
Effects of Breeding Forest Musk Deer on Soil Bacterial Community Structure

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey of the Sampling Area
2.2. Samples Collection
2.3. Component Analysis of Soil Samples
2.4. DNA Extraction and High-Throughput Sequencing
2.5. Statistical and Bioinformatics Analyses
3. Results
3.1. Soil Characteristics Differences between Captive and Non-Captive Forest Musk Deer Soils
3.2. Sequence Richness and Diversity Analysis
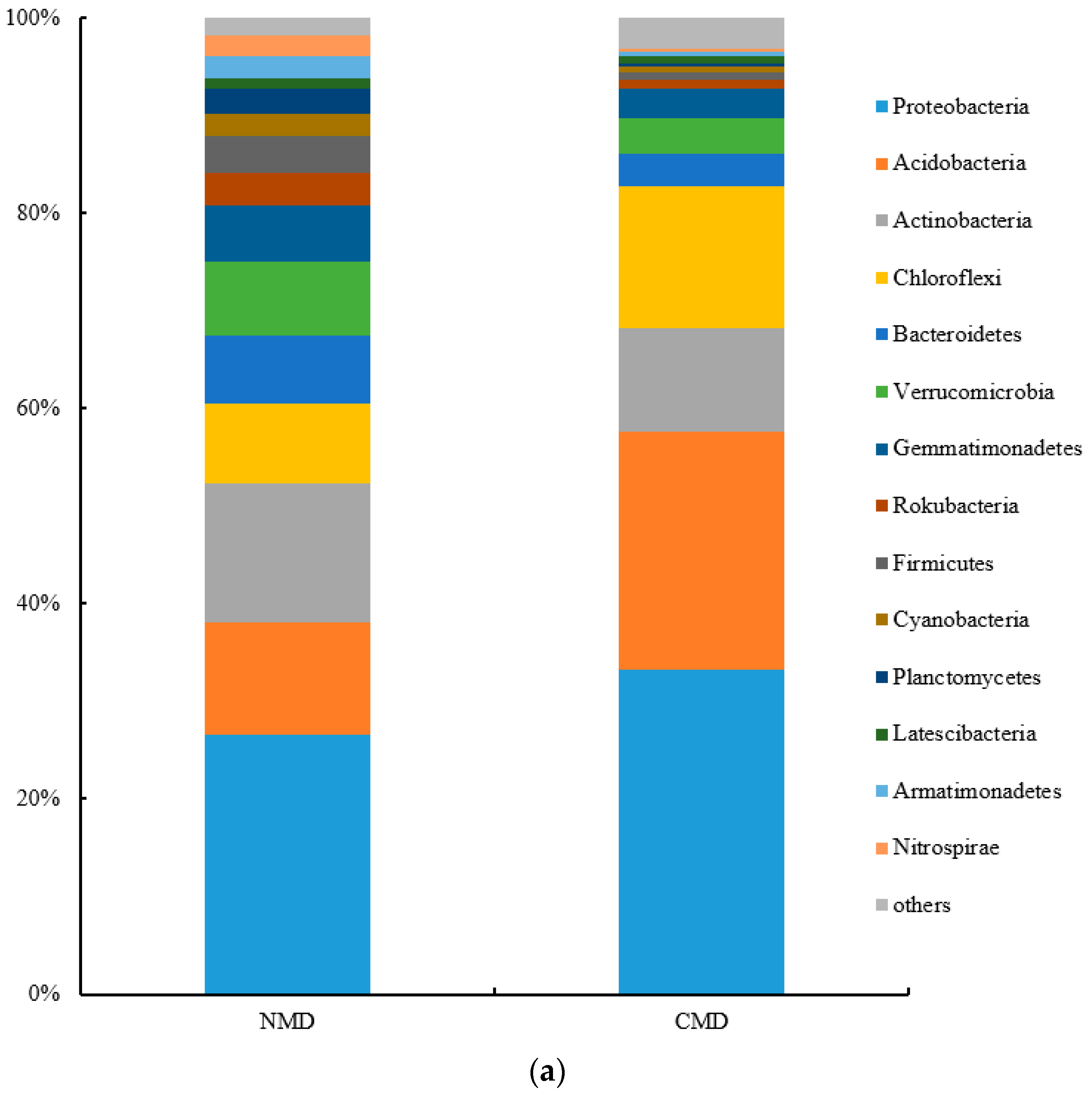
3.3. Microbial Community Structure in Soil Samples of NMD and CMD
3.4. OTU Distribution Characteristics Analysis
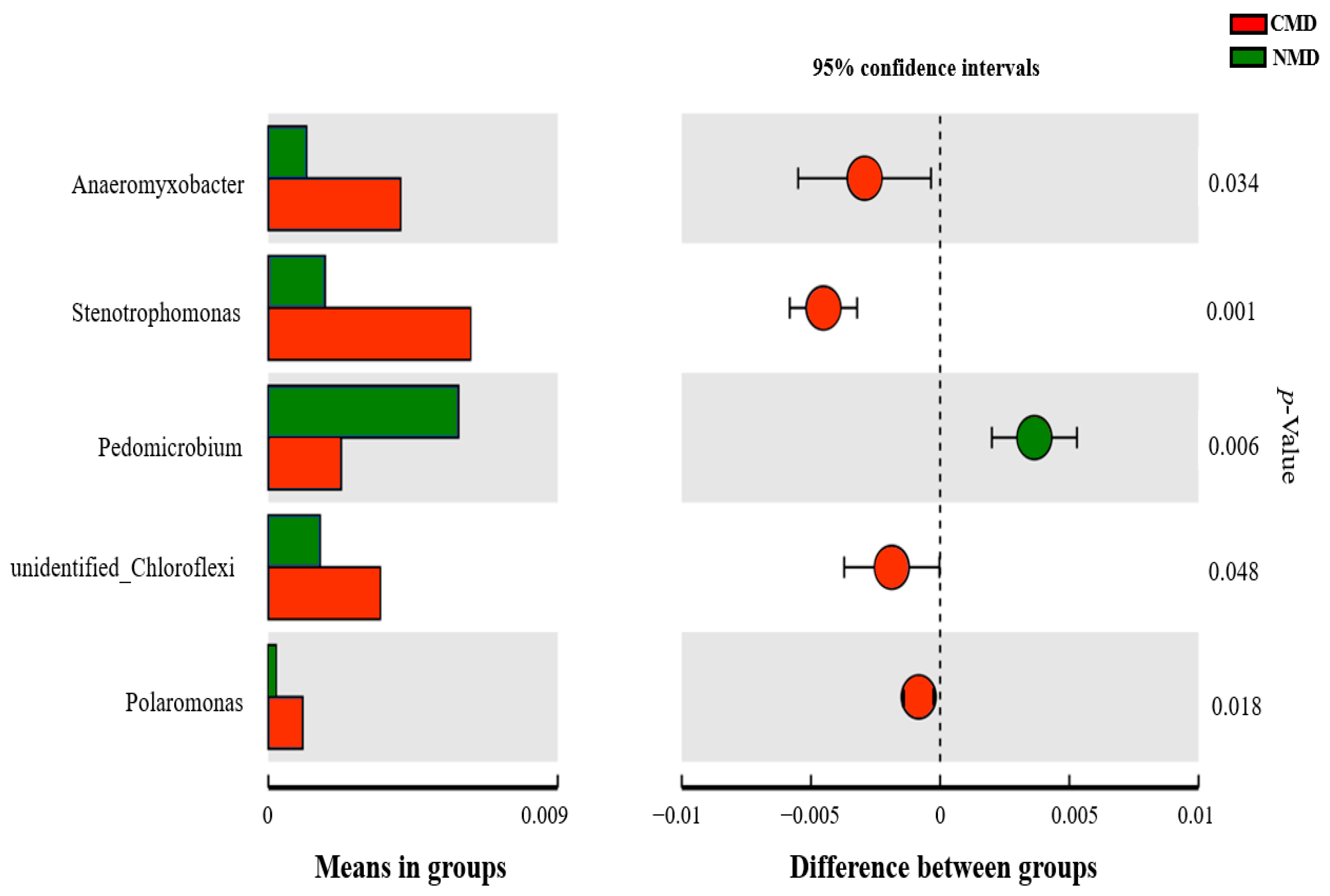
3.5. Difference Analysis of Soil Microbial Flora Abundance
3.6. Correlation Analysis between Microbial Community Structure and Environmental Factors
4. Discussion
4.1. Effects on Soil Microbial Diversity of Captive Forest Musk Deer
4.2. Relationship between Soil Physicochemical Property and Microbial Diversity
4.3. Analysis of the Soil Microbial Community for Captive and Non-Captive Forest Musk Deer Soils
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, J.Y.; Wang, W. The Musk Deer of China; The China Forestry Publishing House: Beijing, China, 2006. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/ (accessed on 1 January 2022).
- Sheng, H.L.; Liu, Z.Y. The Musk Deer in China; The Shanghai Scientific & Technical Publishers: Shanghai, China, 2007. [Google Scholar]
- Liu, K.; Xie, L.; Deng, M.; Zhang, X.M.; Luo, J.; Li, X.F. Zoology, chemical composition, pharmacology, quality control and future perspective of Musk (Moschus): A review. Chin. Med. 2021, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Li, W.; Jin, J.; Cui, K.; Yan, C.; Peng, C.; Jian, Z.; Bu, P.; Price, M.; Zhang, X.; et al. The draft genome sequence of forest musk deer (Moschus berezovskii). Gigascience 2018, 7, giy038. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Meng, X.; Xia, L.; Feng, Z. Conservation status and causes of decline of musk deer (Moschus spp.) in China. Biol. Conserv. 2003, 109, 333–342. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, X. The current status and disease occurrence of captive forest musk deer (Moschus berezovskii) in Shaanxi Province. Chin. J. Ecol. 2014, 10, 2720–2726. [Google Scholar]
- Cai, Y.; Yang, J.; Wang, J.; Yang, Y.; Fu, W.; Zheng, C.; Cheng, J.; Zeng, Y.; Zhang, Y.; Xu, L.; et al. Changes in the Population Genetic Structure of Captive Forest Musk Deer (Moschus berezovskii) with the Increasing Number of Generation under Closed Breeding Conditions. Animals 2020, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zheng, X.; Wang, H.; Qi, H.; Jiang, B.; Qiao, M.; Zhou, J.; Bu, S. Analysis of Genetic Diversity and Population Structure in Three Forest Musk Deer Captive Populations with Different Origins. G3 Genes Genomes Genet. 2019, 9, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ren, Z.; Luo, Y.; Cheng, J.; Wang, J.; Wang, Y.; Yang, Z.; Yao, X.; Zhong, Z.; Yang, W.; et al. Metagenomics analysis of the gut microbiome in healthy and bacterial pneumonia forest musk deer. Genes Genom. 2021, 43, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Zhang, T.X.; Shi, M.H.; Zhang, B.F.; Hu, X.; Xu, S.H.; Ding, J.H.; Liu, S.Q.; Hu, D.F.; Daniel, R. Characterization of intestinal microbiota and fecal cortisol, T3, and IgA in forest musk deer (Moschus berezovskii) from birth to weaning. Integr. Zool. 2021, 16, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Huang, W.; Sun, Y.; You, L.; Jin, H.; Sun, Z. Effect of probiotics on diversity and function of gut microbiota in Moschus berezovskii. Arch. Microbiol. 2021, 203, 3305–3315. [Google Scholar] [CrossRef]
- Liu, C.; Hong, T.; Wang, S.; Dong, X.; Ren, Z. Research Progress on Molecular Mechanism of Musk Secretion in Forest Musk Deer. Chin. J. Zool. 2022, 57, 152–158. [Google Scholar]
- Guo, Y.; Zhou, Y.; Cai, Y.; Cheng, J.; Fu, W.; Long, D.; Meng, X.; Li, W.; Tan, F. Study on the Musk Secretion and Influencing Variables in Captive Male Forest Musk Deer in West Plateau of Sichuan Province. J. Sichuan Agric. Univ. 2018, 36, 273–278. [Google Scholar]
- Bai, K.; Ren, Z.; Wang, Y.; Li, F.; Li, Y.; Zhu, C.; Tang, J.; Tang, Q.; Liu, W.; Wang, F. An association with gonadal hormone level and musk yield in Moschus berezovskii during musk-secreting period. Chin. J. Vet. Sci. 2013, 33, 956–962. [Google Scholar]
- Gao, Y.; Fu, Y.; Yan, L.; Hu, D.; Jiang, B.; Zhang, D. First record of traumatic myiasis obtained from forest musk deer (Moschus berezovskii). Int. J. Parasitol. Parasites Wildl. 2021, 16, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cai, R.; Jin, X.; Shafer, A.B.A.; Hu, X.; Yang, S.; Li, Y.; Qi, L.; Liu, S.; Hu, D. Blood transcriptomics of captive forest musk deer (Moschus berezovskii) and possible associations with the immune response to abscesses. Sci. Rep. 2018, 8, 599. [Google Scholar] [CrossRef]
- Han, H.; Cai, Y.; Cheng, J.; Fu, W.; Sheng, Y.; Guo, F.; Zhou, Y.; Meng, X. Study of the Behavioral Characteristics and the Influence Factors Envolved in Captive Forest Musk Deer (Moschus berezovskii). J. Sichuan Agric. Univ. 2019, 37, 116–121. [Google Scholar]
- Wu, S.Z.; Yang, L.F.; Qian, L.M.; Yang, M.H.; Li, Q.H.; Chen, P.F. Isolation and Identification of Pathogenic Bacteria for Abscess in Captive Forest Musk Deer (Moschus berezovskii). Chin. J. Wildl. 2021, 42, 872–878. [Google Scholar]
- Drenovsky, R.E.; Steenwerth, K.L.; Jackson, L.E.; Scow, K.M. Disease Suppressive Soils: New Insights from the Soil Microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar]
- He, L.; Li, L.h.; Wang, W.X.; Liu, G.; Liu, S.Q.; Liu, W.H.; Hu, D.F. Welfare of farmed musk deer: Changes in the biological characteristics of musk deer in farming environments. Appl. Anim. Behav. Sci. 2014, 156, 1–5. [Google Scholar] [CrossRef]
- Drenovsky, R.; Steenwerth, K.L.; Jackson, L.E.; Scow, K.M. Land use and climatic factors structure regional patterns in soil microbial communities. Glob. Ecol. Biogeogr. 2010, 19, 27–39. [Google Scholar] [CrossRef]
- Lear, G.; Bellamy, J.; Case, B.S.; Lee, J.E.; Buckley, H.L. Fine-scale spatial patterns in bacterial community composition and function within freshwater ponds. ISME J. 2014, 8, 1715–1726. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.J.; Hou, Q.C.; Xu, H.Y.; Sun, Z.H.; Zhang, H.P.; Sun, T.S. Comparative analysis of human and animal gut microbiota diversity and soil microbiota diversity in Xilinguole pasture. Chin. Sci. Bull. 2019, 64, 337–347. [Google Scholar] [CrossRef]
- Wang, W.X.; Zhou, R.; He, L.; Liu, S.Q.; Zhou, J.T.; Qi, L.; Li, L.H.; Hu, D.F. The progress in nutrition research of musk deer: Implication for conservation. Appl. Anim. Behav. Sci. 2015, 172, 1–8. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter, in: Methods of Soil Analysis Part 2. Chem. Microb. Prop. 1982, 9, 539–579. [Google Scholar]
- Hall, D.O.; Coombs, J. Techniques in Bioproductivity and Photosynthesis, 2nd ed.; Pergamon: New York, NY, USA, 1985. [Google Scholar]
- Ye, X.F.; Liu, G.S.; Ling, A.F.; Shi, H.Z.; Li, Y.J.; Zhu, H.B.; Li, S.Q. Canonical correlation between contents of megastigmatrienone in flue cured tobacco leaves and soil physical properties and nutrient conditions. Acta Ecol. Sin. 2009, 29, 4223–4230. [Google Scholar]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schoning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef]
- Cheng, J.G.; Ji, C.H.; Luo, Y.; Zhou, X. Microbial Community Characteristics of Typical Vegetation Soil at Forest Musk Deer Habitats in Sichuan Jiulong Mountain Nature Reserve. J. Northwest. For. Univ. 2017, 32, 205–211. [Google Scholar]
- Shen, G.S.; Zhang, H.; Xia, Q.; Qian, Z.Q.; Nie, W.L.; Zhang, S. Influence of Deer Grazing on Plant Community Structure and Species Diversity. Acta Ecol. Anim. Domastici 2009, 30, 86–90. [Google Scholar]
- Meng, M.J.; Lin, J.; Guo, X.P.; Liu, X.; Wu, J.S.; Zhao, Y.P.; Zhang, J.Z. Impacts of forest conversion on soil bacterial community composition and diversity in subtropical forests. Catena 2019, 175, 167–173. [Google Scholar] [CrossRef]
- Gao, X.F.; Wu, C.Y.; Han, G.D. Steppe Soil Microorganisms Is Impacted by the Grazing and Their Seasonal Changes. Microbiol. China 2010, 37, 1117–1122. [Google Scholar]
- Shu, B.J.; Zhang, X.P.; Jia, S.X. The Effect of Soil Physical and Chemical Properties on Soil Microbial Community in Agro-ecosystem. Siol Crop. 2013, 2, 138–144. [Google Scholar]
- Suriyavirun, N.; Krichels, A.H.; Kent, A.D.; Yang, W.H. Microtopographic differences in soil properties and microbial community composition at the field scale. Soil Biol. Biochem. 2019, 131, 71–80. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Fei, Y.J.; Wu, L.; Yang, C.D. Effects of elk grazing on soil physical and chemical properties of grassland on Tiane Island. Acta Pratacult. Sin. 2010, 19, 115–121. [Google Scholar]
- Marcos, M.S.; Bertiller, M.B.; Olivera, N.L. Microbial community composition and network analyses in arid soils of the Patagonian Monte under grazing disturbance reveal an important response of the community to soil particle size. Appl. Soil Ecol. 2019, 138, 223–232. [Google Scholar] [CrossRef]
- Tang, J.; Xu, Q.R.; Wang, L.M.; Ding, X.; Tang, B.; Wu, L.S.; Feng, S.; Sun, Q.; Yang, Z.R.; Zhang, J. Soil bacterial community diversity under different stages of degradation in zoige wetland. Microbiol. China 2011, 38, 677–686. [Google Scholar]
- Chen, D.D.; Sun, D.S.; Zhang, S.H.; Tan, Y.R.; Du, G.Z.; Shi, X.M. Effect of grazing intensity on soil microbial characteristics of an alpine meadow on the Tibetan plateau. J. Lanzhou Univ. Nat. Sci. 2011, 47, 73–77. [Google Scholar]
- Quoreshi, A.M.; Suleiman, M.K.; Kumar, V.; Manuvel, A.J.; Sivadasan, M.T.; Islam, M.A.; Khasa, D.P. Untangling the bacterial community composition and structure in selected Kuwait desert soils. Appl. Soil Ecol. 2019, 138, 1–9. [Google Scholar] [CrossRef]
- Zhao, S.C.; Qiu, S.J.; Xu, X.P.; Ciampitti, I.A.; Zhang, S.Q.; He, P. Change in straw decomposition rate and soil microbial community composition after straw addition in different long-term fertilization soils. Appl. Soil Ecol. 2019, 138, 123–133. [Google Scholar] [CrossRef]
- Arefieva, O.; Nazarkina, A.V.; Gruschakova, N.V.; Skurikhina, J.E.; Kolycheva, V.B. Impact of mine waters on chemical composition of soil in the Partizansk Coal Basin, Russia. Int. Soil Water Conserv. Res. 2019, 7, 57–63. [Google Scholar] [CrossRef]
- Zhan, P.F.; Xiao, D.R.; Yan, P.F.; Liu, Z.Y.; Ma, J.C.; Chen, Z.M.; Ge, R.; Tian, W.; Wang, H. Soil Degradation-Associated Microbial Community Structure Changes in an Alpine Meadow Under Tibetan Pig Herding. Environ. Sci. 2018, 39, 1840–1850. [Google Scholar]
- Wang, S.K.; Zhao, X.Y.; Zhao, H.; Guo, Y.R.; Yun, J.Y. Characteristic of microbe numbers in sandy grassland under different grazing intensities after years of post-grazing restoration. J. Arid. Land Resour. Environ. 2008, 22, 164–167. [Google Scholar]
- Li, Y.M.; Hu, X.L.; Yang, S.; Zhou, J.T.; Qi, L.; Sun, X.N.; Fan, M.Y.; Xu, S.H.; Cha, M.H.; Zhang, M.S.; et al. Comparison between the Fecal Bacterial Microbiota of Healthy and Diarrheic Captive Musk Deer. Front. Microbiol. 2018, 9, 300. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | NMD | CMD | p-Value | One-Way ANOVA |
|---|---|---|---|---|
| pH | 7.42 ± 0.130 | 7.10 ± 0.158 | 0.020 | ** |
| Organic matter (%) | 0.93 ± 0.055 | 1.19 ± 0.047 | 0.002 | ** |
| Total N (%) | 0.69 ± 0.053 | 0.89 ± 0.031 | <0.001 | ** |
| P (mg/kg) | 353.9 ± 20.6 | 454.6 ± 26.37 | <0.001 | ** |
| K (mg/kg) | 4032.6 ± 58.3 | 6096.0 ± 65.8 | <0.001 | ** |
| Ca (mg/kg) | 3297.9 ± 214.9 | 3166.9 ± 154.9 | 0.463 | NS |
| Mg (mg/kg) | 6728.9 ± 259.5 | 11,235.6 ± 262.5 | <0.001 | ** |
| Zn (mg/kg) | 56.7 ± 6.43 | 68.3 ± 14.3 | 0.168 | NS |
| Fe (mg/kg) | 14,117.4 ± 1992.6 | 17,986.0 ± 2685.3 | 0.137 | * |
| Na (mg/kg) | 338.7 ± 15.6 | 107.4 ± 12.2 | <0.001 | ** |
| Sample | No. of OTUs | Shannon Index | Simpson Index | Chao | Observed-Species | PD-Whole-Tree |
|---|---|---|---|---|---|---|
| NMD | 1518 ± 65 | 7.01 ± 0.68 | 0.807 ± 0.02 | 1504.76 ± 70.34 | 1360.2 ± 164.48 | 143.92 ± 20.74 |
| CMD | 1959 ± 97 | 8.99 ± 0.38 | 0.989 ± 0.01 | 2080.19 ± 94.70 | 1955.80 ± 128.30 | 168.16 ± 14.94 |
| p-value | 0.002 | 0.01 | <0.001 | 0.001 | 0.003 | 0.03 |
| one-way ANOVA | ** | ** | ** | ** | ** | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Suo, L.; Li, F.; Bian, K.; Wang, Q.; Wang, Y.; Lei, Y.; Yang, C. Effects of Breeding Forest Musk Deer on Soil Bacterial Community Structure. Sustainability 2022, 14, 10307. https://doi.org/10.3390/su141610307
Tang J, Suo L, Li F, Bian K, Wang Q, Wang Y, Lei Y, Yang C. Effects of Breeding Forest Musk Deer on Soil Bacterial Community Structure. Sustainability. 2022; 14(16):10307. https://doi.org/10.3390/su141610307
Chicago/Turabian StyleTang, Jie, Lijuan Suo, Feiran Li, Kun Bian, Qingxiong Wang, Yan Wang, Yinhu Lei, and Chao Yang. 2022. "Effects of Breeding Forest Musk Deer on Soil Bacterial Community Structure" Sustainability 14, no. 16: 10307. https://doi.org/10.3390/su141610307
APA StyleTang, J., Suo, L., Li, F., Bian, K., Wang, Q., Wang, Y., Lei, Y., & Yang, C. (2022). Effects of Breeding Forest Musk Deer on Soil Bacterial Community Structure. Sustainability, 14(16), 10307. https://doi.org/10.3390/su141610307