
Toward Feeds for Circular Multitrophic Food Production Systems: Holistically Evaluating Growth Performance and Nutrient Excretion of African Catfish Fed Fish Meal-Free Diets in Comparison to Nile Tilapia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Preparation and Chemical Analysis
2.3. Calculations
2.4. Statistical Analysis
3. Results
3.1. Fish Rearing and Performance
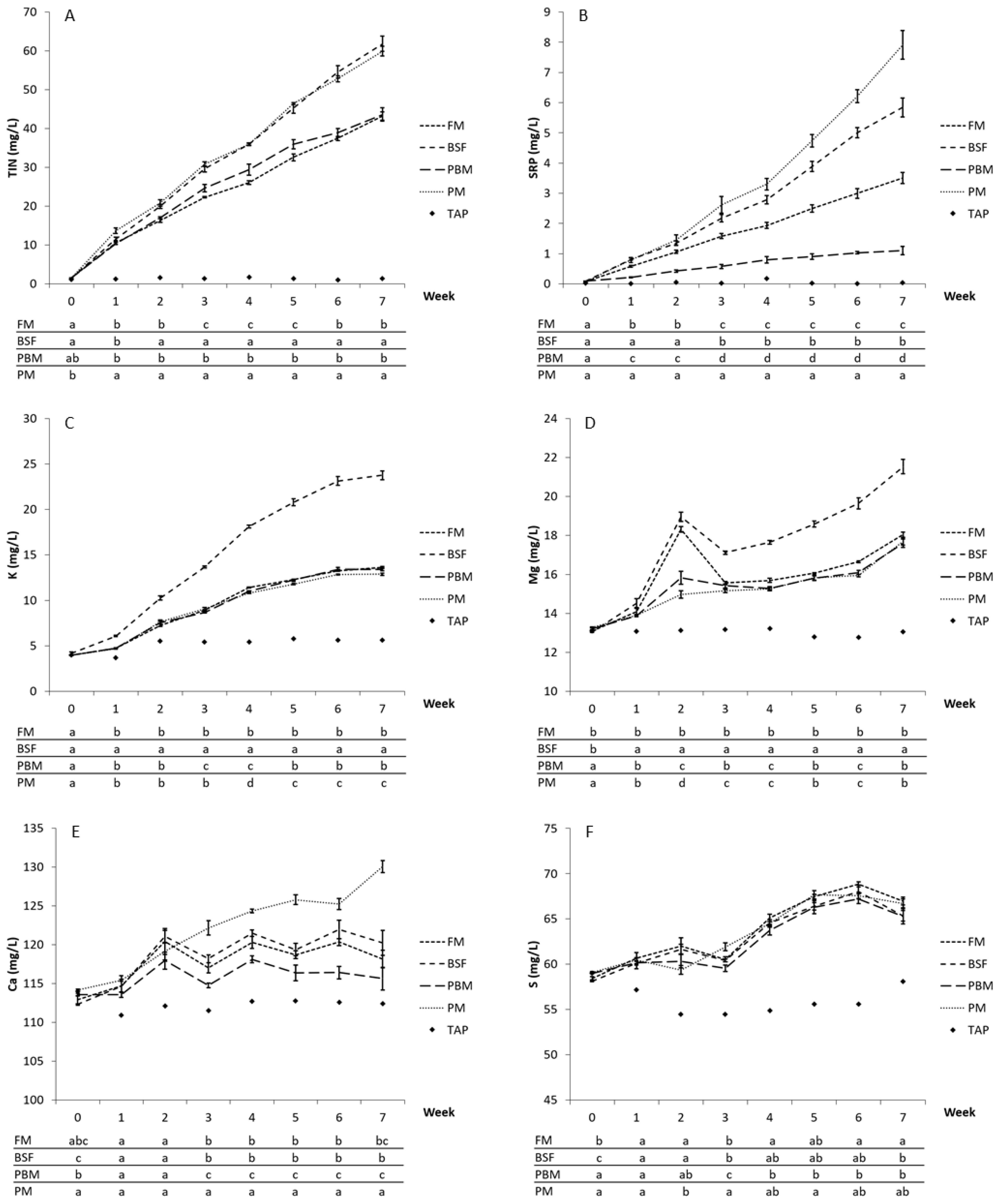
3.2. Water—Dissolved Nutrient Excretion Patterns
3.3. Feces—Solid Nutrient Excretion Patterns
3.3.1. Proximate Composition and Nutrient Recovery
3.3.2. Minerals and Amino Acids
3.4. Comparison between African Catfish and Nile Tilapia
3.4.1. Growth Performance
3.4.2. Production of Dissolved Plant Macronutrients per g of Feed—TIN, SRP and K
3.4.3. Feces Nutrient Composition and DM Recovery
4. Discussion
4.1. Fish Performance
4.2. Water—Implictions for Plant Production
4.3. Fish Feces—A Resource in CMFS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.I.M.; Eding, E.H.; Verdegem, M.C.J.; Heinsbroek, L.T.N.; Schneider, O.; Blancheton, J.P.; D’Orbcastel, E.R.; Verreth, J.A.J. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability. Aquac. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, J.M.; Timmons, M.B. Recirculating aquaculture systems. In Aquaculture Production Systems; Tidwell, J.H., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 245–277. [Google Scholar]
- Badiola, M.; Mendiola, D.; Bostock, J. Recirculating aquaculture systems (RAS) analysis: Main issues on management and future challenges. Aquac. Eng. 2012, 51, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Baulcombe, D.; Crute, I.; Davies, B.; Dunwell, J.; Gale, M.; Jones, J.; Pretty, J.; Sutherland, W.; Toulmin, C. Reaping the Benefits—Science and the Sustainable Intensification of Global Agriculture; The Royal Society: London, UK, 2009. [Google Scholar]
- Baganz, G.F.M.; Junge, R.; Portella, M.C.; Goddek, S.; Keesman, K.J.; Baganz, D.; Staaks, G.; Shaw, C.; Lohrberg, F.; Kloas, W. The aquaponic principle—It is all about coupling. Rev. Aquac. 2022, 14, 252–264. [Google Scholar] [CrossRef]
- Kloas, W.; Groß, R.; Baganz, D.; Graupner, J.; Monsees, H.; Schmidt, U.; Staaks, G.; Suhl, J.; Tschirner, M.; Wittstock, B.; et al. A new concept for aquaponic systems to improve sustainability, increase productivity, and reduce environmental impacts. Aquac. Environ. Interact. 2015, 7, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Monsees, H.; Kloas, W.; Wuertz, S. Decoupled systems on trial: Eliminating bottlenecks to improve aquaponic processes. PLoS ONE 2017, 12, e0183056. [Google Scholar] [CrossRef] [Green Version]
- Eck, M.; Körner, O.; Jijakli, M.H. Nutrient cycling in aquaponics systems. In Aquaponics Food Production Systems; Goddek, S., Joyce, A., Kotzen, B., Burnell, G., Eds.; Springer: Cham, Switzerland, 2019; pp. 231–246. [Google Scholar]
- Stegmann, P.; Londo, M.; Junginger, M. The circular bioeconomy: Its elements and role in European bioeconomy clusters. Resour. Conserv. Recycl. X 2020, 6, 100029. [Google Scholar] [CrossRef]
- Carus, M.; Dammer, L. The “circular bioeconomy”—Concepts, opportunities and limitations. Ind. Biotechnol. 2018, 14, 83–91. [Google Scholar] [CrossRef]
- Davis, C.B.; Aid, G.; Zhu, B. Secondary resources in the bio-based economy: A computer assisted survey of value pathways in academic literature. Waste Biomass Valoriz. 2017, 8, 2229–2246. [Google Scholar] [CrossRef]
- Ulrichs, C. CUBES Circle. Available online: https://www.cubescircle.de/en/home/project (accessed on 25 January 2022).
- Shaw, C.; Knopf, K.; Kloas, W. Fish feeds in aquaponics and beyond: A novel concept to evaluate protein sources in diets for circular multitrophic food production systems. Sustainability 2022, 14, 4064. [Google Scholar] [CrossRef]
- Villarroel, M.; Junge, R.; Komives, T.; König, B.; Plaza, I.; Bittsánszky, A.; Joly, A. Survey of aquaponics in Europe. Water 2016, 8, 468. [Google Scholar] [CrossRef] [Green Version]
- Love, D.C.; Fry, J.P.; Li, X.; Hill, E.S.; Genello, L.; Semmens, K.; Thompson, R.E. Commercial aquaponics production and profitability: Findings from an international survey. Aquaculture 2015, 435, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Rakocy, J.E.; Masser, M.P.; Losordo, T.M. Recirculating Aquaculture Tank Production Systems: Aquaponics—Integrating Fish and Plant Culture; SRAC Publication No. 454; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2006. [Google Scholar]
- Yep, B.; Zheng, Y. Aquaponic trends and challenges–A review. J. Clean. Prod. 2019, 228, 1586–1599. [Google Scholar] [CrossRef]
- Popma, T.J.; Lovshin, L.L. Worldwide Prospects for Commercial Production of Tilapia; International Center for Aquaculture and Aquatic Environments Department of Fisheries and Allied Aquacultures Auburn University: Auburn, AL, USA, 1996. [Google Scholar]
- El-Sayed, A.-F.M. Tilapia Culture; CABI Publishing, CABI International: Willingford, UK, 2006. [Google Scholar]
- Haylor, G.S. The case for the African catfish, Clarias gariepinus Burchell, 1822, Clariidae: A comparison of the relative merits of tilapiine fishes, especially Oreochromis niloticus (L.) and C. gariepinus Burchell, for African aquaculture. Aquac. Res. 1989, 20, 279–285. [Google Scholar] [CrossRef]
- Pasch, J.; Palm, H.W. Economic analysis and improvement opportunities of African catfish (Clarias gariepinus) aquaculture in northern Germany. Sustainability 2021, 13, 3569. [Google Scholar] [CrossRef]
- Huisman, E.A.; Richter, C.J.J. Reproduction, growth, health control and aquacultural potential of the African catfish, Clarias gariepinus (Burchell 1822). Aquaculture 1987, 63, 1–14. [Google Scholar] [CrossRef]
- Hecht, T.; Uys, W.; Britz, P.J. The Culture of Shartooth Catfish, Clarias gariepinus in Southern Africa; South African National Scientific Programmes Report No. 153; National Scientific Programmes Unit (CSIR): Pretoria, South Africa, 1988. [Google Scholar]
- Bovendeur, J.; Eding, E.H.; Henken, A.M. Design and performance of a water recirculation system for high-density culture of the African catfish, Clarias gariepinus (Burchell 1822). Aquaculture 1987, 63, 329–353. [Google Scholar] [CrossRef]
- Endut, A.; Jusoh, A.; Ali, N.; Wan Nik, W.B.; Hassan, A. A study on the optimal hydraulic loading rate and plant ratios in recirculation aquaponic system. Bioresour. Technol. 2010, 101, 1511–1517. [Google Scholar] [CrossRef]
- Endut, A.; Jusoh, A.; Ali, N.; Wan Nik, W.N.S.; Hassan, A. Effect of flow rate on water quality parameters and plant growth of water spinach (Ipomoea aquatica) in an aquaponic recirculating system. Desalin. Water Treat. 2009, 5, 19–28. [Google Scholar] [CrossRef]
- Suhl, J.; Dannehl, D.; Baganz, D.; Schmidt, U.; Kloas, W. An innovative suction filter device reduces nitrogen loss in double recirculating aquaponic systems. Aquac. Eng. 2018, 82, 63–72. [Google Scholar] [CrossRef]
- Oladimeji, S.A.; Okomoda, V.T.; Olufeagba, S.O.; Solomon, S.G.; Abol-Munafi, A.B.; Alabi, K.I.; Ikhwanuddin, M.; Martins, C.O.; Umaru, J.; Hassan, A. Aquaponics production of catfish and pumpkin: Comparison with conventional production systems. Food Sci. Nutr. 2020, 8, 2307–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diatin, I.; Shafruddin, D.; Hude, N.; Sholihah, M.; Mutsmir, I. Production performance and financial feasibility analysis of farming catfish (Clarias gariepinus) utilizing water exchange system, aquaponic, and biofloc technology. J. Saudi Soc. Agric. Sci. 2021, 20, 344–351. [Google Scholar] [CrossRef]
- Knaus, U.; Zimmermann, J.; Appelbaum, S.; Palm, H.W. Spearmint (Mentha spicata) cultivation in decoupled aquaponics with three hydro-components (grow pipes, raft, gravel) and African catfish (Clarias gariepinus) production in northern Germany. Sustainability 2022, 14, 305. [Google Scholar] [CrossRef]
- Siqwepu, O.; Salie, K.; Goosen, N. Evaluation of chelated iron and iron sulfate in the diet of African catfish, Clarias gariepinus to enhance iron excretion for application in integrated aquaponics systems. J. World Aquac. Soc. 2020, 51, 1034–1053. [Google Scholar] [CrossRef]
- Siqwepu, O.; Salie, K.; Goosen, N. Evaluation of potassium diformate and potassium chloride in the diet of the African catfish, Clarias gariepinus in a recirculating aquaculture system. Aquaculture 2020, 526, 735414. [Google Scholar] [CrossRef]
- Pasch, J.; Appelbaum, S.; Palm, H.W.; Knaus, U. Growth of basil (Ocimum basilicum) in aeroponics, DRF, and raft systems with effluents of African catfish (Clarias gariepinus) in decoupled aquaponics (s.s.). AgriEngineering 2021, 3, 559–574. [Google Scholar] [CrossRef]
- Baßmann, B.; Brenner, M.; Palm, H.W. Stress and welfare of African catfish (Clarias gariepinus Burchell, 1822) in a coupled aquaponic system. Water 2017, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Baßmann, B.; Harbach, H.; Weißbach, S.; Palm, H.W. Effect of plant density in coupled aquaponics on the welfare status of African catfish, Clarias gariepinus. J. World Aquac. Soc. 2020, 51, 183–199. [Google Scholar] [CrossRef]
- Alltech Coppens. RAS-Zucht von Welsen—Einen Blick hinter die Kulissen. Available online: https://www.alltechcoppens.com/de/aktuelles/farming-catfish-in-ras (accessed on 14 September 2022).
- van de Nieuwegiessen, P.G.; Olwo, J.; Khong, S.; Verreth, J.A.J.; Schrama, J.W. Effects of age and stocking density on the welfare of African catfish, Clarias gariepinus Burchell. Aquaculture 2009, 288, 69–75. [Google Scholar] [CrossRef]
- Palm, H.W.; Knaus, U.; Wasenitz, B.; Bischoff, A.A.; Strauch, S.M. Proportional up scaling of African catfish (Clarias gariepinus Burchell, 1822) commercial recirculating aquaculture systems disproportionally affects nutrient dynamics. Aquaculture 2018, 491, 155–168. [Google Scholar] [CrossRef]
- Conceição, L.E.C.; Houlihan, D.F.; Verreth, J.A.J. Fast growth, protein turnover and costs of protein metabolism in yolk-sac larvae of the African catfish (Clarias gariepinus). Fish Physiol. Biochem. 1997, 16, 291–302. [Google Scholar] [CrossRef]
- Conceição, L.E.C.; Dersjant-Li, Y.; Verreth, J.A.J. Cost of growth in larval and juvenile African catfish (Clarias gariepinus) in relation to growth rate, food intake and oxygen consumption. Aquaculture 1998, 161, 95–106. [Google Scholar] [CrossRef]
- Belão, T.C.; Leite, C.A.C.; Florindo, L.H.; Kalinin, A.L.; Rantin, F.T. Cardiorespiratory responses to hypoxia in the African catfish, Clarias gariepinus (Burchell 1822), an air-breathing fish. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2011, 181, 905–916. [Google Scholar] [CrossRef] [PubMed]
- van de Nieuwegiessen, P.G.; Boerlage, A.S.; Verreth, J.A.J.; Schrama, J.W. Assessing the effects of a chronic stressor, stocking density, on welfare indicators of juvenile African catfish, Clarias gariepinus Burchell. Appl. Anim. Behav. Sci. 2008, 115, 233–243. [Google Scholar] [CrossRef]
- Haylor, G.S. Controlled hatchery production of Clarias gariepinus (Burchell 1822): Growth and survival of fry at high stocking density. Aquac. Fish. Manag. 1991, 22, 405–422. [Google Scholar] [CrossRef]
- Roques, J.A.C.; Schram, E.; Spanings, T.; van Schaik, T.; Abbink, W.; Boerrigter, J.; de Vries, P.; van de Vis, H.; Flik, G. The impact of elevated water nitrite concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2015, 46, 1384–1395. [Google Scholar] [CrossRef]
- Schram, E.; Roques, J.A.C.; Abbink, W.; Yokohama, Y.; Spanings, T.; de Vries, P.; Bierman, S.; van de Vis, H.; Flik, G. The impact of elevated water nitrate concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2014, 45, 1499–1511. [Google Scholar] [CrossRef]
- Ip, Y.K.; Chew, S.F.; Wilson, J.M.; Randall, D.J. Defences against ammonia toxicity in tropical air-breathing fishes exposed to high concentrations of environmental ammonia: A review. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2004, 174, 565–575. [Google Scholar] [CrossRef]
- Ip, Y.K.; Zubaidah, R.M.; Liew, P.C.; Loong, A.M.; Hiong, K.C.; Wong, W.P.; Chew, S.F. African sharptooth catfish Clarias gariepinus does not detoxify ammonia to urea or amino acids but actively excretes ammonia during exposure to environmental ammonia. Physiol. Biochem. Zool. 2004, 77, 242–254. [Google Scholar] [CrossRef]
- FishBase. Oreochromis Niloticus (Linnaeus, 1758)—Nile Tilapia. Available online: https://www.fishbase.se/summary/oreochromis-niloticus.html (accessed on 20 July 2022).
- Tesfahun, A.; Temesgen, M. Food and feeding habits of Nile tilapia Oreochromis niloticus (L.) in Ethiopian water bodies: A review. Int. J. Fish. Aquat. Stud. 2018, 6, 43–47. [Google Scholar]
- Oso, J.A.; Ayodele, I.A.; Fagbuaro, O. Food and feeding habits of Oreochromis niloticus (L.) and Sarotherodon galilaeus (L.) in a tropical reservoir. World J. Zool. 2006, 1, 118–121. [Google Scholar]
- Kadye, W.T.; Booth, A.J. Integrating stomach content and stable isotope analyses to elucidate the feeding habits of non-native sharptooth catfish Clarias gariepinus. Biol. Invasions 2012, 14, 779–795. [Google Scholar] [CrossRef]
- Dadebo, E. Reproductive biology and feeding habits of the catfish in Lake Awassa, Ethiopia. Ethiop. J. Sci. 2000, 23, 231–246. [Google Scholar] [CrossRef] [Green Version]
- FishBase. Clarias gariepinus (Burchell, 1822)—North African catfish. Available online: https://www.fishbase.se/summary/1934# (accessed on 20 July 2022).
- Phrompanya, P.; Saenphet, K.; Saenphet, S. Comparative histochemical study of the gastrointestinal tracts of the Nile tilapia (Oreochromis niloticus) and the hybrid catfish (Clarias batrachus × Clarias gariepinus). Acta Histochem. 2019, 121, 261–267. [Google Scholar] [CrossRef] [PubMed]
- El Nahass, E.E.M.; Elwan, M.M.M. Comparitive histological, histochemical and ultrastructural studies on the ileum of African catfish (Clarias gariepinus) and (Oreochromis niloticus). Egypt. J. Exp. Biol. 2019, 15, 295–303. [Google Scholar] [CrossRef]
- Elwan, M.M.M.; El Nahass, E.E.S.M. Comparitive histological and ultrastructural studies on the stomach of African catfish (Clarias gariepinus) and (Oreochromis niloticus). J. Histol. Histopathol. 2019, 6, 7243. [Google Scholar] [CrossRef] [Green Version]
- Rust, M.B. Nutritional physiology. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 367–452. [Google Scholar]
- Lall, S.P. The minerals. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 259–308. [Google Scholar]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Bureau, D.P.; Kaushik, S.J.; Cho, C.Y. Bioenergetics. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 1–59. [Google Scholar]
- Kleiber, M. The Fire of Life. An Introduction to Animal Energetics; Krieger Publishing: New York, NY, USA, 1975. [Google Scholar]
- DIN EN ISO 16634-2; Food Products—Determination of the Total Nitrogen Content by Combustion according to the Dumas Principle and Calculation of the Crude Protein Content—Part 2: Cereals, Pulses and Milled Cereal Products. DIN Deutsches Institut für Normung e.V.: Berlin, Germany, 2016.
- EC (European Commission). Commission Regulation (EC) No 152/2009 of 27 January 2009 Laying Down the Methods of Sampling and Analysis for the Official Control of Feed; European Commission: Brussels, Belgium, 2009. [Google Scholar]
- DIN EN ISO 13903; Animal Feeding Stuffs—Determination of Amino Acids Content. DIN Deutsches Institut für Normung e.V.: Berlin, Germany, 2005.
- FOOD-PA 662; Bestimmung des Gehaltes an Gesamt-Tryptophan (Summe: Frei und gebunden) in Lebens- und Futtermitteln mittels LC-Aminosäureanalysator mit Ninhydrin. SGS (SGS Analytics GmbH Germany): Berlin, Germany, 2021.
- DIN EN 16174; Sludge, Treated Biowaste and Soil—Digestion of Aqua Regia Soluble Fractions of Elements. DIN Deutsches Institut für Normung e.V.: Berlin, Germany, 2012.
- DIN EN ISO 11885; Water Quality—Determination of Selected Elements by Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES). DIN Deutsches Institut für Normung e.V.: Berlin, Germany, 2009.
- Cho, C.Y. Feeding systems for rainbow trout and other salmonids with reference to current estimates of energy and protein requirements. Aquaculture 1992, 100, 107–123. [Google Scholar] [CrossRef]
- Bureau, D.P. Rendered products in fish aquaculture feeds. In Essential Rendering—All About the Animal By-Products Industry; Meeker, D.L., Ed.; The National Renderers Association (NRA): Alexandria, VA, USA, 2006; pp. 179–184. [Google Scholar]
- Ataguba, G.A.; Kamble, M.T.; Salin, K.R. Food industry by-products as protein replacement in aquaculture diets of tilapia and catfishs. In Food Processing By-Products and their Utilization; Anal, A.K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 471–507. [Google Scholar]
- Luthada-Raswiswi, R.; Mukaratirwa, S.; O’brien, G. Animal protein sources as a substitute for fishmeal in aquaculture diets: A systematic review and meta-analysis. Appl. Sci. 2021, 11, 3854. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.S.C.; Wilson, A.E.; Davis, D.A. Success of fishmeal replacement through poultry by-product meal in aquaculture feed formulations: A meta-analysis. Rev. Aquac. 2020, 12, 1624–1636. [Google Scholar] [CrossRef]
- El-Husseiny, O.M.; Issoufou, H.M.; Ashraf, S. Determination of the order of amino acid limitation in slaughterhouse poultry by-product meal in African catfish diet by amino acid addition assay. J. Arab. Aquac. Soc. 2013, 8, 373–384. [Google Scholar]
- El-Husseiny, O.M.; Hassan, M.I.; El-Haroun, E.R.; Suloma, A. Utilization of poultry by-product meal supplemented with l-lysine as fish meal replacer in the diet of African catfish Clarias gariepinus (Burchell, 1822). J. Appl. Aquac. 2018, 30, 63–75. [Google Scholar] [CrossRef]
- Goda, A.M.; El-Haroun, E.R.; Kabir Chowdhury, M.A. Effect of totally or partially replacing fish meal by alternative protein sources on growth of African catfish Clarias gariepinus (Burchell, 1822) reared in concrete tanks. Aquac. Res. 2007, 38, 279–287. [Google Scholar] [CrossRef]
- Abdel-Warith, A.A.; Russell, P.M.; Davies, S.J. Inclusion of a commercial poultry by-product meal as a protein replacement of fish meal in practical diets for African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2001, 32, 296–305. [Google Scholar] [CrossRef]
- Falaye, A.E.; Omoike, A.; Ajani, E.K.; Kolawole, O.T. Replacement of fishmeal using poultry offal meal in practical feeds for fry of the African catfish (Clarias gariepinus). Isr. J. Aquac.-Bamidgeh 2011, 63, 1–6. [Google Scholar] [CrossRef]
- Olugbenga, O. Utilisation of a poultry wastes meal as a replacement for fishmeal in diets of Clarias gariepinus. Afr. J. Fish. Aquat. Resour. Manag. 2018, 3, 1–7. [Google Scholar]
- Degani, G.; Revach, A. Digestive capabilities of three commensal fish species: Carp, Cyprinus carpio L., tilapia, Oreochromis aureus × O. niloticus, and African catfish, Clarias gariepinus (Burchell 1822). Aquac. Res. 1991, 22, 397–403. [Google Scholar] [CrossRef]
- García-Pérez, O.D.; Cruz-Valdez, J.C.; Ramírez-Martínez, C.; Villarreal-Cavazos, D.; Gamboa-Delgado, J. Exploring the contribution of dietary protein from poultry by-product meal and fish meal to the growth of catfish Ictalurus punctatus by means of nitrogen stable isotopes. Lat. Am. J. Aquat. Res. 2018, 46, 37–44. [Google Scholar] [CrossRef]
- Fagbenro, O.A.; Balogun, A.M.; Bello-Olusoji, O.A.; Fasakin, E.A. Dietary lysine requirement of the African catfish, Clarias gariepinus. J. Appl. Aquac. 1998, 8, 71–77. [Google Scholar] [CrossRef]
- Fagbenro, O.A.; Balogun, A.M.; Fasakin, E.A. Dietary methionine requirement of the African catfish, Clarias gariepinus. J. Appl. Aquac. 1999, 8, 47–54. [Google Scholar] [CrossRef]
- Elesho, F.E.; Sutter, D.A.H.; Swinkels, M.A.C.; Verreth, J.A.J.; Kröckel, S.; Schrama, J.W. Quantifying methionine requirement of juvenile African catfish (Clarias gariepinus). Aquaculture 2021, 532, 736020. [Google Scholar] [CrossRef]
- Khan, M.A.; Abidi, S.F. Optimum histidine requirement of fry African catfish, Clarias gariepinus (Burchell). Aquac. Res. 2009, 40, 1000–1010. [Google Scholar] [CrossRef]
- Fagbenro, O.A.; Nwanna, L.C. Dietary tryptophan requirement of the African catfish, Clarias gariepinus. J. Appl. Aquac. 1999, 9, 65–72. [Google Scholar] [CrossRef]
- Liland, N.S.; Araujo, P.; Xu, X.X.; Lock, E.; Radhakrishnan, G.; Prabhu, A.J.P.; Belghit, I. A meta-analysis on the nutritional value of insects in aquafeeds. J. Insects Food Feed 2021, 7, 743–759. [Google Scholar] [CrossRef]
- Hua, K. A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 2021, 530, 735732. [Google Scholar] [CrossRef]
- Rapatsa, M.; Moyo, N. A review and meta-analysis of the effects of replacing fishmeal with insect meals on growth of tilapias and sharptooth catfish. Aquac. Nutr. 2022, 2022, 9367587. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Gougbedji, A.; Detilleux, J.; Lalèyè, P.A.; Francis, F.; Caparros Megido, R. Can insect meal replace fishmeal? A meta-analysis of the effects of black soldier fly on fish growth performances and nutritional values. Animals 2022, 15, 1700. [Google Scholar] [CrossRef]
- Priyadarshana, K.C.; Ruwandeepika, D. Substitution of fishmeal with black soldier fly Hermetia illucens Linnaeus, 1758 larvae in finfish aquaculture—A review. Asian Fish. Sci. 2021, 34, 114–126. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Mohd, N.; Firdaus, T.; Hasniyati, A. The effect of dietary cricket meal (Gryllus bimaculatus) on growth performance, antioxidant enzyme activities, and haematological response of African catfish (Clarias gariepinus). Fish Physiol. Biochem. 2016, 42, 1143–1155. [Google Scholar] [CrossRef]
- Taufek, N.M.; Muin, H.; Raji, A.A.; Alias, Z.; Razak, S.A. Potential of field crickets meal (Gryllus bimaculatus) in the diet of African catfish (Clarias gariepinus). J. Appl. Anim. Res. 2018, 2119, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Alegbeleye, W.O.; Obasa, S.O.; Olude, O.O.; Otubu, K.; Jimoh, W. Preliminary evaluation of the nutritive value of the variegated grasshopper (Zonocerus variegatus L.) for African catfish Clarias gariepinus. Aquac. Res. 2012, 43, 412–420. [Google Scholar] [CrossRef]
- Fasakin, E.A.; Balogun, A.M.; Ajayi, O.O. Evaluation of full-fat and defatted maggot meals in the feeding of clariid catfish Clarias gariepinus fingerlings. Aquac. Res. 2003, 34, 733–738. [Google Scholar] [CrossRef]
- Kurbanov, A.R.; Milusheva, R.Y.; Rashidova, S.S.; Kamilov, B.G. Effect of replacement of fish meal with silkworm (Bombyx mori) pupa protein on the growth of Clarias gariepinus fingerling. Int. J. Fish. Aquat. Stud. 2015, 2, 25–27. [Google Scholar]
- Ng, W.K. Potential of mealworm (Tenebrio molifor) as an alternative protein source in practical diets for African catfish, Clarias gariepinus. Aquac. Res. 2001, 32, 273–280. [Google Scholar] [CrossRef]
- Grace, O.I. Effects of grasshopper meal in the diet of Clarias gariepinus fingerlings. J. Aquac. Res. Dev. 2015, 6, 321. [Google Scholar] [CrossRef] [Green Version]
- Taufek, N.M.; Muin, H.; Raji, A.A.; Shaharudin, A.; Yusof, H.M. Apparent digestibility coefficients and amino acid availability of cricket meal, Gryllus bimaculatus, and fishmeal in African catfish, Clarias gariepinus, diet. J. World Aquac. Soc. 2016, 47, 798–805. [Google Scholar] [CrossRef]
- Adeoye, A.A.; Akegbejo-samsons, Y.; Fawole, F.J.; Davies, S.J. Preliminary assessment of black soldier fly (Hermetia illucens) larval meal in the diet of African catfish (Clarias gariepinus): Impact on growth, body index, and hematological parameters. J. World Aquac. Soc. 2020, 51, 1024–1033. [Google Scholar] [CrossRef]
- Azri, N.A.M.; Chun, L.K.; Hasan, H.A.; Jaya-Ram, A.; Kari, Z.A.; Hamid, N.K.A. The effects of partial replacement of fishmeal with Hermetia meal on the growth and fatty acid profile of African catfish fry. Agric. Rep. 2022, 1, 17–27. [Google Scholar]
- Fawole, F.J.; Adeoye, A.A.; Tiamiyu, L.O.; Ajala, K.I.; Obadara, S.O.; Ganiyu, I.O. Substituting fishmeal with Hermetia illucens in the diets of African catfish (Clarias gariepinus): Effects on growth, nutrient utilization, haemato-physiological response, and oxidative stress biomarker. Aquaculture 2020, 518, 734849. [Google Scholar] [CrossRef]
- Gana, A.B.; Tijani, S.; Ibrahim, R. Nutrient utilization, haematological indices and carcass compositions of heteroclarias fingerlings fed Hermetia illucens (Linnaeus, 1758) larvae meal. Int. J. Fish. Aquat. Stud. 2020, 8, 42–48. [Google Scholar]
- Elesho, F.E.; Kröckel, S.; Sutter, D.A.H.; Nuraini, R.; Chen, I.J.; Verreth, J.A.J.; Schrama, J.W. Effect of feeding level on the digestibility of alternative protein-rich ingredients for African catfish (Clarias gariepinus). Aquaculture 2021, 544, 737108. [Google Scholar] [CrossRef]
- Anvo, M.P.M.; Aboua, B.R.D.; Compaoré, I.; Sissao, R.; Zoungrana-Kaboré, C.Y.; Kouamelan, E.P.; Toguyéni, A. Fish meal replacement by Cirina butyrospermi caterpillar’s meal in practical diets for Clarias gariepinus fingerlings. Aquac. Res. 2017, 48, 5243–5250. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Shiau, S.Y.; Yu, Y.P. Dietary supplementation of chitin and chitosan depresses growth in tilapia, Oreochromis niloticus × O. aureus. Aquaculture 1999, 179, 439–446. [Google Scholar] [CrossRef]
- Fontes, T.V.; Oliveira, K.R.B.; Almeida, I.L.G.; Orlando, T.M.; Borges, R.P.; Costa, D.V.; Rosa, P.V. Digestibility of insect meals for Nile tilapia fingerlings. Animals 2019, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Ogunji, J.O.; Iheanacho, S.C.; Abe, G.A.; Ikeh, O.R. Assessing effects of substituting dietary fish meal with boiled donkey and cow blood meal on growth performance and digestive enzyme activities of Clarias gariepinus juvenile. J. World Aquac. Soc. 2020, 51, 1066–1079. [Google Scholar] [CrossRef]
- Ogunji, J.; Iheanacho, S. Alternative protein source: Acceptability of cow blood meal in place of fish meal assessed via growth, antioxidant enzymes functions and haematological response in Clarias gariepinus (Burchell, 1822). Aquac. Res. 2021, 52, 2651–2661. [Google Scholar] [CrossRef]
- Olukunle, O.A.; Ogunsanmi, A.O.; Taiwo, V.O.; Samuel, A.A. The nutritional value of cow blood meal and its effects on growth performance, haematology and plasma enzymes of hybrid catfish. Trop. J. Anim. Sci 2002, 5, 75–85. [Google Scholar] [CrossRef]
- Otubusin, S.O. Effects of different levels of blood meal in pelleted feeds on tilapia, Oreochromis niloticus, production in floating bamboo net-cages. Aquaculture 1987, 65, 263–266. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Total replacement of fish meal with animal protein sources in Nile tilapia, Oreochromis niloticus (L.), feeds. Aquac. Res. 1998, 29, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Kirimi, J.G.; Musalia, L.M.; Magana, A.; Munguti, J.M. Performance of Nile tilapia (Oreochromis niloticus) fed diets containing blood meal as a replacement of fish meal. J. Agric. Sci. 2016, 8, 79–87. [Google Scholar] [CrossRef]
- Nwanna, L.C.; Adebayo, I.A.; Omitoyin, B.O. Phosphorus requirements of African catfish, Clarias gariepinus, based on broken-line regression analysis methods. ScienceAsia 2009, 35, 227–233. [Google Scholar] [CrossRef]
- Nwanna, L.; Oni, O. Determination of optimum calcium and phosphorous ratio for the production of African catfish Clarias gariepinus (Burchell, 1822). J. Appl. Sci. Environ. Manag. 2018, 22, 689. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and metabolism of minerals in fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef]
- Sugiura, S.H.; Babbitt, J.K.; Dong, F.M.; Hardy, R.W. Utilization of fish and animal by-product meals in low-pollution feeds for rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2000, 31, 585–593. [Google Scholar] [CrossRef]
- Lazzari, R.; Baldisserotto, B. Nitrogen and phosphorus waste in fish farming. Bol. Do Inst. Pesca 2008, 34, 591–600. [Google Scholar]
- Munguti, J.M.; Kirimi, J.G.; Obiero, K.O.; Ogello, E.O.; Kyule, D.N.; Liti, D.M.; Musalia, L.M. Aqua-feed wastes: Impact on natural systems and practical mitigations—A review. J. Agric. Sci. 2021, 13, 111. [Google Scholar] [CrossRef]
- Teodósio, R.; Engrola, S.; Colen, R.; Masagounder, K.; Aragão, C. Optimizing diets to decrease environmental impact of Nile tilapia (Oreochromis niloticus) production. Aquac. Nutr. 2020, 26, 422–431. [Google Scholar] [CrossRef]
- Turcios, A.E.; Papenbrock, J. Sustainable treatment of aquaculture effluents—What can we learn from the past for the future? Sustainability 2014, 6, 836–856. [Google Scholar] [CrossRef] [Green Version]
- Graber, A.; Junge, R. Aquaponic systems: Nutrient recycling from fish wastewater by vegetable production. Desalination 2009, 246, 147–156. [Google Scholar] [CrossRef]
- Seawright, D.E.; Stickney, R.R.; Walker, R.B. Nutrient dynamics in integrated aquaculture-hydroponics systems. Aquaculture 1998, 160, 215–237. [Google Scholar] [CrossRef]
- Delaide, B.; Delhaye, G.; Dermience, M.; Gott, J.; Soyeurt, H.; Jijakli, M.H. Plant and fish production performance, nutrient mass balances, energy and water use of the PAFF Box, a small-scale aquaponic system. Aquac. Eng. 2017, 78, 130–139. [Google Scholar] [CrossRef]
- Suhl, J.; Dannehl, D.; Kloas, W.; Baganz, D.; Jobs, S.; Scheibe, G.; Schmidt, U. Advanced aquaponics: Evaluation of intensive tomato production in aquaponics vs. conventional hydroponics. Agric. Water Manag. 2016, 178, 335–344. [Google Scholar] [CrossRef]
- Villarroel, M.; Alvariño, J.M.R.; Duran, J.M. Aquaponics: Integrating fish feeding rates and ion waste production for strawberry hydroponics. Span. J. Agric. Res. 2011, 9, 537. [Google Scholar] [CrossRef]
- Robaina, L.; Pirhonen, J.; Mente, E.; Sánchez, J.; Goosen, N. Fish diets in aquaponics. In Aquaponics Food Production Systems; Springer: Cham, Switzerland, 2019; pp. 333–352. [Google Scholar] [CrossRef] [Green Version]
- Roy, K.; Kajgrova, L.; Mraz, J. TILAFeed: A bio-based inventory for circular nutrients management and achieving bioeconomy in future aquaponics. N. Biotechnol. 2022, 70, 9–18. [Google Scholar] [CrossRef]
- da Silva Cerozi, B.; Fitzsimmons, K. Effect of dietary phytase on phosphorus use efficiency and dynamics in aquaponics. Aquac. Int. 2017, 25, 1227–1238. [Google Scholar] [CrossRef]
- Stoyanova, S.; Sirakov, I.; Velichkova, K.; Ali, M. Effect of feed protein level on water chemical and technological parameters of a recirculating aquaponics system for carp (Cyprinus carpio L.) and lettuce (Lactuca sativa L.) farming. Turk. J. Fish. Aquat. Sci. 2019, 19, 885–891. [Google Scholar] [CrossRef]
- Strauch, S.M.; Wenzel, L.C.; Bischoff, A.; Dellwig, O.; Klein, J.; Schüch, A.; Wasenitz, B.; Palm, H.W. Commercial African catfish (Clarias gariepinus) recirculating aquaculture systems: Assessment of element and energy pathways with special focus on the phosphorus cycle. Sustainability 2018, 10, 1805. [Google Scholar] [CrossRef] [Green Version]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Deutsch, L.; Gra, S.; Folke, C.; Troell, M.; Huitric, M. Feeding aquaculture growth through globalization: Exploitation of marine ecosystems for fishmeal. Glob. Environ. Chang. 2007, 17, 238–249. [Google Scholar] [CrossRef]
- Marsh, L.; Subler, S.; Mishra, S.; Marini, M. Suitability of aquaculture effluent solids mixed with cardboard as a feedstock for vermicomposting. Bioresour. Technol. 2005, 96, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Monsees, H.; Keitel, J.; Paul, M.; Kloas, W.; Wuertz, S. Potential of aquacultural sludge treatment for aquaponics: Evaluation of nutrient mobilization under aerobic and anaerobic conditions. Aquac. Environ. Interact. 2017, 9, 9–18. [Google Scholar] [CrossRef]
- Goddek, S.; Delaide, B.P.L.; Joyce, A.; Wuertz, S.; Jijakli, M.H.; Gross, A.; Eding, E.H.; Bläser, I.; Reuter, M.; Keizer, L.C.P.; et al. Nutrient mineralization and organic matter reduction performance of RAS-based sludge in sequential UASB-EGSB reactors. Aquac. Eng. 2018, 83, 10–19. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Insects Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Shafi, M.E.; Alghamdi, W.Y.; Abdelnour, S.A.; Shehata, A.M.; Noreldin, A.E.; Ashour, E.A.; Swelum, A.A.; Al-sagan, A.A.; Alkhateeb, M.; et al. Black soldier fly (Hermetia illucens) meal as a promising feed ingredient for poultry: A comprehensive review. Agriculture 2020, 10, 339. [Google Scholar] [CrossRef]
- Schmitt, E.; Belghit, I.; Johansen, J.; Leushuis, R.; Lock, E.J.; Melsen, D.; Ramasamy Shanmugam, R.K.; Van Loon, J.; Paul, A. Growth and safety assessment of feed streams for black soldier fly larvae: A case study with aquaculture sludge. Animals 2019, 9, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Chen, L.; Yang, K.; Wang, T.; Wang, Y.; Jia, Y.; Yin, Y.; Gu, P.; Miao, H. Toxic effects of industrial flocculants addition on bioconversion of black soldier fly larvae (Hermetia illucens L.). Insects 2022, 13, 683. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient Composition (% Incorporation) | Experimental Diets | |||
|---|---|---|---|---|
| FM | BSF | PBM | PM | |
| Fish meal | 51.0 | - | - | - |
| Black soldier fly larvae meal | - | 61.6 | - | - |
| Poultry blood meal | - | - | 37.2 | - |
| Poultry meal | - | - | - | 56.4 |
| Wheat bran | 29.8 | 19.9 | 39.4 | 26.0 |
| Corn meal | 11.0 | 11.0 | 11.0 | 11.0 |
| Vitamin and mineral premix | 1.0 | 1.0 | 1.0 | 1.0 |
| Dicalcium phosphate (DCP) | 1.2 | 1.2 | 1.2 | 1.2 |
| Fish oil | 3.0 | 3.0 | 3.0 | 3.0 |
| Poultry fat | 3.0 | 2.3 | 7.2 | 1.4 |
| Ingredient cost (EUR/kg) | 1.11 | 1.99 | 0.72 | 0.72 |
| Proximate composition (%—as fed) 1 | ||||
| Dry matter (DM) | 91.90 ± 0.10 | 92.30 ± 0.10 | 91.60 ± 0.10 | 93.05 ± 0.15 |
| Crude protein (N × 6.25) (CP) | 40.30 ± 0.40 | 40.35 ± 0.05 | 40.90 ± 0.10 | 43.70 ± 0.10 |
| Crude fat (CF) | 11.55 ± 0.15 | 11.10 ± 0.20 | 11.60 ± 0.10 | 11.85 ± 0.25 |
| Crude fiber (CFB) | 2.85 ± 0.05 | 8.05 ± 0.15 | 4.20 ± 0.00 | 3.10 ± 0.00 |
| Ash | 11.65 ± 0.05 | 8.25 ± 0.05 | 4.35 ± 0.05 | 10.85 ± 0.05 |
| Starch | 12.05 ± 0.45 | 12.65 ± 0.25 | 13.45 ± 0.05 | 9.00 ± 1.00 |
| Nitrogen-free extract (NFE) 2 | 25.55 ± 0.05 | 24.55 ± 0.55 | 30.55 ± 0.35 | 23.55 ± 0.45 |
| Gross energy (GE) (MJ/kg) 3 | 18.48 ± 0.03 | 18.14 ± 0.00 | 19.49 ± 0.00 | 19.05 ± 0.05 |
| P/E ratio (g protein/MJ GE) | 21.81 ± 0.03 | 22.25 ± 0.03 | 20.98 ± 0.05 | 22.93 ± 0.00 |
| Essential amino acids (EAAs) (%—as fed) | ||||
| Arginine (Arg) | 2.48 | 1.88 | 2.32 | 3.07 |
| Histidine (His) | 1.24 | 1.18 | 2.07 | 0.99 |
| Isoleucine (Ile) | 1.79 | 1.59 | 1.71 | 1.60 |
| Leucine (Leu) | 3.20 | 2.52 | 4.24 | 2.96 |
| Lysine (Lys) | 3.11 | 1.91 | 3.22 | 2.51 |
| Methionine (Met) | 1.10 | 0.53 | 0.53 | 0.78 |
| Phenylalanine (Phe) | 1.71 | 1.43 | 2.43 | 1.67 |
| Threonine (Thr) | 1.75 | 1.47 | 1.96 | 1.63 |
| Tryptophan (Trp) | 0.51 | 0.53 | 0.77 | 0.37 |
| Valine (Val) | 1.86 | 1.89 | 2.29 | 1.71 |
| Met + Cys | 1.53 | 0.85 | 1.09 | 1.26 |
| Phe + Tyr | 2.92 | 3.06 | 3.42 | 2.85 |
| Minerals (g/kg—as fed) | ||||
| Ca | 26.71 | 14.22 | 5.72 | 27.60 |
| P | 19.12 | 12.10 | 7.83 | 18.64 |
| S | 4.99 | 2.98 | 3.43 | 4.34 |
| Mg | 2.21 | 3.48 | 1.41 | 1.76 |
| Fe | 0.31 | 0.16 | 0.85 | 0.13 |
| Na | 3.27 | 0.93 | 1.25 | 2.78 |
| K | 8.57 | 12.63 | 5.95 | 7.68 |
| Al | 0.25 | 0.04 | 0.01 | 0.02 |
| Zn | 0.089 | 0.151 | 0.055 | 0.092 |
| Mn | 0.047 | 0.242 | 0.050 | 0.046 |
| Cu | 0.015 | 0.021 | 0.014 | 0.018 |
| FM | BSF | PBM | PM | |
|---|---|---|---|---|
| O2 (mg/L) 1 | 7.50 ± 0.16 | 7.58 ± 0.19 | 7.55 ± 0.13 | 7.53 ± 0.14 |
| Temperature (°C) 1 | 26.6 ± 0.70 | 26.7 ± 0.88 | 26.8 ± 0.60 | 26.7 ± 0.63 |
| pH 1 | 7.88 ± 0.26 | 7.73 ± 0.47 | 7.90 ± 0.25 | 7.72 ± 0.44 |
| Conductivity (µS/cm) 1 | 980 ± 64.02 | 1014 ± 85.34 | 972 ± 59.11 | 1011 ± 83.45 |
| NH4+-N (mg/L) 2 | 0.28 ± 0.17 | 0.30 ± 0.16 | 0.26 ± 0.16 | 0.27 ± 0.14 |
| NO2−-N (mg/L) 2 | <0.05 | <0.05 | <0.05 | <0.05 |
| Fish Performance | ||||
|---|---|---|---|---|
| FM | BSF | PBM | PM | |
| Survival (%) A | 100.0 ± 0.0 a | 100.0 ± 0.0 a | 99.2 ± 1.7 a | 99.2 ± 1.7 a |
| Initial body weight (g) C | 11.64 ± 1.42 a | 11.52 ± 1.35 a | 11.21 ± 1.34 a | 11.14 ± 1.39 b |
| Final body weight (g) B | 38.02 ± 9.97 a | 24.77 ± 7.01 b | 22.17 ± 4.98 c | 34.82 ± 9.64 a |
| BWG (g) 1,A | 26.38 ± 0.27 a | 13.25 ± 0.08 c | 10.96 ± 0.15 d | 23.69 ± 0.86 b |
| Initial total length (cm) C | 11.96 ± 0.54 a | 11.98 ± 0.51 a | 11.85 ± 0.51 a | 11.85 ± 0.53 a |
| Final total length (cm) C | 18.23 ± 1.52 a | 15.77 ± 1.44 b | 15.40 ± 1.14 b | 17.74 ± 1.61 a |
| LG (cm) 2,A | 6.27 ± 0.10 a | 3.79 ± 0.08 c | 3.50 ± 0.07 d | 5.89 ± 0.14 b |
| Initial biomass (g) A | 349 ± 9 a | 345 ± 10 a | 336 ± 6 a | 334 ± 6 a |
| Final biomass (g) A | 1141 ± 9 a | 743 ± 7 c | 660 ± 8 d | 1036 ± 17 b |
| Feed administered (g—as is) | 578 | 578 | 578 | 578 |
| MDR (% biomass/d) 3,D | 1.74 ± 0.36 c | 2.20 ± 0.45 b | 2.39 ± 0.51 a | 1.84 ± 0.37 c |
| FCR (as fed) 4,A | 0.73 ± 0.01 d | 1.45 ± 0.01 b | 1.76 ± 0.02 a | 0.81 ± 0.03 c |
| FCR (DM basis) 4,A | 0.67 ± 0.01 d | 1.34 ± 0.01 b | 1.61 ± 0.02 a | 0.76 ± 0.03 c |
| SGR 5,A | 2.42 ± 0.05 a | 1.56 ± 0.04 c | 1.39 ± 0.01 d | 2.33 ± 0.03 b |
| CF 6,B | 0.62 ± 0.03 a | 0.62 ± 0.04 a | 0.60 ± 0.03 b | 0.61 ± 0.03 ab |
| PER 7,A | 3.40 ± 0.03 a | 1.71 ± 0.01 c | 1.39 ± 0.02 d | 2.81 ± 0.10 b |
| TGC 8,A | 0.84 ± 0.02 a | 0.50 ± 0.01 c | 0.43 ± 0.00 d | 0.79 ± 0.02 b |
| Feed ingredient cost (EUR/kg body weight gain) A | 0.81 ± 0.01 c | 2.89 ± 0.02 a | 1.27 ± 0.02 b | 0.59 ± 0.02 d |
| Proximate Composition (%) | Feces | |||
|---|---|---|---|---|
| FM | BSF | PBM | PM | |
| Crude protein (CP) | 14.50 ± 0.22 c | 28.10 ± 0.45 b | 51.43 ± 1.37 a | 14.85 ± 0.66 c |
| Crude fat (CF) | 3.05 ± 0.13 b | 4.95 ± 0.41 a | 2.08 ± 0.60 c | 3.70 ± 0.14 b |
| Crude fiber (CFB) | 15.18 ± 0.55 c | 26.63 ± 0.74 a | 12.03 ± 0.53 d | 17.33 ± 0.13 b |
| Ash | 21.78 ± 0.13 a | 9.68 ± 0.31 c | 5.25 ± 0.24 d | 14.65 ± 0.62 b |
| Starch | 1.98 ± 0.10 b | 3.40 ± 0.43 a | 1.58 ± 0.30 b | 2.28 ± 0.40 b |
| Nitrogen-free extract (NFE) | 45.50 ± 0.85 b | 30.65 ± 0.19 c | 29.23 ± 1.90 c | 49.48 ± 1.36 a |
| Gross energy (GE) (MJ/kg) | 12.44 ± 0.07 d | 13.86 ± 0.26 b | 17.99 ± 0.21 a | 13.46 ± 0.11 c |
| P/E ratio (g protein/MJ GE) | 11.66 ± 0.22 c | 20.28 ± 0.24 b | 28.59 ± 0.84 a | 11.03 ± 0.51 c |
| EAAs/NEAAs (%) | ||||
| Arginine (Arg) | 0.61 ± 0.02 b | 0.67 ± 0.02 b | 2.62 ± 0.09 a | 0.75 ± 0.09 b |
| Histidine (His) | 0.29 ± 0.02 c | 0.67 ± 0.03 b | 2.87 ± 0.11 a | 0.23 ± 0.01 d |
| Isoleucine (Ile) | 0.48 ± 0.02 d | 0.84 ± 0.05 b | 2.21 ± 0.09 a | 0.61 ± 0.03 c |
| Leucine (Leu) | 0.71 ± 0.03 d | 1.32 ± 0.07 a | 5.16 ± 1.60 a | 0.96 ± 0.04 c |
| Lysine (Lys) | 0.46 ± 0.04 c | 0.77 ± 0.04 b | 3.94 ± 0.10 a | 0.47 ± 0.02 c |
| Methionine (Met) | 0.33 ± 0.01 b | 0.29 ± 0.02 c | 0.63 ± 0.02 a | 0.21 ± 0.01 d |
| Phenylalanine (Phe) | 0.56 ± 0.01 c | 0.88 ± 0.05 b | 3.36 ± 0.14 a | 0.65 ± 0.04 c |
| Threonine (Thr) | 0.49 ± 0.02 d | 0.79 ± 0.03 b | 2.57 ± 0.08 a | 0.64 ± 0.03 c |
| Tryptophan (Trp) | 0.18 ± 0.01 c | 0.31 ± 0.01 b | 1.08 ± 0.05 a | 0.19 ± 0.01 c |
| Valine (Val) | 0.56 ± 0.02 d | 1.13 ± 0.05 b | 3.29 ± 0.20 a | 0.78 ± 0.06 c |
| Sum EAAs | 4.68 ± 0.18 c | 7.64 ± 0.34 b | 27.71 ± 2.00 a | 5.46 ± 0.32 bc |
| Alanine (Ala) | 0.79 ± 0.02 c | 1.36 ± 0.04 b | 4.18 ± 0.11 a | 0.86 ± 0.04 c |
| Cysteine (Cys) | 0.22 ± 0.01 c | 0.24 ± 0.01 c | 0.76 ± 0.01 a | 0.34 ± 0.03 b |
| Glycine (Gly) | 1.01 ± 0.03 d | 1.49 ± 0.04 b | 2.15 ± 0.05 a | 1.12 ± 0.07 c |
| Proline (Pro) | 0.61 ± 0.02 d | 0.97 ± 0.05 b | 2.15 ± 0.04 a | 0.80 ± 0.05 c |
| Serine (Ser) | 0.52 ± 0.02 d | 0.81 ± 0.06 b | 2.15 ± 0.07 a | 0.66 ± 0.05 c |
| Aspartic acid (Asp) + asparagine (Asn) | 1.05 ± 0.04 c | 1.39 ± 0.09 b | 4.51 ± 0.15 a | 1.15 ± 0.06 c |
| Glutamic acid (Glu) + glutamine (Gln) | 1.21 ± 0.06 c | 1.63 ± 0.08 b | 5.31 ± 0.12 a | 1.32 ± 0.09 c |
| Tyrosine (Tyr) | 0.43 ± 0.03 b | - | 1.05 ± 0.05 a | 0.53 ± 0.06 b |
| Sum NEAAs | 5.84 ± 0.17 d | 7.89 ± 0.35 b | 22.27 ± 0.56 a | 6.77 ± 0.42 c |
| Mineral composition (g/kg) | ||||
| Ca | 72.48 ± 6.77 a | 31.11 ± 0.45 c | 14.67 ± 0.14 d | 53.62 ± 2.10 b |
| P | 33.58 ± 2.51 a | 13.60 ± 0.26 c | 9.67 ± 0.15 d | 25.21 ± 0.57 b |
| S | 2.94 ± 0.09 c | 3.16 ± 0.05 b | 5.56 ± 0.13 a | 3.00 ± 0.03 bc |
| Mg | 2.74 ± 0.10 a | 1.69 ± 0.04 b | 1.21 ± 0.02 c | 1.63 ± 0.04 b |
| Fe | 1.12 ± 0.06 b | 0.61 ± 0.05 c | 2.19 ± 0.03 a | 0.51 ± 0.02 d |
| Na | 1.07 ± 0.07 a | 0.60 ± 0.01 c | 0.38 ± 0.01 d | 0.77 ± 0.02 b |
| K | 0.84 ± 0.01 a | 0.79 ± 0.02 b | 0.43 ± 0.01 d | 0.55 ± 0.01 c |
| Al | 0.81 ± 0.04 a | 0.18 ± 0.01 b | 0.04 ± 0.00 d | 0.10 ± 0.01 c |
| Zn | 0.44 ± 0.05 b | 0.57 ± 0.02 a | 0.20 ± 0.01 c | 0.44 ± 0.03 b |
| Mn | 0.14 ± 0.01 b | 0.45 ± 0.01 a | 0.07 ± 0.00 d | 0.11 ± 0.01 c |
| Cu | 0.07 ± 0.00 a | 0.07 ± 0.01 a | 0.04 ± 0.00 b | 0.07 ± 0.00 a |
| Nutrient Recovery | ||||
|---|---|---|---|---|
| FM | BSF | PBM | PM | |
| Feed administered (g—as is) | 578 | 578 | 578 | 578 |
| Feed administered (g DM) | 531 | 533 | 529 | 538 |
| Feces collected (g—as is) | 674 ± 13 c | 793 ± 30 b | 1063 ± 57 a | 664 ± 20 c |
| Feces collected (g DM) | 77.67 ± 1.17 b,c | 80.91 ± 2.48 b | 148.88 ± 8.15 a | 69.05 ± 3.29 c |
| (A) Recovery (% of the total amount of DM/the respective nutrient/energy fed throughout the trial) | ||||
| DM | 14.63 ± 0.22 b,c | 15.24 ± 0.47 b | 28.04 ± 1.54 a | 13.00 ± 0.62 c |
| Crude protein | 4.84 ± 0.13 c | 9.75 ± 0.37 b | 32.41 ± 2.19 a | 4.06 ± 0.15 d |
| Crude fat | 3.55 ± 0.14 b | 6.24 ± 0.54 a | 4.60 ± 1.31 a,b | 3.73 ± 0.15 b |
| Crude fiber | 71.57 ± 2.59 b | 46.30 ± 1.63 c | 73.72 ± 4.17 ab | 66.77 ± 2.79 b |
| Ash | 25.13 ± 0.40 b | 16.43 ± 0.91 c | 31.12 ± 2.50 a | 16.13 ± 0.86 c |
| Starch | 2.20 ± 0.10 b | 3.77 ± 0.53 a | 3.01 ± 0.53 ab | 3.02 ± 0.57 a,b |
| NFE | 23.94 ± 0.60 a | 17.48 ± 0.54 b | 24.64 ± 2.02 a | 25.12 ± 1.69 a |
| Gross energy (MJ/kg) | 9.05 ± 0.16 c | 10.70 ± 0.39 b | 23.78 ± 1.38 a | 8.44 ± 0.41 c |
| (B) Recovery per kg of feed (as is) (g/kg) | ||||
| DM | 134.43 ± 2.03 b,c | 140.03 ± 4.30 b | 257.66 ± 14.11 a | 119.50 ± 5.70 c |
| Crude protein | 19.49 ± 0.52 c | 39.35 ± 1.49 b | 132.55 ± 8.94 a | 17.73 ± 0.65 d |
| Crude fat | 4.10 ± 0.16 b | 6.93 ± 0.60 a | 5.34 ± 1.52 a,b | 4.42 ± 0.18 b |
| Crude fiber | 20.40 ± 0.74 c | 37.27 ± 1.31 a | 30.96 ± 1.75 b | 20.70 ± 0.86 c |
| Ash | 29.27 ± 0.47 a | 13.55 ± 0.75 c | 13.54 ± 1.09 c | 17.50 ± 0.93 b |
| Starch | 2.65 ± 0.13 b | 4.76 ± 0.67 a | 4.05 ± 0.71 a | 2.72 ± 0.51 b |
| NFE | 61.17 ± 1.52 b | 42.92 ± 1.32 c | 75.28 ± 6.17 a | 59.16 ± 3.98 b |
| Gross energy (MJ/kg) | 1.67 ± 0.03 c | 1.94 ± 0.07 b | 4.64 ± 0.27 a | 1.61 ± 0.08 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaw, C.; Knopf, K.; Kloas, W. Toward Feeds for Circular Multitrophic Food Production Systems: Holistically Evaluating Growth Performance and Nutrient Excretion of African Catfish Fed Fish Meal-Free Diets in Comparison to Nile Tilapia. Sustainability 2022, 14, 14252. https://doi.org/10.3390/su142114252
Shaw C, Knopf K, Kloas W. Toward Feeds for Circular Multitrophic Food Production Systems: Holistically Evaluating Growth Performance and Nutrient Excretion of African Catfish Fed Fish Meal-Free Diets in Comparison to Nile Tilapia. Sustainability. 2022; 14(21):14252. https://doi.org/10.3390/su142114252
Chicago/Turabian StyleShaw, Christopher, Klaus Knopf, and Werner Kloas. 2022. "Toward Feeds for Circular Multitrophic Food Production Systems: Holistically Evaluating Growth Performance and Nutrient Excretion of African Catfish Fed Fish Meal-Free Diets in Comparison to Nile Tilapia" Sustainability 14, no. 21: 14252. https://doi.org/10.3390/su142114252
APA StyleShaw, C., Knopf, K., & Kloas, W. (2022). Toward Feeds for Circular Multitrophic Food Production Systems: Holistically Evaluating Growth Performance and Nutrient Excretion of African Catfish Fed Fish Meal-Free Diets in Comparison to Nile Tilapia. Sustainability, 14(21), 14252. https://doi.org/10.3390/su142114252