wu, C.; Gu, C.; Wang, X. Evidence for Satellite Observed Changes in the Relative Influence of Climate Indicators on Autumn Phenology over the Northern Hemisphere. Glob. Planet. Chang. 2020, 187, 103131. [Google Scholar] [CrossRef]
- Wang, X.; Xiao, J.; Li, X.; Cheng, G.; Ma, M.; Zhu, G.; Altaf Arain, M.; Andrew Black, T.; Jassal, R.S. No Trends in Spring and Autumn Phenology during the Global Warming Hiatus. Nat. Commun. 2019, 10, 2389. [Google Scholar] [CrossRef]
- Li, P. Combined Control of Multiple Extreme Climate Stressors on Autumn Vegetation Phenology on the Tibetan Plateau under Past and Future Climate Change. Agric. For. Meteorol. 2021, 308–309, 108571. [Google Scholar] [CrossRef]
- Liu, Q.; Fu, Y.H.; Zhu, Z.; Liu, Y.; Liu, Z.; Huang, M.; Janssens, I.A.; Piao, S. Delayed Autumn Phenology in the Northern Hemisphere Is Related to Change in Both Climate and Spring Phenology. Glob. Chang. Biol. 2016, 22, 3702–3711. [Google Scholar] [CrossRef]
- Ren, S.; Yi, S.; Peichl, M.; Wang, X. Diverse Responses of Vegetation Phenology to Climate Change in Different Grasslands in Inner Mongolia during 2000–2016. Remote Sens. 2017, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Zu, J.; Zhang, Y.; Huang, K.; Liu, Y.; Chen, N.; Cong, N. Biological and Climate Factors Co-Regulated Spatial-Temporal Dynamics of Vegetation Autumn Phenology on the Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2018, 69, 198–205. [Google Scholar] [CrossRef]
- Li, P.; Peng, C.; Wang, M.; Luo, Y.; Li, M.; Zhang, K.; Zhang, D.; Zhu, Q. Dynamics of Vegetation Autumn Phenology and Its Response to Multiple Environmental Factors from 1982 to 2012 on Qinghai-Tibetan Plateau in China. Sci. Total Environ. 2018, 637–638, 855–864. [Google Scholar] [CrossRef]
- Li, P.; Zhu, Q.; Peng, C.; Zhang, J.; Wang, M.; Zhang, J.; Ding, J.; Zhou, X. Change in Autumn Vegetation Phenology and the Climate Controls From 1982 to 2012 on the Qinghai–Tibet Plateau. Front. Plant Sci. 2020, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, X.; Wang, H.; Ciais, P.; Peñuelas, J.; Myneni, R.B.; Desai, A.R.; Gough, C.M.; Gonsamo, A.; Black, A.T.; et al. Contrasting Responses of Autumn-Leaf Senescence to Daytime and Night-Time Warming. Nat. Clim. Chang. 2018, 8, 1092–1096. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wang, X.; Silander, J.A. Deciduous Forest Responses to Temperature, Precipitation, and Drought Imply Complex Climate Change Impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef] [Green Version]
- Ahlström, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The Dominant Role of Semi-Arid Ecosystems in the Trend and Variability of the Land CO2 Sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Fu, Y.H.; Zeng, Z.; Huang, M.; Li, X.; Piao, S. Temperature, Precipitation, and Insolation Effects on Autumn Vegetation Phenology in Temperate China. Glob. Chang. Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef]
- Wen, K.; Ren, G.; Li, J.; Zhang, A.; Ren, Y.; Sun, X.; Zhou, Y. Recent Surface Air Temperature Change over Mainland China Based on an Urbanization-Bias Adjusted Dataset. J. Clim. 2019, 32, 2691–2705. [Google Scholar] [CrossRef]
- Ren, S.; Peichl, M. Enhanced Spatiotemporal Heterogeneity and the Climatic and Biotic Controls of Autumn Phenology in Northern Grasslands. Sci. Total Environ. 2021, 788, 147806. [Google Scholar] [CrossRef]
- Park, T.; Chen, C.; Macias-Fauria, M.; Tømmervik, H.; Choi, S.; Winkler, A.; Bhatt, U.S.; Walker, D.A.; Piao, S.; Brovkin, V.; et al. Changes in Timing of Seasonal Peak Photosynthetic Activity in Northern Ecosystems. Glob. Chang. Biol. 2019, 25, 2382–2395. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; He, H.S.; Zhao, J.; Larsen, D.R.; Zhang, H.; Sunde, M.G.; Duan, S. Climate and Spring Phenology Effects on Autumn Phenology in the Greater Khingan Mountains, Northeastern China. Remote Sens. 2018, 10, 449. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Shi, J. Evaluation and Comparison of Growing Season Metrics in Arid and Semi-Arid Areas of Northern China under Climate Change. Ecol. Indic. 2021, 121, 107055. [Google Scholar] [CrossRef]
- Tao, Z.; Wang, H.; Liu, Y.; Xu, Y.; Dai, J. Phenological Response of Different Vegetation Types to Temperature and Precipitation Variations in Northern China during 1982–2012. Int. J. Remote Sens. 2017, 38, 3236–3252. [Google Scholar] [CrossRef]
- Zhang, R.; Qi, J.; Leng, S.; Wang, Q. Long-Term Vegetation Phenology Changes and Responses to Preseason Temperature and Precipitation in Northern China. Remote Sens. 2022, 14, 1396. [Google Scholar] [CrossRef]
- Kumari, N.; Srivastava, A.; Dumka, U.C. A Long-Term Spatiotemporal Analysis of Vegetation Greenness over the Himalayan Region Using Google Earth Engine. Climate 2021, 9, 109. [Google Scholar] [CrossRef]
- Wu, C.; Chen, J.M.; Black, T.A.; Price, D.T.; Kurz, W.A.; Desai, A.R.; Gonsamo, A.; Jassal, R.S.; Gough, C.M.; Bohrer, G.; et al. Interannual Variability of Net Ecosystem Productivity in Forests Is Explained by Carbon Flux Phenology in Autumn: Autumn Phenology and NEP. Glob. Ecol. Biogeogr. 2013, 22, 994–1006. [Google Scholar] [CrossRef]
- Zhang, J.; Tong, X.; Zhang, J.; Meng, P.; Li, J.; Liu, P. Dynamics of Phenology and Its Response to Climatic Variables in a Warm-Temperate Mixed Plantation. For. Ecol. Manag. 2021, 483, 118785. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Y.; Wang, D.; Chen, J.; Gu, X. Photoperiod Explains the Asynchronization Between Vegetation Carbon Phenology and Vegetation Greenness Phenology. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005636. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation Phenology on the Qinghai-Tibetan Plateau and Its Response to Climate Change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking Near-Surface and Satellite Remote Sensing Measurements of Deciduous Broadleaf Forest Phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, B.; Yang, Q.; Chen, G.; Yang, B.; Lu, L.; Shen, M.; Peng, Y. Responses of Net Primary Productivity to Phenological Dynamics in the Tibetan Plateau, China. Agric. For. Meteorol. 2017, 232, 235–246. [Google Scholar] [CrossRef]
- Guo, M.; Wu, C.; Peng, J.; Lu, L.; Li, S. Identifying Contributions of Climatic and Atmospheric Changes to Autumn Phenology over Mid-High Latitudes of Northern Hemisphere. Glob. Planet. Chang. 2021, 197, 103396. [Google Scholar] [CrossRef]
- White, J.H.R.; Walsh, J.E.; Thoman Jr, R.L. Using Bayesian Statistics to Detect Trends in Alaskan Precipitation. Int. J. Climatol. 2021, 41, 2045–2059. [Google Scholar] [CrossRef]
- Peng, J.; Wu, C.; Zhang, X.; Wang, X.; Gonsamo, A. Satellite Detection of Cumulative and Lagged Effects of Drought on Autumn Leaf Senescence over the Northern Hemisphere. Glob. Chang. Biol. 2019, 25, 2174–2188. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Zhao, L.; Lin, A.; Li, Q.; She, D.; Qu, S. How Do Climatic and Non-Climatic Factors Contribute to the Dynamics of Vegetation Autumn Phenology in the Yellow River Basin, China? Ecol. Indic. 2020, 112, 106112. [Google Scholar] [CrossRef]
- Yang, Z.; Shen, M.; Jia, S.; Guo, L.; Yang, W.; Wang, C.; Chen, X.; Chen, J. Asymmetric Responses of the End of Growing Season to Daily Maximum and Minimum Temperatures on the Tibetan Plateau. J. Geophys. Res. Atmos. 2017, 122, 13278–13287. [Google Scholar] [CrossRef]
- Jin, H.; Tong, S.; Chen, J.; Huang, X.; Shao, C.; Mandakh, U.; Chopping, M.; Du, L. Autumn Phenology and Its Covariation with Climate, Spring Phenology and Annual Peak Growth on the Mongolian Plateau. Agric. For. Meteorol. 2021, 298–299, 108312. [Google Scholar]
- Wu, L.; Ma, X.; Dou, X.; Zhu, J.; Zhao, C. Impacts of Climate Change on Vegetation Phenology and Net Primary Productivity in Arid Central Asia. Sci. Total Environ. 2021, 796, 149055. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Chen, S.; Jin, J.; Lu, X.; Jiang, H. Temporal and Spatial Dynamics of Phenology along the North–South Transect of Northeast Asia. Int. J. Remote Sens. 2019, 40, 7922–7940. [Google Scholar] [CrossRef]
- Zhu, W.; Tian, H.; Xu, X.; Pan, Y.; Chen, G.; Lin, W. Extension of the Growing Season Due to Delayed Autumn over Mid and High Latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- Cong, N.; Piao, S.; Chen, A.; Wang, X.; Lin, X.; Chen, S.; Han, S.; Zhou, G.; Zhang, X. Spring Vegetation Green-up Date in China Inferred from SPOT NDVI Data: A Multiple Model Analysis. Agric. For. Meteorol. 2012, 165, 104–113. [Google Scholar] [CrossRef]
- Pei, T.; Ji, Z.; Chen, Y.; Wu, H.; Hou, Q.; Qin, G.; Xie, B. The Sensitivity of Vegetation Phenology to Extreme Climate Indices in the Loess Plateau, China. Sustainability 2021, 13, 7623. [Google Scholar] [CrossRef]
- Wang, G.; Huang, Y.; Wei, Y.; Zhang, W.; Li, T.; Zhang, Q. Inner Mongolian Grassland Plant Phenological Changes and Their Climatic Drivers. Sci. Total Environ. 2019, 683, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xi, Z.; He, X.; Chen, S.; Rossi, S.; Smith, N.G.; Liu, J.; Chen, L. Contrasting Temporal Variations in Responses of Leaf Unfolding to Daytime and Nighttime Warming. Glob. Chang. Biol. 2021, 27, 5084–5093. [Google Scholar] [CrossRef]
- An, S.; Chen, X.; Zhang, X.; Lang, W.; Ren, S.; Xu, L. Precipitation and Minimum Temperature Are Primary Climatic Controls of Alpine Grassland Autumn Phenology on the Qinghai-Tibet Plateau. Remote Sens. 2020, 12, 431. [Google Scholar] [CrossRef] [Green Version]
- Seyednasrollah, B.; Young, A.M.; Li, X.; Milliman, T.; Ault, T.; Frolking, S.; Friedl, M.; Richardson, A.D. Sensitivity of Deciduous Forest Phenology to Environmental Drivers: Implications for Climate Change Impacts Across North America. Geophys. Res. Lett. 2020, 47, e2019GL086788. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Zhang, Z.; Zhang, H.; Guo, X.; Yu, S.; Du, W.; Huang, F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sens. 2016, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Gonsamo, A.; Chen, J.M.; Ooi, Y.W. Peak Season Plant Activity Shift towards Spring Is Reflected by Increasing Carbon Uptake by Extratropical Ecosystems. Glob. Chang. Biol. 2018, 24, 2117–2128. [Google Scholar] [CrossRef]
- Fu, Y.S.H.; Campioli, M.; Vitasse, Y.; De Boeck, H.J.; Van den Berge, J.; AbdElgawad, H.; Asard, H.; Piao, S.; Deckmyn, G.; Janssens, I.A. Variation in Leaf Flushing Date Influences Autumnal Senescence and next Year’s Flushing Date in Two Temperate Tree Species. Proc. Natl. Acad. Sci. USA 2014, 111, 7355–7360. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Wang, X.; Meng, F.; Liu, Q.; Li, X.; Zhang, Y.; Piao, S. Three-dimensional Change in Temperature Sensitivity of Northern Vegetation Phenology. Glob. Chang. Biol. 2020, 26, 5189–5201. [Google Scholar] [CrossRef]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Infuence of Spring and Autumn Phenological Transitions on Forest Ecosystem Productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 1555, 3227–3246. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Zhao, L.; Lin, A.; Wang, L.; Li, Q.; She, D.; Qu, S. Impacts of Preseason Drought on Vegetation Spring Phenology across the Northeast China Transect. Sci. Total Environ. 2020, 738, 140297. [Google Scholar] [CrossRef]
- Keenan, T.F.; Richardson, A.D. The Timing of Autumn Senescence Is Affected by the Timing of Spring Phenology: Implications for Predictive Models. Glob. Chang. Biol. 2015, 21, 2634–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Luedeling, E.; Xu, J. Winter and Spring Warming Result in Delayed Spring Phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | EOS (DOY) | SD (Days) | Type | EOS (DOY) | SD (Days) |
|---|---|---|---|---|---|
| Broadleaved deciduoud forest (DBF) | 294.69 ± 10.65 | 3.70 ± 5.58 | Meadow (MEA) | 291.81 ± 11.87 | 7.33 ± 7.53 |
| Needleleaved deciduoud forest (DNF) | 282.13 ± 6.06 | 4.06 ± 4.68 | Slope grassland (SG) | 298.53 ± 22.39 | 9.64 ± 13.40 |
| Broadleaved evergreen forest (EBF) | 313.15 ± 6.35 | 6.71 ± 3.48 | Plain grassland (PG) | 295.26 ± 13.77 | 9.49 ± 6.72 |
| Needleleaved evergreen forest (ENF) | 296.99 ± 12.43 | 5.74 ± 4.76 | Desert grassland (DG) | 297.42 ± 15.17 | 11.86 ± 10.55 |
| Alpine and sub-alpine meadow (AM) | 286.28 ± 12.39 | 7.37 ± 8.52 | Alpine and sub-alpine plain grass (APG) | 280.02 ± 12.60 | 9.87 ± 11.71 |
| Type | Percentage (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Tmin (P) | Tmin (N) | Tmax (P) | Tmax (N) | Pre (P) | Pre (N) | Rad (P) | Rad (N) | |
| DBF | 31.68 | 4.57 | 6.45 | 5.46 | 4.46 | 2.85 | 8.77 | 3.96 |
| DNF | 16.46 | 4.15 | 2.38 | 12.18 | 5.61 | 2.96 | 13.71 | 5.03 |
| EBF | 6.52 | 6.52 | 7.61 | 6.52 | 0 | 20.65 | 7.61 | 4.35 |
| ENF | 24.24 | 3.76 | 11.79 | 9.51 | 5.90 | 3.17 | 7.15 | 3.61 |
| AM | 30.46 | 7.29 | 8.19 | 4.91 | 6.97 | 2.96 | 9.07 | 5.38 |
| MEA | 23.15 | 6.10 | 11.32 | 7.12 | 6.46 | 2.74 | 9.43 | 2.99 |
| SG | 29.16 | 3.73 | 13.80 | 2.65 | 3.67 | 2.23 | 6.39 | 2.05 |
| PG | 33.80 | 6.79 | 13.13 | 4.21 | 4.23 | 2.73 | 6.30 | 1.85 |
| DG | 38.78 | 4.55 | 10.92 | 3.41 | 3.50 | 2.07 | 7.77 | 1.62 |
| APG | 29.52 | 11.14 | 7.63 | 1.95 | 5.53 | 4.60 | 9.42 | 4.67 |
| Study Area | Trend (Days/Year) | Period | Reference |
|---|---|---|---|
| Temperate China | 0.12 ± 0.01 | 1982–2011 | [25] |
| Tibetan Plateau | 0.07 | 1982–2011 | [13] |
| Mongolian Plateau | −0.06 | 1982–2013 | [45] |
| Central Asia | −0.69 | 2000–2019 | [46] |
| North-south transect of Northeast Asia | −0.09 | 1982–2014 | [47] |
| This study | 0.21 | 1982–2016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Wang, R.; Liu, B.; Qian, Z.; Wu, Y.; Li, C. Responses of Vegetation Autumn Phenology to Climatic Factors in Northern China. Sustainability 2022, 14, 8590. https://doi.org/10.3390/su14148590
Li Z, Wang R, Liu B, Qian Z, Wu Y, Li C. Responses of Vegetation Autumn Phenology to Climatic Factors in Northern China. Sustainability. 2022; 14(14):8590. https://doi.org/10.3390/su14148590
Chicago/Turabian StyleLi, Zhaozhe, Ranghui Wang, Bo Liu, Zhonghua Qian, Yongping Wu, and Cheng Li. 2022. "Responses of Vegetation Autumn Phenology to Climatic Factors in Northern China" Sustainability 14, no. 14: 8590. https://doi.org/10.3390/su14148590