Abstract
The opportunity to introduce Phacelia tanacetifolia is still being debated, even if the species is already employed, as it is often sown in rotational set-aside schemes or to support pollinators. Therefore, we need further evidence on its effects on the environment and its attractiveness towards pollinators. This work aims at confirming its appeal towards the honeybee in a mixed agricultural and urban environment in the north of Italy, during late spring and early autumn flowering. We assessed flowering characteristics and honeybee visitation rates by employing observational plots; we recorded resource preferences and behaviours by focal observations and video records; finally, we annotated other flower visitors. Each plant is characterised by one or more flowering stems on which not all flowers open at the same time: an increased number of flowers is, therefore, due to an increase in the number of available flowering stems. Honeybees were highly interested in lacy phacelia resources, especially in nectar collection: pollen collection may not be sufficiently rewarding for bees to dedicate time to it. The only correlation we found was between visitation rates and temperature. Other insects were recorded visiting the flowers: these were especially Hymenoptera; Coleoptera and Lepidoptera were also abundant, while relatively few individuals of Hemiptera and Diptera were recorded. Our results indicate that the mass-flowering of the species is certainly of interest to local pollinators and can be considered to counteract their decline when sown as a single species or in flower mixtures. However, further studies are needed to assess the competition of this plant with native species and its ability to self-regenerate when not purposely planted.
1. Introduction
There is an increasing appreciation by farmers of the value linked to the use of cover crops [1] which can provide various ecosystem services (such as sequestration of carbon, soil macroporosity, microbial diversity, abundance and diversity of useful insects). For example, rotational set-aside schemes which require the establishment of a cover crop each year have been largely adopted in the UK and are considered to be an option to plant floral resources for pollinators [2]. Providing extra resources to pollinators has been a main goal in recent years considering the alarming decline that is recorded, especially in numbers of managed and wild bees [3]. However, a discussion is in progress as to which plant species to consider, with various seed mixtures under evaluation [2,4]. In any discussion, particular interest is given to exotic species, often preferred for their rapid growth.
Phacelia tanacetifolia Benth, 1853 (common name: lacy phacelia) is a herbaceous plant belonging to the family Hydrophyllaceae, native to North America, commonly used in arable rotations [5]. Petanidou [6] summarised the history of the introduction and naturalisation of this species in Europe, focusing on its potential use in Mediterranean xeric areas and pointed out its attractiveness, especially to honeybees. The species is thought to have a considerable potential as a food plant for insect pollinators, attracting Hymenoptera thanks to the combined provision of nectar and pollen resources. Its attractiveness can be attributed to its abundant flowering, occurring in dense inflorescences, each with two to four branches, and flowers that open sequentially on one side, leading the inflorescence to take on a curled shape. Castle et al. [7] found the protein content of lacy phacelia pollen higher (25% in fresh bee bread) than that recorded in maize, or in maize combined with flowering strips. The authors also recorded a decrease in the sensitivity of bees to pesticides related to the good nutritional value of the pollen. The value of lacy phacelia can also refer to the mineral content of its monofloral pollen pellets collected from returning foragers which are especially high in phosphorus, potassium and calcium [8]. Therefore, its use is increasing, both as part of flower mixtures for pollinators and as a cover crop. In Italy, beekeepers are especially interested in this species and require more experimental evidence on its suitability as a food supplement for honeybees.
However, the species has been recognised as potentially invasive out of its home range: Randall [9] indicated it as a species “naturalised in Australia, once it had escaped from gardens or cultivation elsewhere in the world”. Smither-Kopperl [10] advised vigilance on the potential of the species to become weedy or invasive, possibly displacing native vegetation when not well managed. In Italy, contrasting information exists on its status. Cecchi and Selvi [11] reported the species as being seldom recorded in locations outside of fields or road margins where it is intentionally cultivated. The spontaneous populations do not seem to become permanently established, and the authors reported sporadic records, especially in the northern part of the country. However, the data sheet on the website, Dryades [12], indicates that lacy phacelia is present in most Italian regions, from north to south, as a casual alien species, and only in Friuli-Venezia Giulia and Lazio as already naturalised. Some evidence exists on the potential competition of P. tanacetifolia with native plant species due to its high attractiveness to bee pollinators, although it apparently has little effect on native plant reproduction [13,14].
Regarding pollinators’ interest and visits paid to lacy phacelia, some literature already exists. Petanidou [6] carried out some field observations in the experimental plots (xeric or irrigated) that she established: she recorded Apis mellifera Linnaeus, 1758 as the most abundant visitors (hives were actually placed close to sown fields), accompanied by hoverflies, beetles, sweat bees and butterflies. The abundance and diversity of pollinator species varied according to the periods and field site. She also found honeybee abundance varied according to the season, with a stronger tendency to visit for nectar. She also recorded that lacy phacelia pollen constituted just a small fraction of the pollen collected by honeybees. Similar results were obtained by Sprague and colleagues [15] in New Zealand as they also found less pollen- than nectar-foragers in the field and by Owayss and colleagues [16] who indicated lacy phacelia as a good source for honey production in the drought environments of Saudi Arabia.
Local environmental conditions can modify the relationships that P. tanacetifolia establishes with local pollinators. More studies are needed to better understand the relationship of P. tanacetifolia flowering characteristics with the interactions established with local pollinators. Authors recorded different durations of overall flowering period; longer in the UK [17] and shorter in Greece [6]. Furthermore, flower lifespan and visits can be affected by local conditions, undergoing variations due to the main climatic conditions such as temperature and insolation. On the one hand, more studies are needed to better understand the relationship that P. tanacetifolia establishes with local pollinators, since this could prompt its use to sustain declining populations of insect pollinators and merge it with the interests of stakeholders such as farmers and beekeepers. Conversely, whenever observations of the uncontrolled spread of P. tanacetifolia arise, its employment should be prevented, and informative campaigns should be planned. In this work we address the first issue, as a response to a pressing interest of stakeholders in our area (Lombardy, northern Italy). We carried out field observations to investigate the plant species characteristics when sown in the area of interest and the visitation rate by Apis mellifera. Beekeepers and farmers are the stakeholders’ groups, therefore we carried out data collection following their usual activity of sowing and beekeeping. Although this approach implies some methodological restrictions, it allows larger participation at the local level, that may facilitate awareness of results. We focus on different aspects of A. mellifera worker bees on flowers in relation to the foraged resource (number of flowers visited per time unit, resource—pollen or nectar—collected during the visit) and coupled our behavioural observations with data on the flowering characteristics (floral display as number of stems with flowers and flowers in anthesis). We also recorded other insect pollinators and give an indication on abundance of various insect groups on the exotic species P. tanacetifolia.
2. Materials and Methods
2.1. Field Sites
The study was carried out in 2018, 2019, 2020 and 2021 in five agricultural fields (Figure 1a) sown with Phacelia tanacetifolia. Information on the field locations is summarised in Table 1. Sowing was carried out by farmers with different aims: to allow experimental observations; as a food plant to sustain local bee populations or to obtain green fodder yield. The scorpioid inflorescences (detail in Figure 1b) of P. tanacetifolia may be the sole source of food in the field (monoculture) or one component of a flower mixture. Agricultural fields were scattered within an area of roughly 100 km2, in the north of Italy, characterised by mixed agricultural and urban environments. The climate, according to Köppen’s Geiger classification, is temperate oceanic without a dry season and with warm summers (Cfa) [18].

Figure 1.
(a) View of the agricultural field with flowering P. tanacetifolia and apiaries placed on the edge; (b) detail of the scorpioid inflorescence of P. tanacetifolia; (c) schematic grid of observational plot distribution; (d) honeybee collecting nectar and (e) honeybee collecting pollen.

Table 1.
Information on the agricultural fields. Localities were very close to each other, in Como (CO) and Monza-Brianza (MB) provinces (Italy).
2.2. Observational Plots
We employed observational plots placed in the agricultural fields. The plot positions were randomly selected during each day of observations but followed an established grid (Figure 1c). A scheme was designed to place the plots accordingly to two variables: distance from the hives and inter-distance between plots (minimum distance: 16 m). Plots were randomly positioned into the field according to the distance from the hives: 16 m (P1–P3 in Figure 1c) or 32 m (D1–D3 in Figure 1c). Minimal distances were decided on the basis of the agricultural field size. Knowing that honeybee foraging is most efficient at 3 km [19], we expected the distance of 16 m and 32 m from the hive would not influence the visitation rate of different plots in the field. However, we decided to take the opportunity to evaluate if any gradient in the visitation rates could be detected according to the limited field size, and organised plot distribution accordingly.
We placed apiaries with an average of 6 hives (small differences may depend on normal beekeeping activity) managed by beekeepers that constantly controlled the health of the colonies. The colonies were different depending on site and season, but all underwent constant management. Since apiaries of Apis mellifera were intentionally positioned at the field edges just prior to the blossoming period, the distance from the apiary was one of the variables recorded during each individual observation. Other variables were time of observation (solar hour) and temperature. Temperature data were acquired in real time from the webpages of “Centro Meteo Lombardo” [20] choosing those stations closer to the field site in a radius of 10 km [Carate Brianza (Monza and Brianza province) at 10 km from Site 2, Vertemate con Minoprio (Como province) at less than 2 km from Site 1, 3, 4, 5]. The temperature provided by the stations was measured at a 2 m height from the soil.
Observational plots were 25 × 25 cm and were physically identified by a wooden tool placed underneath the flower canopy. Observations lasted 5 min and were usually carried out during a two-hour period during the interval from 10:00 to 16:30. Before starting the observations, data on the flowering state were recorded: the number of floral branches with at least one flower in anthesis and the number of flowers in anthesis on individual floral branches. Out of these two types of data, averages were later calculated to estimate flower availability in the plot at each observation period. During the 5-min observation, we recorded all visiting individuals of A. mellifera. An individual was considered visiting when entering the plot and visiting at least one flower to collect nectar or pollen.
Regarding flowering characteristics of the species, we considered height of floral stems (from the ground to the tip of the highest inflorescence of a plant). The floral stem is the part of the plant that hosts the flowers; it may be divided into multiple branches. We also recorded the number of floral branches with at least one flower in anthesis, the mean number of flowers per floral branch and the predicted abundance of flowers in anthesis at the observational plot.
2.3. Foraging Behaviour
We were interested in verifying the resource collection and preference of visiting Apis mellifera bees. Bees could visit the flowers to collect nectar and pollen. When collecting nectar (Figure 1d), they land on the flower and seek nectar inside the corolla, while pollen collection is a more elusive behaviour carried out quickly while hovering on top of the anthers (Figure 1e). The latter needs to be better understood.
To investigate visitation rates, we normalised data to the number of bees/flowers observed in the 5-min periods and then by verifying bee abundance according to the following variables: distance from apiaries, temperature and time of the day. To investigate daily activity, we assigned records to three time-classes (time class I: 9.00–12.30; time class II: 12.31–16.00 and time class III: 16.01–19.30).
To investigate resource preferences, we first monitored, whenever possible, the resource collected by bees entering the observational plots. These data could not be collected for each visit; we were able to define the resource in 469 observational sets. However, due to the elusive pollen collection behaviour, we believe pure pollen collection was underestimated during these observations: in fact, we recorded a single visit solely for pollen out of all observation sets. Therefore, we performed targeted observations, carrying out random focal observations in the field. Focal observations were based on an individual bee during its foraging bout, following it by sight for as long as possible up to a maximum of 1 min. During these observations, we recorded the number of visited flowers, the collected resource and the total monitoring duration. Data refer to 225 individual bees during their foraging bouts in the lacy phacelia fields. Finally, we video-recorded (Lumix DMC-ZX3, Panasonic Corporation, Osaka, Japan) foraging bees. From video analyses performed at a later stage, we could describe, in detail, the most elusive behaviour, the pollen collection.
2.4. Other Flower Visitors
Although our focus was on A. mellifera, we also recorded other insects during the observations on plots. Identification was carried out directly in the field, when possible. Otherwise, unknown insects were photographed or captured. Identification was later checked, and if the species level was not reached, we kept the information at genus or family level. Special attention was paid to Apoidea, which were collected in the field using plastic tubes (diameter of 3.5 cm, height of 12.3 cm) and kept in a portable freezer (5–15 °C). Later, morphological identification was carried out under a stereoscope until identification was made in the laboratory by consulting the dichotomous keys of Falk [21], Cappellari [22] and specialised sites [23,24].
2.5. Statistical Analysis
The data were analysed mostly using generalised linear models in the statistical software R (version 4.2.0, R Core Team, Indianapolis, IN, USA, 2020) with RStudio (1.3.1093; RStudio Team, Indianapolis, IN, USA, 2020) and SPSS V 26.0 (IBM, Armonk, NY, USA: IBM Corp.). For the analysis of integer response variables (e.g., bees/flower) we used log-linear models, assuming quasi-Poisson errors. This approach assumed error distributions (from within the exponential family) that are likely to match the natural patterns in different types of data and then allows further adjustment via empirically estimated scaling parameters, retaining a Type I error rate of approximately 5% without the need for prior transformation. Normally distributed data were described by the mean and standard error, and factors were compared with a univariate analysis of variance (ANOVA) through the general linear model procedure. Results from factors showing significant overall changes were subjected to post hoc Tukey’s tests with a Type I error rate < 0.05. The correlation analysis between the mean number of flowers and the mean number of honeybees was estimated with the Pearson’s correlation coefficient. All hypothesis testing was carried out using backwards elimination of explanatory variables from initial models and by aggregation of factor levels to find the minimum adequate statistical models. For pollinator monitoring, a pivot table with the summary of observations was imported into the statistical software R (version 4.2.0), using the additional packages Readxl [25] and Bipartite [26].
3. Results
Since we followed stakeholders’ sowing activity, our data refer mainly to the late-spring flowering period, except for one set of observations carried out during an early-autumn flowering. We also had the chance to record lacy phacelia while flowering as monoculture or as part of a flower mixture.
3.1. P. tanacetifolia Flowering Characteristics at Observational Plots
The height of floral stems varied slightly during each late-spring flowering period, roughly from 60 to 100 cm. We recorded: in late-spring flowering, an average height (±SE) of 92.63 ± 2.30 cm; in early-autumn flowering, an average height (±SE) of 72.73 ± 0.55 cm. We detected an exception in plants sown in the flower mixture, observed in 2021: in this case, floral stems were on average taller than those planted as a pure cover crop (average height (±SE) in the mixture 116.92 ± 2.66 cm vs. 77.30 ± 2.33 cm in the monoculture).
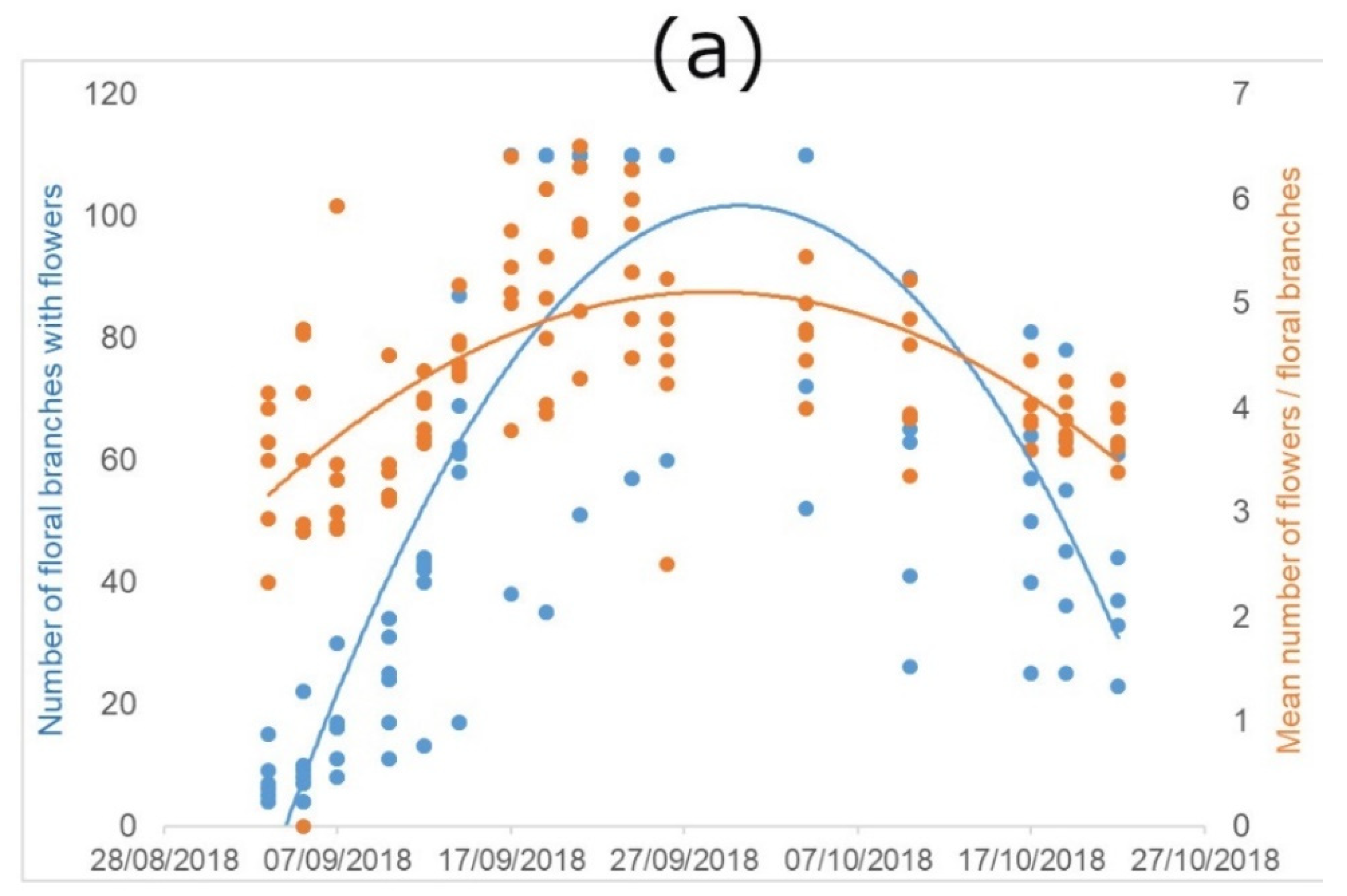
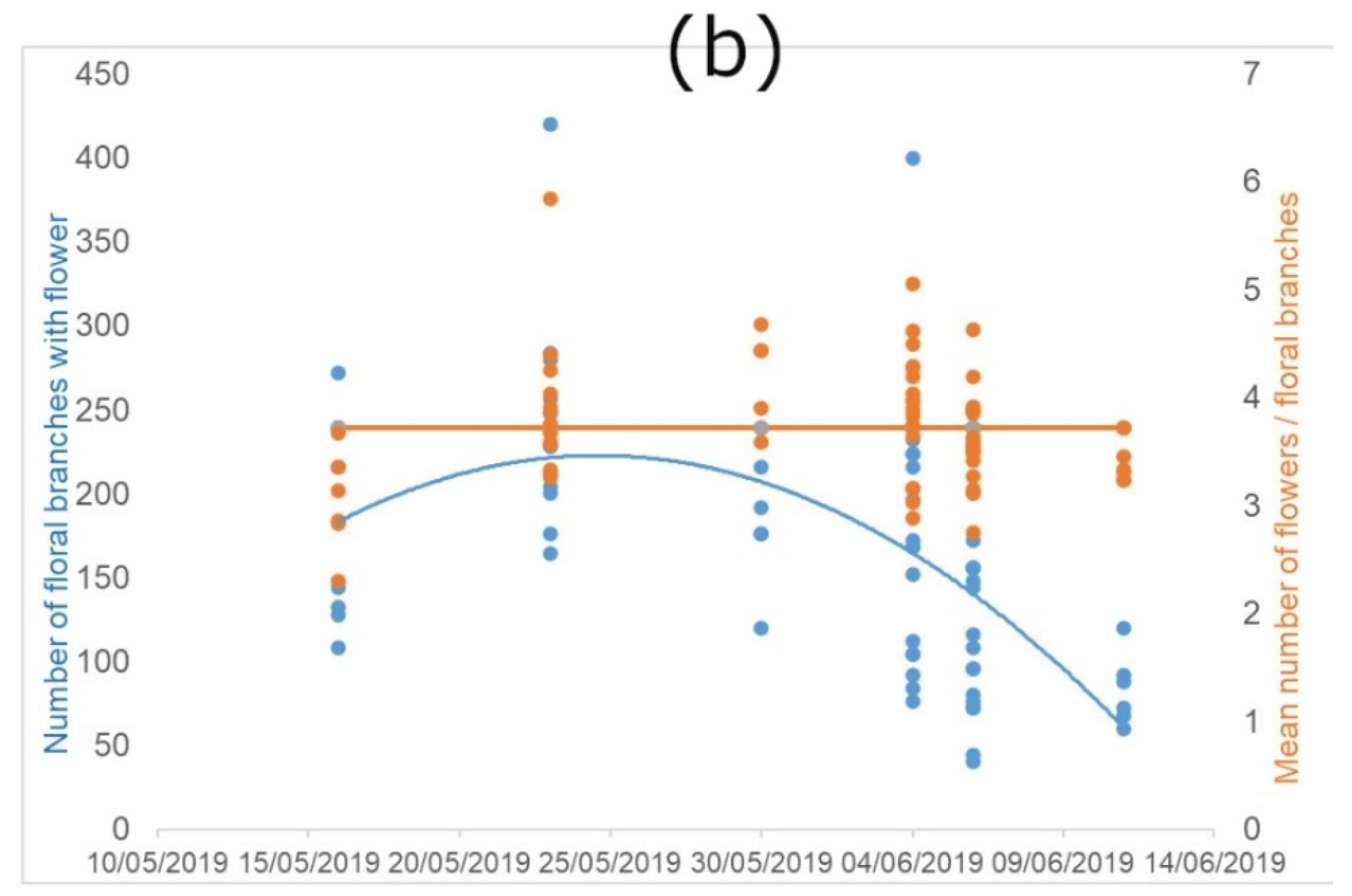
For the estimation of the floral display, we combined the information on the average number of flower stems with at least one flower in anthesis, and the average number of flowers per stem. We treated data separately for the two periods. However, we noticed very similar trends (Figure 2).
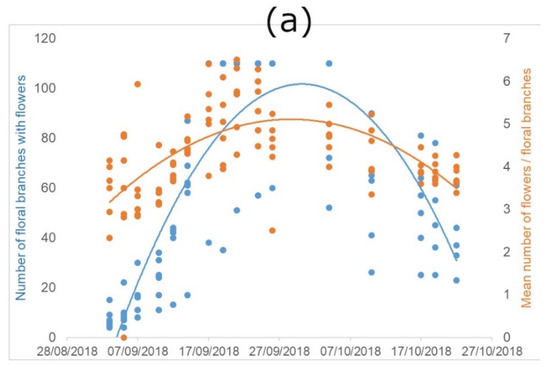
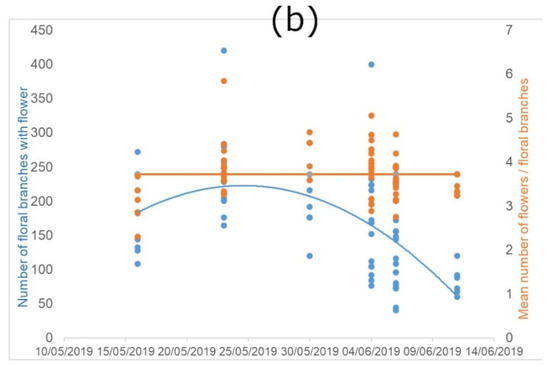
Figure 2.
Flowering characteristics of P. tanacetifolia. “Flowers” indicate flowers in anthesis. (a) represents the trend of the early-autumn flowering and (b) the trend of the late-spring flowering, considered as development of new floral branches with flowers, and number of flowers per branch along the flowering period (number of floral branches with flowers = blue dots and lines; mean number of flowers per floral branches = orange dots and lines).
The number of flowering stems shows a clear bell-shape, indicating that there is a peak in the flowering, with variability depending on the season. The regression lines of the early-autumn flowering (year 2018; Figure 2a) showed significant differences between time and branches (R2 = 0.38; F(2, 308) = 349.2; p < 0.0001) and also between time and flower in anthesis (R2 = 0.69; F(2, 308) = 96.08; p < 0.0001). We found the same significant differences in the regression line of late-spring flowering (year 2019; Figure 2b) between time and branches: (R2 = 0.2 F(1, 63) = 15.34; p < 0.001), but no significance between time and flower in anthesis.
As expected, we found a positive correlation (Pearson’s r = 0.612; p < 0.001) between the mean number of flowers and the mean number of bees.
3.2. Apis Mellifera Foraging Behaviour
From the observational plots, we normalised our records by number of bees/flowers observed in the 5-min periods. Our variables were distance from apiaries, time of day (by time classes), temperature at foraging and foraging along the flowering season.
Regarding the visitation rates and the distance from apiaries, by opposing data collected at sites P1–P3 in Figure 1c and D1–D3, we found no significant difference (F(1, 476) = 0.779; p = 0.38). Time of day and temperatures are related, time class I had the significantly (F(2, 470) = 15.75, p < 0.001) lowest mean temperature (22.109 ± 0.42 °C). The average temperature trends were very similar notwithstanding the different years and seasons (only the year 2021 had significantly higher mean temperature of 26.7 ± 0.42 °C (F(3, 470) = 19.85, p < 0.001) (Figure 3) without significant interaction between years and time class (F(4, 470) = 0.08, p = 0.9)). However, the two variables may differently influence bee activity.
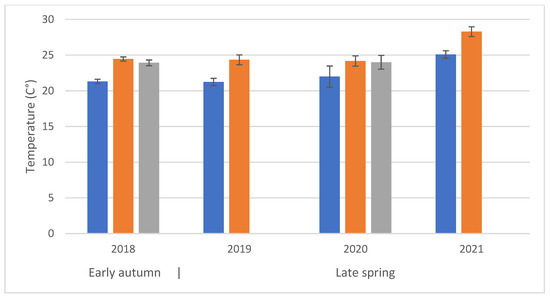
Figure 3.
Average temperature during observations at different times of day (time classes: Blue = time class I; Orange = time class II; Grey = time class III). The first set of observations (2018) refers to an early-autumn flowering; the following three to late-spring flowering periods.
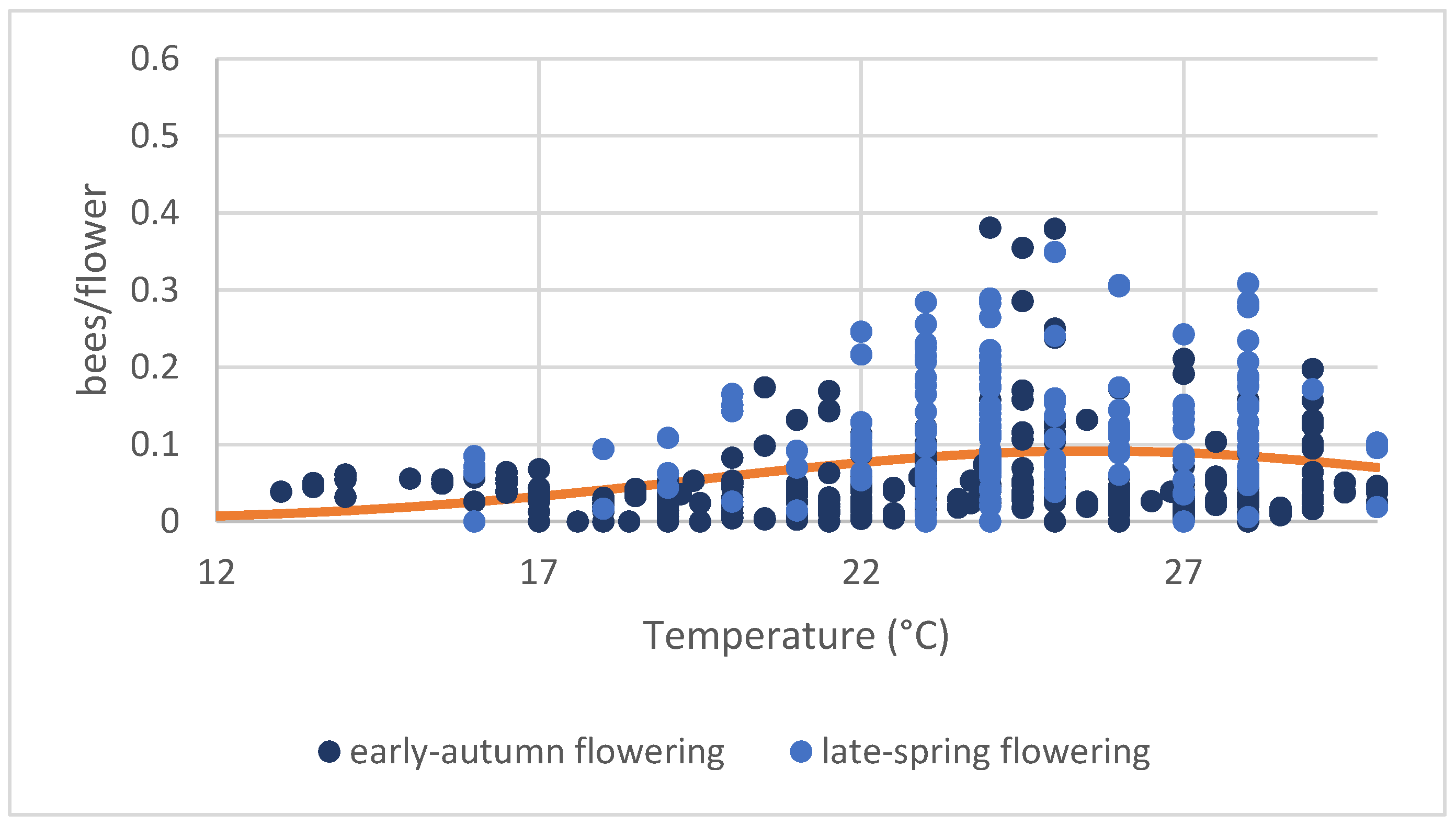
When considering daily activity by comparing the abundance of records among time classes, no significant differences in the number of bees/flowers emerged (F(2, 475) = 2.66; p = 0.071). However, the opposite was true for temperatures (Figure 4). The average number of bees showed a significantly curvilinear response to increasing temperature (log-linear regression, including a quadratic term: temperature: F(1, 476) = 17.886, p < 0.001, %Dev = 0.01; temperature2: F(1, 475) = 13.392, p < 0.001, %Dev = 0.01).
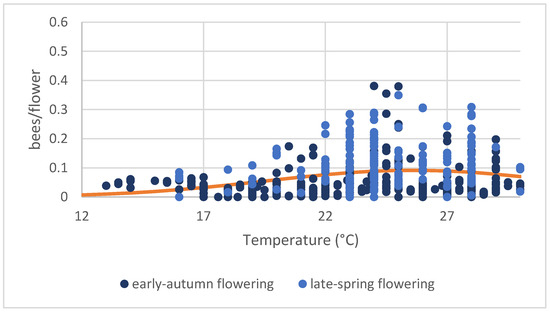
Figure 4.
Bee abundance per flower (in five minutes) related to temperature considering the two different sowings: early-autumn (blue dots) and late-spring (light-blue dots). Two points were excluded from the graph because they were considered outliers (0.71 bees/flowers at 25 °C and 1.1 bee/flowers at 27 °C). The regression line between the two variables is shown in orange (log-linear regression including a quadratic term: temperature: F(1, 476) = 17.886, p < 0.001, %Dev = 0.01; temperature2: F(1, 475) = 13.392, p < 0.001, %Dev = 0.01).
Regarding resource preferences, data from the focal observations indicated that the bees manage to visit a greater number of flowers if the resource collected is only pollen (Video S1). Bee foragers collecting only nectar visited, on average, 11.31 ± 0.6 SE flowers; foragers looking for both resources visited, on average, 11.03 ± 0.75 SE flowers; foragers collecting only pollen visited, on average, 20.6 ± 1.33 SE flowers. The difference in the average number of visited flower is significant (F(2, 87) = 22.03; p < 0.001). However, only 11 of the individual bees observed foraged exclusively for pollen, contrary to the 55% that foraged exclusively for nectar.
Video-recordings highlighted the complexity of manipulating anthers to get lacy phacelia pollen. Stamens protrude out of the corolla and the flower does not offer a stable platform to land, stopping flight and reaching the anthers at once. To collect the pollen, bees needed to maintain an unstable position, grabbing one or two fragile filaments, and using the mouth and forelegs to break the anthers and get the pollen (See Video S1). This action certainly influences the amount of pollen finally accumulated. In Figure 1e, we can appreciate how the unstable position is maintained by the middle legs and the buzzing wings, while forelegs are holding a single filament, the anther possibly being held in the mouth.
3.3. Other Visitors
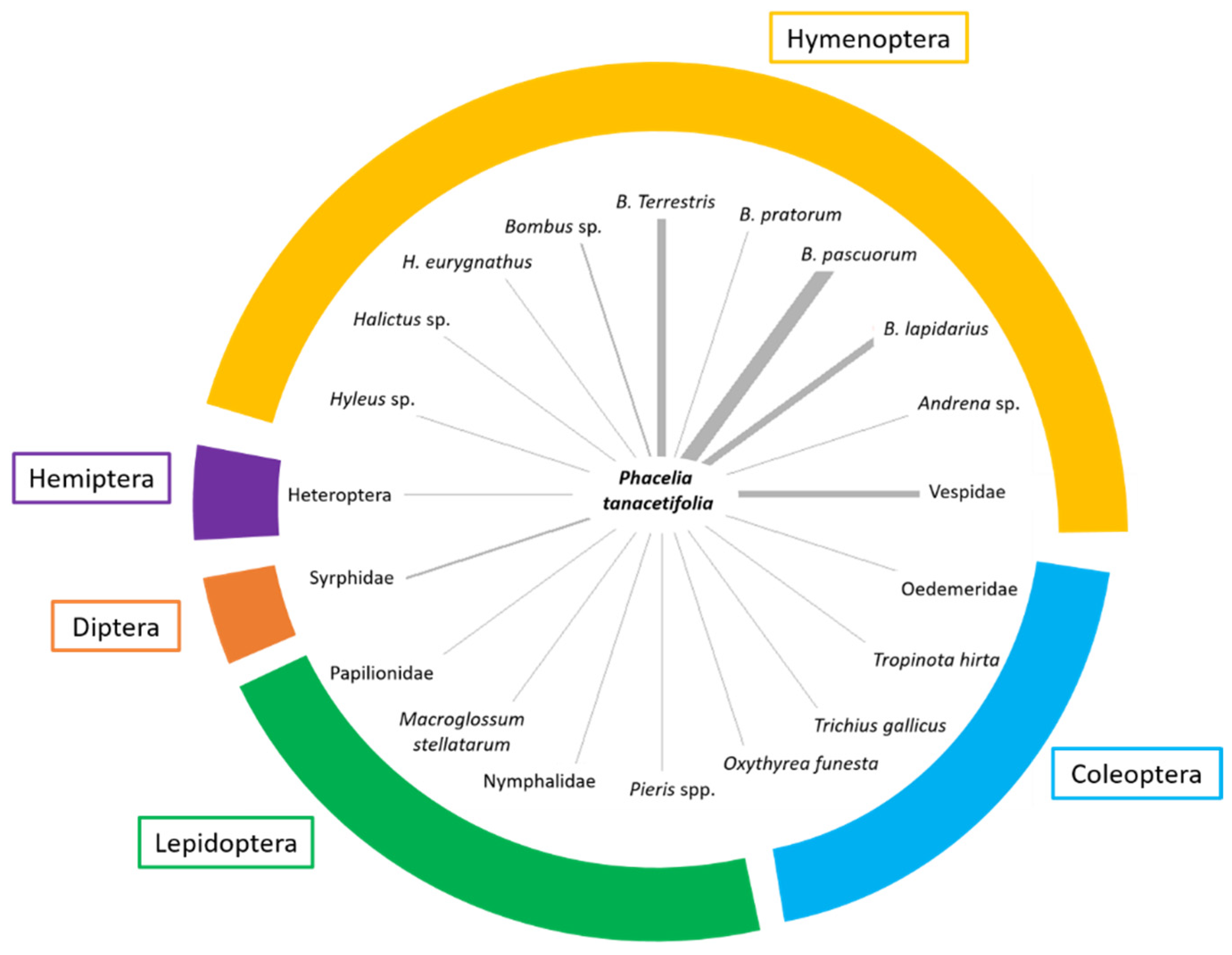
Excluding A. mellifera, we observed many other insects on the flowers (Figure 5). They were recorded when they entered the observational plots. The largest group is represented by the order Hymenoptera, with a special contribution of Bombus species (in order of abundance: Bombus pascuorum (Scopoli, 1763), B. terrestris (Linnaeus, 1758), B. lapidarius (Linnaeus, 1758) and B. pratorum (Linnaeus, 1761), that were found in association with P. tanacetifolia especially in the early-autumn before mating and overwintering. Among wild bees, Halictus spp. and Hylaeus sp. were also detected. We also recorded a similar abundance of individuals belonging to the orders Lepidoptera, mainly Papilionidae [Iphiclides podalirius (Linnaeus, 1758) and Papilio machaon Linnaeus, 1758]; Pieridae [Pieris spp.]; Nymphalidae [Issoria lathonia (Linnaeus, 1758); Hyponephele lycaon (Küns, 1774)] and Sphingidae (Macroglossum stellatarum (Linnaeus, 1758)]. Both Hymenoptera and Lepidoptera are able to enter, with their ligula, the corolla of P. tanacetifolia flowers to have access to nectar similarly to what was observed with A. mellifera (see Video S1). On the contrary, few hoverflies (Diptera, Syrphidae) were observed on flowers as their proboscis is less adapted to enter the narrow flowers to have access to nectar, and pollen is localised in anthers which are not easy to reach. Among Coleoptera, we detected four species, and three of them, Oxythyrea funesta (Poda, 1761), Tropinota (Epicometis) hirta (Poda, 1761) and Trichius gallicus Dejean, 1821 are Scarabeidae and one is the Odemeridae Oedemera sp. Hemiptera were rarer, and it was possible to detect a few specimens of Pentatomidae [Nezara viridula (Linnaeus, 1758), Dolycoris baccarum Linnaeus, Carpocoris purpuripennis (De Geer, 1773)] and Lygaidae (Lygus rugulipennis Poppius, 1911). Among Diptera, we detected just a few specimens of Syrphidae (Syrphus spp.).
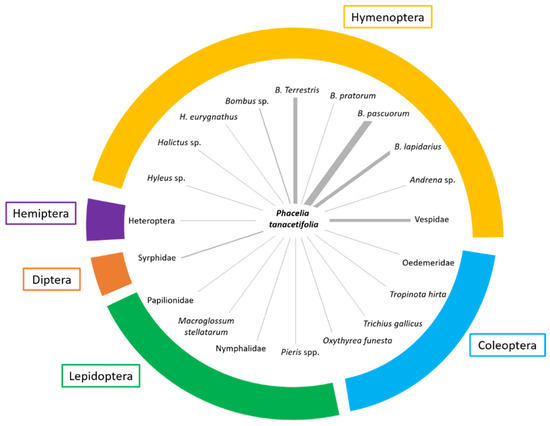
Figure 5.
Distribution and abundance of flower visitors other than A. mellifera in the different orders. The thickness of the rays indicates the abundance of the taxa in the monitoring periods. Data of different periods and localities were pulled together.
4. Discussion
In this work we provide a set of data to better understand the interest that P. tanacetifolia raises in Apis mellifera and other pollinators. This interest is currently positively received by beekeepers who foster the sowing of the flower crop to sustain honey production [15,16,27]. Positive feedback also came from the community at large, interested in increasing resources for wild pollinators by planting floral strips: lacy phacelia seeds are therefore part of mixtures used for this purpose, with an added value of the pleasure given by the view of their intense coloured flowers. We performed plot and focal observations on honeybees visiting lacy phacelia and recorded other flower visitors and plant growth.
As other authors pointed out [6,15,17], P. tanacetifolia is a species that honeybees frequently visit when it is present in the environment, especially for nectar collection. Pollen is, in fact, less frequently collected and our video records and behavioural observations highlighted that pollen collection is not particularly easy on this species: the stamen filaments, protruding high on top of the corolla, do not facilitate pollen gathering. Even data on flowers visited exclusively for pollen confirm the above: possibly due to the unstable position, an individual bee cannot stay too long in a hovering position and, therefore, visits a larger number of flowers to complete a pollen-collecting bout. Petanidou [6] and Sprague et al. [15] recorded limited lacy phacelia pollen in the pollen traps that they placed: another indication that the pollen of P. tanacetifolia is not to be considered a good resource, while nectar is. Nectar is actively pursued as the primary resource by more than half of the honeybee individuals we recorded. This result may support the interest of beekeepers in sowing this species or looking for its presence as a monoculture.
However, some preconditions to sowing should be ascertained: the sugar content of nectar may vary among cultivars [28] and temperature may strongly influence nectar viscosity and bees’ ability to suck. Moreover, the nectar standing crop may be highly variable, as reported by Williams [29]. In this study, Williams [29] analysed flower age and stage (the second possibly linked to flower age, or visitation rates and environmental microclimate or a combination of these factors). These are two variables often strongly linked with nectar content. Only flower stage related to nectar content, and this information seems to be considered by bees when deciding which flower to visit [30]. However, a large percentage of the secretion measured by Williams [29] remained unexplained. Further analyses are certainly needed to explain nectar (and honey) productivity. Furthermore, the growing conditions of the plants may be differently influenced by the season or the presence of other plant species in a seed mixture. In our study, only small differences were recorded in the height of floral stems independently from the season and the place where it was cultivated when it was in a monoculture. On the contrary, the plants grown in a mixed seeded plot were taller for increasing interspecific competition with other plants coexisting with lacy phacelia. The number of flowers per stem was constant, but the total number of flowers increased when the stem number was greater. This is also an important since flower opening is controlled by the plant and may be reduced, prolonging the overall flowering period. This may be another point of interest for beekeepers interested in increasing honey production by prolonging the flowering season.
We could not compare honeybee visits with those paid by other pollinator species since, in our experimental design, apiaries were placed close to the crop. However, we did record other flower visitors. The most frequent belonged to the genus Bombus, and foragers of various species of this genus were already recorded as highly interested in collecting lacy phacelia nectar [17,31]. Other studies highlighted how various bee species are attracted by P. tanacetifolia when planted in gardens [14]. The association of different Lepidoptera species also emphasises the role of this plant in sustaining these pollinators, which have undergone a serious decline in most European countries following rapid changes in land use in recent decades [32]. Among Coleoptera, we found Odemera sp. that, similarly to bees, are known to feed on both the pollen and nectar from many plant species [33] but also some Scarabeidae that are known to damage flowers as they eat all its parts [34] and, in the case of strong gradation, can interfere with bee activity.
5. Conclusions
Our main findings support the following:
- P. tanacetifolia is a good source of nectar for A. mellifera, while its pollen is not as attractive. More than half of our records on honeybee foragers refer to nectar collection, and our video records highlighted the difficulties in pollen collection.
- P. tanacetifolia flowering characteristics show relatively small variability among individuals. However, overall floral display may vary greatly. We found a constant number of flowers per stem, but an increased total number of flowers linked to the stem number.
- As for sustaining beekeeping activity and honey production, sowing P. tanacetifolia even as a monoculture may result in good practice. The intrinsic characteristics of the plant need to be investigated further to find the best balance between sowing time, plant variety and honey production.
In spite of the positive conclusions so far underlined, we would like to record the need to remain alert on the use of exotic species without a sound assessment of the interactions between local plants and their mutualistic partners. Totland and colleagues [13] investigated how P. tanacetifolia was interfering with the reproduction of a local species in boreal forest ecosystems. They found an overall increased number of floral visitors that turned to the exotic species for most of their visits. In their study, unfortunately and contrasting with the negative impact on pollinators’ visits, the presence of P. tanacetifolia did not affect seed production of the native floral species. We need to carefully assess the value of exotic species, balancing risks and benefits.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/article/10.3390/su141610186/s1, Video S1: pollen and nectar collection by honeybees.
Author Contributions
Conceptualization, D.L. and M.G.; methodology, D.L.; formal analysis, S.M., D.L. and M.G.; investigation, M.Z. and S.M.; data curation, S.M., M.G. and D.L.; writing—original draft preparation, M.G.; writing—review and editing, M.G., D.L. and S.M.; supervision, D.L.; funding acquisition, D.L. All authors have read and agreed to the published version of the manuscript.
Funding
The research was partially supported by APIGIS—Project ID: 2019-4675 founded by Cariplo Foundation.
Institutional Review Board Statement
Not applicable for studies involving insects.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to acknowledge all beekeepers that provided access to their apiaries and fields (Giancarlo Costenaro and Riccardo Galizioli) and Fondazione Minoprio that hosted the research in 2018.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Storr, T.; Simmons, R.W.; Hannam, J.A. A UK survey of the use and management of cover crops. Ann. Appl. Biol. 2019, 174, 179–189. [Google Scholar] [CrossRef]
- Carreck, N.L.; Williams, I.H. Food for insect pollinators on farmland: Insect visits to flowers of annual seed mixtures. J. Insect Conserv. 2002, 6, 13–23. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Carvell, C.; Westrich, P.; Meek, W.R.; Pywell, R.F.; Nowakowski, M. Assessing the value of annual and perennial forage mixtures for bumblebees by direct observation and pollen analysis. Apidologie 2006, 37, 326–340. [Google Scholar] [CrossRef]
- Bacq-Labreuil, A.; Crawford, J.; Mooney, S.J.; Neal, A.L.; Ritz, K. Phacelia (Phacelia tanacetifolia Benth.) affects soil structure differently depending on soil texture. Plant Soil 2019, 441, 543–554. [Google Scholar] [CrossRef]
- Petanidou, T. Introducing plants for bee-keeping at any cost? Assessment of Phacelia tanacetifolia as nectar source plant under xeric Mediterranean conditions. Plant Syst. Evol. 2003, 238, 155–168. [Google Scholar] [CrossRef]
- Castle, D.; Alkassab, A.T.; Bischoff, G.; Steffan-Dewenter, I.; Pistorius, J. High nutritional status promotes vitality of honey bees and mitigates negative effects of pesticides. Sci. Total Environ. 2022, 806, 151280. [Google Scholar] [CrossRef]
- Liolios, V.; Tananaki, C.; Papaioannou, A.; Kanelis, D.; Rodopoulou, M.A.; Argena, N. Mineral content in monofloral bee pollen: Investigation of the effect of the botanical and geographical origin. J. Food Meas. Charact. 2019, 13, 1674–1682. [Google Scholar] [CrossRef]
- Randall, R. Garden thugs, a national list of invasive and potentially invasive garden plants. Plant Prot. Q 2001, 16, 138–171. [Google Scholar]
- Smither-Kopperl, M. Plant Guide for Lacy Phacelia (Phacelia tanacetifolia); USDA-Natural Resources Conservation Service, Lockeford Plant Materials Center: Lockeford, CA, USA, 2018; p. 95237.
- Cecchi, L.; Selvi, F. Flora Critica d’Italia: Hydrophyllaceae; Fondazione per la Flora Italiana, Ed.; Società Botanica Italiana and Fondazione Internazionale pro Herbario Mediterraneo: Palermo, Italy, 2015. [Google Scholar]
- DRYADES. Phacelia tanacetifolia Benth. 2022. Available online: http://dryades.units.it/FVG/index.php?procedure=taxon_page&id=4243&num=4835 (accessed on 9 May 2022).
- Totland, Ø.; Nielsen, A.; Bjerknes, A.L.; Ohlson, M. Effects of an exotic plant and habitat disturbance on pollinator visitation and reproduction in a boreal forest herb. Am. J. Bot. 2006, 93, 868–873. [Google Scholar] [CrossRef]
- Frankie, G.; Feng, I.; Thorp, R.; Pawelek, J.; Chase, M.H.; Jadallah, C.C.; Rizzardi, M. Native and non-native plants attract diverse bees to urban gardens in California. J. Pollinat. Ecol. 2019, 25, 16–23. [Google Scholar] [CrossRef]
- Sprague, R.; Boyer, S.; Stevenson, G.M.; Wratten, S.D. Assessing pollinators’ use of floral resource subsidies in agri-environment schemes: An illustration using Phacelia tanacetifolia and honeybees. PeerJ 2016, 4, e2677. [Google Scholar] [CrossRef] [PubMed]
- Owayss, A.A.; Shebl, M.A.; Iqbal, J.; Awad, A.M.; Raweh, H.S.; Alqarni, A.S. Phacelia tanacetifolia can enhance conservation of honey bees and wild bees in the drastic hot-arid subtropical Central Arabia. J. Apic. Res. 2020, 59, 569–582. [Google Scholar] [CrossRef]
- Williams, I.H.; Christian, D.G. Observations on Phacelia tanacetifolia Bentham (Hydrophyllaceae) as a food plant for honey bees and bumble bees. J. Apic. Res. 1991, 30, 3–12. [Google Scholar] [CrossRef]
- Fratianni, S.; Acquaotta, F. The climate of Italy. In Landscapes and Landforms of Italy; Soldati, M., Marchetti, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 29–38. [Google Scholar]
- Visscher, P.K.; Seeley, T.D. Foraging strategy of honeybee colonies in a temperate deciduous forest. Ecology 1982, 63, 1790–1801. [Google Scholar] [CrossRef]
- Dati Rete CML, Stazioni Online 2021. Available online: www.centrometeolombardo.com (accessed on 7 July 2022).
- Falk, S. Field Guide to the Bees of Great Britain and Ireland; Bloomsbury Publishing: London, UK, 2019. [Google Scholar]
- Cappellari, A.; Mei, M.; Lopresti, M.; Cerretti, P. BumbleKey: An interactive key for the identification of bumblebees of Italy and Corsica (Hymenoptera, Apidae). ZooKeys 2018, 12, 127–138. [Google Scholar] [CrossRef]
- Beewatching. Scopri le Api. 2021. Available online: https://www.beewatching.it/scopri-le-api/ (accessed on 10 May 2022).
- BWARS. Bees, Wasp and Ant Recording Society. 2020. Available online: https://www.bwars.com/ (accessed on 10 October 2020).
- Wickham, H.; Bryan, J. readxl: Read Excel Files. 2019 R Package Version 1.3.1. Available online: https://CRAN.R-project.org/package=readxl (accessed on 20 May 2022).
- Dormann, C.F.; Gruber, B.; Fruend, J. Introducing the bipartite Package: Analysing Ecological Networks. R News 2008, 8, 8–11. [Google Scholar]
- Howes, F.N. Plants and Beekeeping-an Account of Those Plants, Wild and Cultivated, of Value to the Hive Bee, and for Honey Production in the British Isles; Read Books Ltd.: Plano, TX, USA, 2013. [Google Scholar]
- Nikolova, T.; Petrova, V. The climate factors impact on the nectariferous qualities of Phacelia tanacetifolia Benth. Sci. Pap. Ser. D Anim. Sci. 2019, 62, 267–271. [Google Scholar]
- Williams, C.S. Nectar secretion rates, standing crops and flower choice by bees on Phacelia tanacetifolia. J. Apic. Res. 1997, 36, 23–32. [Google Scholar] [CrossRef]
- Giovanetti, M. Foraging choices balanced between resource abundance and handling concerns: How the honeybee, Apis mellifera, select the flowers of Robinia pseudoacacia. Bull. Entomol. Res. 2019, 109, 316–324. [Google Scholar] [CrossRef]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Bumblebees experience landscapes at different spatial scales: Possible implications for coexistence. Oecologia 2006, 149, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Warren, M.S.; Bourn, N.A. Ten challenges for 2010 and beyond to conserve Lepidoptera in Europe. J. Insect Conserv. 2011, 15, 321–326. [Google Scholar] [CrossRef]
- Sivilov, O.; Atanassova, J.; Zlatkov, B. Food plant spectrum of Oedemeridae species (Insecta, Coleoptera) based on pollen analysis (a preliminary study). Comptes Rendus De L’Académie Bulg. Des Sci. 2011, 64, 224–230. [Google Scholar]
- Vuts, J.; Baric, B.; Razov, J.; Toshova, T.B.; Subchev, M.; Sredkov, I.; Tóth, M. Performance and selectivity of floral attractant-baited traps targeted for cetoniin scarabs (Coleoptera: Scarabaeidae) in Central and Southern Europe. Crop Prot. 2010, 29, 1177–1183. [Google Scholar] [CrossRef]

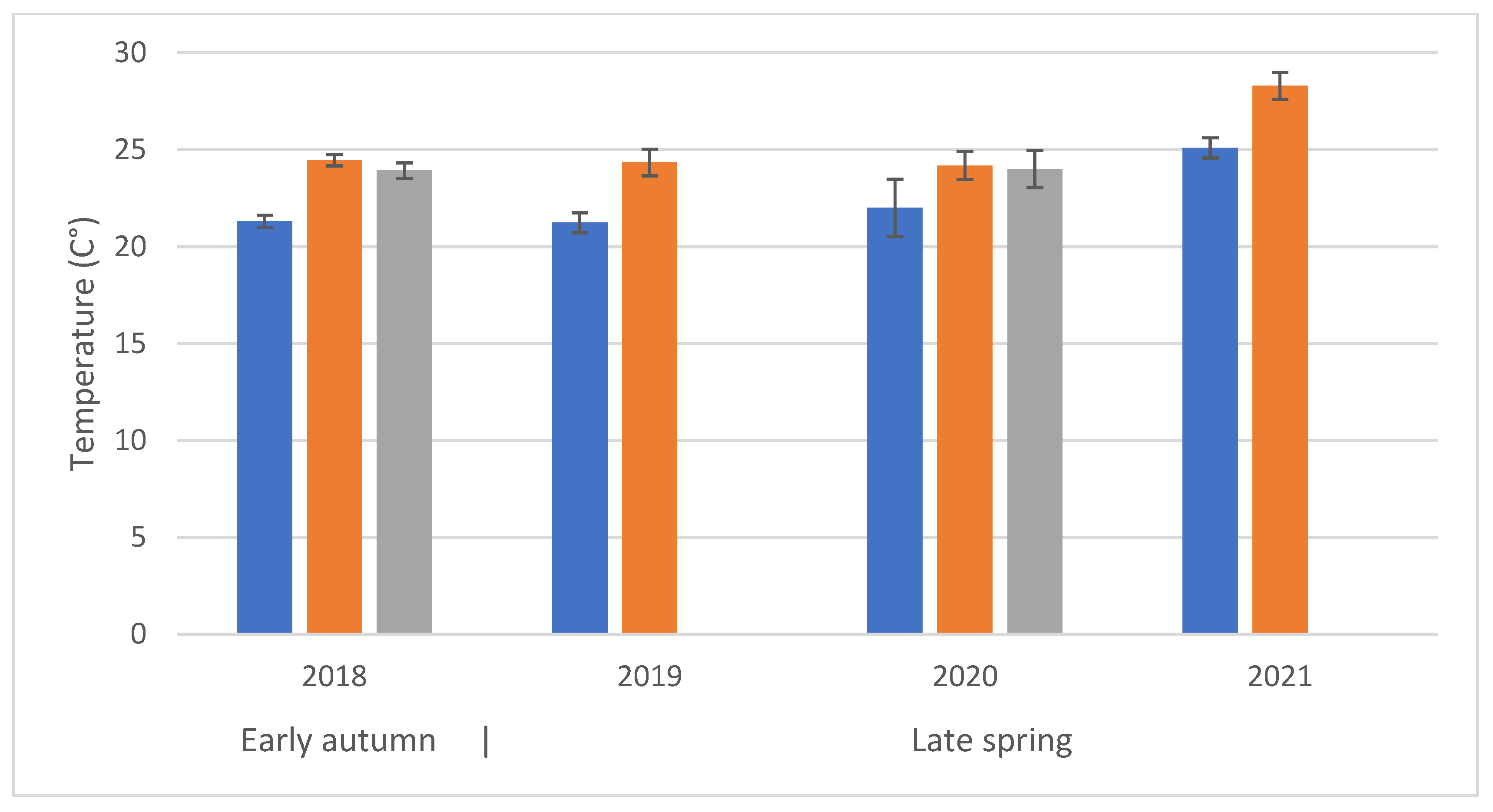
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).