Abstract
Babesiosis is a significant tick-borne disease that causes varying degrees of losses to animals and humans, as well as a severe economic impact. In Nigeria, there have been several reports on the prevalence of Babesia infection in sheep; however, to date, there is no documented report on the molecular characterization of Babesia ovis in sheep. Here, we determined the prevalence of Babesia infection in sheep using microscopy and PCR and further characterized Babesia ovis in sheep in Nigeria. In this study, 198 blood samples were collected from Abuja and Abeokuta, Nigeria. Microscopic and polymerase chain reactions were used to detect the presence of B. ovis in sheep. Genomic DNA was extracted from blood samples, and generic RLB forward and reverse primers were used to amplify the 18S rRNA segment of B. ovis. Sequence analysis of the generic molecular marker was used to determine the genetic characteristics of B. ovis in sheep in Nigeria. The prevalence of B. ovis infection using microscopy and PCR was 61.1% and 36.9%, respectively. There was a higher prevalence of Babesia infection in Abeokuta (38.4%) than in Abuja. Study animals of one and half years to three years had the highest percentage (45.8%) of Babesia infection, and higher infection of B. ovis was recorded in male animals (37.5%), balami breeds (40.0%), white coat colors (44.4%), emaciated animals (75.0%), and polycythaemic animals (57.1%). Sequencing analysis indicated that B. ovis 18S rRNA from southwestern Abeokuta and north-central Abuja, Nigeria, showed 90–95% identity of documented isolates from other countries. BioEdit and MEGAX software was used to clean sequences and construct a phylogenetic tree to show evolutionary relationships. In conclusion, the findings from this study offer significant information on the molecular characteristics of B. ovis infection for the first time in Nigeria, as well as its present prevalence status. Furthermore, sheep have been identified as a potential reservoir for this tick-borne pathogen; thus, the information from this study can serve as a basis to formulate effective control strategies for tick-borne pathogens circulating amongst the ruminant population in Nigeria and Africa by extension.
1. Introduction
Hemoparasites have great economic impacts on ruminant production and are responsible for over 3 billion USD in losses in the livestock industry. Sheep production bears approximately 60% of these losses [1]. Infections with hemoparasites pose a significant danger to food security, particularly in livestock-dependent populations in Sub-Saharan Africa [2]. The presence and significance of hemoparasites are a result of intricate interactions between pathogens that cause illness, their vectors, the vertebrate host, and the environment [3]. Babesia is a major hemoparasite affecting different animals worldwide.
The Babesia parasite is an intra-erythrocytic organism of the phylum Apicomplexa [4]. It manifests a variety of clinical signs in animals [5]; some of these signs include fever, splenomegaly, anemia, jaundice, hemoglobinuria, anorexia, weakness, dehydration, and even death [6,7,8]. Babesia is transmitted to susceptible hosts via tick bites [9].
Tick-borne diseases (TBDs) are widespread worldwide, although they are more common in tropical and subtropical regions [10]. Endemic tick and tick-borne diseases have caused an annual loss of approximately 13.6 billion USD [11]. Huruma et al. [12] showed that different ticks had various preferred sites on the host’s body. Additionally, they decrease milk production and bodyweight gain while also opening up pathways for secondary pathogen invasion [13]. Ticks transmit more harmful microorganisms than any other arthropod vector group and are one of the most significant vectors of animal diseases [14]. There has been evidence linking some Rhipicephalus species, notably R. bursa, R. sanguineus, and R. turanicus, to the spread of Babesia spp. in sheep. According to reports, R. bursa is the sole vector capable of transmitting B. ovis. Despite this knowledge, there is a scarcity of information regarding the epidemiology and enzootic potential of this parasite [15].
B. ovis cause Babesiosis in sheep; it is also called Piroplasmosis or Red-water fever [16]. Pathogenic agents for sheep and goats are recognized to be B. ovis and B. motasi, respectively [17]. Hard ticks, which transmit Babesia species, allow the Babesia organism to spread trans-ovarially from one tick generation to the next via the egg [18,19]. Generally, Babesiosis occurs worldwide, but the geographic and seasonal distribution of the vectors determines where the pathogenic protozoa are found [17,20]. Both B. ovis and B. motasi infections can affect sheep; however, B. ovis is more threatening to sheep than to goats [21].
In Nigeria, there had been reported cases of Babesia infection in sheep, with varying prevalence rates of the causative agent. In Abeokuta (southwestern part of Nigeria), a study was carried out on ‘Haemoprotozoan parasites of sheep’ by Takeet et al. [22], in which B. ovis had a prevalence of 12%. In Ibadan, Oyo State Adejinmi et al. [23] reported a 1.9% prevalence. Jatau et al. [24], Samaila and Musa [25], Opara et al. [26], and Egbe-Nwiyi et al. [27] reported Babesia spp. prevalence rates of 0.97%, 1%, 3.1%, and 1%, in Kano, Bauchi, Nasarawa, and Borno in the northern part of the country, respectively. All these reports utilized microscopic techniques for the detection and identification of the Babesia parasite in the studied animals; however, the sensitivity of microscopic or morphological identification of Babesia species has been queried [28].
It should be noted, however, that none of the published studies on B. ovis of sheep in Nigeria have used molecular methods to describe the organism. This study used microscopy and PCR for the detection and characterization of B. ovis infection in relation to age, sex, coat color, breed, packed cell volume evaluation, body condition score, and location in Nigerian sheep.
2. Materials and Methods
2.1. Study Area
Samples were collected from one abattoir (Karu) in Abuja, Federal Capital Territory (the largest commercial abattoir in the state), and eight individual farms reared on a semi-intensive management system in Abeokuta, Ogun state, Nigeria. The samples were collected between February and April 2021, which was a late dry season entering the wet season in Nigeria.
Nigeria has a tropical climate. Abuja is located in the North-central part of the country (9°4′ N, 7°29′ E), 477 m above sea level with an average annual temperature of 25.7 °C and 1389 mm of precipitation annually. Abeokuta, Ogun State is located in the southwestern part of the country (7°9′39″ N, 3°20′54″ E). Abeokuta lies 80 m above sea level with an average temperature of 27.1 °C and average precipitation of 1238 mm.
2.2. Study Population
In this study, sheep of different breeds (WAD, Balami, Yankasa and Uda) were randomly sampled. The age of the animals was determined using their dentition. Data on breed, sex, coat color, body condition, and weight were also recorded. Using a visual examination, the body condition score was classified into one of five groups (emaciated, thin, average, fat, or obese) based on fat accumulation at the back and flank area. Young lambs that were less than 6 weeks, pregnant, and recently lambed sheep were excluded from sampling for safety reasons.
2.3. Sample Collection
Blood samples were collected from 198 sheep in Abeokuta and Abuja using a 21-gauge hypodermic needle in a 5 mL syringe from the jugular vein of each sheep. Approximately 5 mL of whole blood was aseptically collected from each sheep into an EDTA bottle and mixed gently to enable an even spread of anticoagulant with the blood. The sample was transported to the laboratory on ice for further analysis.
2.4. Thin Blood Smear Technique
The thin blood smear technique was carried out in the Veterinary Parasitology and Entomology Laboratory, College of Veterinary medicine, Federal University of Agriculture, Abeokuta, following standard procedures [29]. The smears were stained for 20 s each using a field stain containing methylene blue and Eosin in jars A and B, respectively. The slides were allowed to air dry and viewed with a Light microscope under 100× magnification (oil immersion).
2.5. Packed Cell Volume Determination
A microhematocrit centrifuge was used to assess the packed cell volume (PCV) as described by [30]. It was determined by centrifuging EDTA-treated blood in a capillary tube (also known as a microhematocrit tube) at 10,000 rpm for five minutes. This separates the blood into layers of plasma, buffy coat, and red cells. The PCV reader was used to determine the percentage of packed cell volume.
2.6. DNA Extraction of Blood Samples
The DNA of blood samples was extracted using the ZYMO miniprep kit (INQUABA BIOTECH) according to the manufacturer’s instructions. The purity and concentration of DNA were assessed using the Nanodrop, 2000. Samples were stored at −20 °C until use.
2.7. Primer Set Determination
After genomic DNA samples were extracted, a primer for the ribosomal RNA gene that was used to amplify a portion of the rRNA gene of the B. ovis organism was retrieved from the literature [31]. A small sub-unit rRNA gene of B. ovis was amplified with a pair of Oligo-Nucleotide primers, with the primers RLB-F-Forward primer sequence (5′–3′); GACACAGGGAGGTAGTGACAAG and RLB-R-Reverse primer sequence (5′–3′); CTAAGAATTTCACCTCTGACAGT with an expected fragment length of 480 base pairs (bp) [31].
2.8. PCR Conditions
To confirm B. ovis presence, a 50 uL volume of the PCR reaction mixture was used containing the Dreamtaq Green PCR Master Mix (DreamTaq DNA polymerase, 2× Dreamtaq Green buffer, dATP, dCTP, dGTP, and dTTP, 0.4 mM each, and 4 mM MgCl2) and 1 uM each of forward and reverse primers, the template DNA sample, and distilled water. The PCR conditions were set at initial denaturing at 95 °C for 3 min, with 35 cycles of 95 °C for 3 min, 50 °C for 1 min 30 s, and 72 °C for 1 min, with a final extension of 72 °C for 5 min.
2.9. Agarose Gel Electrophoresis
B. ovis PCR amplicons were analyzed in 1.5% agarose gel electrophoresis stained with 10 mg/mL of ethidium bromide. Furthermore, a 100 bp ladder was used as a guide to determine the various amplicon sizes. Gel images were viewed under UV light using a Chemi-Doc system from Bio-Rad (Bio-Rad Laboratories, Hercules, CA, USA).
2.9.1. Sequencing
Positive results from PCR were sequenced uni-directionally at the Inquaba biotech sequencing facility, South Africa, using Sanger sequencing, and were subjected to analysis using MEGAX and BioEdit software.
The UPGMA algorithm was used to infer the phylogenetic relationships [32]. The evolutionary history of the species under study is predicted to be represented by the bootstrap consensus tree generated from 1000 replicates [33]. The Tajima–Nei approach was used to evaluate evolutionary distances [34]. Evolutionary analyses were conducted in MEGA X [35]. Trypanosoma brucei and Toxoplasma gondii were used as the outgroup of the phylogeny (Figure 1).
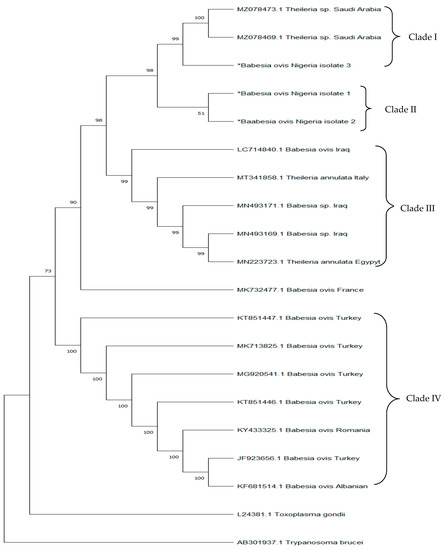
Figure 1.
The evolutionary relationships of different isolates from the Genbank with isolates from this study. ‘∗’ indicates ‘Isolates of B. ovis confirmed from this study’.
2.9.2. Nucleotide Sequence
The nucleotide sequence data reported in this study have been deposited in the GenbankTM database under accession numbers ON307466. ON307467, and ON307468 as representatives of B. ovis in this study.
2.9.3. Statistical Data Analysis
Phenotypic data were summarized using descriptive statistics and analyzed using Epi Info Software 7.0. The prevalence of Babesia spp. obtained by PCR in relation to sex, breeds, age, coat color, body condition score, PCV evaluation, and location were compared using the chi-square test. Significant differences were declared at p < 0.05. The confidence intervals of all variables were declared at 95%.
3. Results
3.1. Demographics Distribution of Different Variables of Sheep from This Study
Table 1 highlights the frequency distribution of different variables from this study. West African dwarf animals (57.6%), females (63.6%), animals more than 1.5 years and up to 3 years (41.9%), anemic animals (51.5%), white–black coat color animals (73.2%), and animals categorized as average based on their body condition score (55.1%) all had higher frequencies of samples when equal numbers of animals (50%) were sampled based on location.

Table 1.
Frequency distribution of the various variables from this study.
3.2. Prevalence of Babesia Ovis Infection in Sheep Detected by Microscopy and PCR
The overall prevalence of Babesia infection in sheep using the microscopic and PCR diagnostic methods was 61.1% (121/198) and 36.9% (73/198), respectively, with Abeokuta and Abuja having a prevalence of 67.7% (67/99) and 54.6% (54/99) for microscopy and 38.4% (38/99) and 35.4% (35/99) for PCR, respectively (Table 2).
Table 2.
Prevalence of Babesia ovis infection relative to different variables using PCR diagnostic method.
The highest infection rate of 45.8% was found in sheep between 1.5 and 3 years of age while those that were 1.5 years and below had the lowest rate (27.1%). There was no significant difference (p = 0.123) between the age groups of sheep and their infection with Babesia.
Sexes of sheep in relation to Babesia infection showed that males had a higher infection rate (37.5%) than females (36.5%), and there was no significant difference (p = 0.889) between the sexes of the animals and Babesia infection. Babesia infection as it relates to different breeds of sheep was also observed in this study. The Balami breed had the highest infection rate of B. ovis (40.0%), while the Uda breed had the lowest rate (31.0%) with no significant difference (p = 0.792).
Amongst the animals sampled, animals with a white coat color had a higher infection rate (44.4%) while the brown and white-colored animals had the lowest (30.8%), with no statistically significant difference (p = 0.831). Polycythemic animals were found to have a higher infection rate with B. ovis (57.1%) while animals that were anemic had the lowest infection rate (35.3%). There was, however, no significant difference (p = 0.510) observed amongst the various categories of PCV evaluation in relation to infection.
The body condition score evaluation of animals in relation to infected animals was also considered in this study. Emaciated animals had the highest infection rate (75.0%) of Babesia ovis while the average animals had the lowest rate (33.0%). There was no significant difference (p = 0.180) observed (Table 2).
3.3. Sequence Similarities and Phylogenetic Analysis
Three isolates were used as representative isolates of B. ovis for sequence analysis. The partial sequence of the 18S rRNA gene fragment of the isolates shared a similarity of 99.55% to 99.78% amongst themselves and 100% similarities with isolates from Saudi Arabia (MZ078472 and MZ078473). Our isolates also shared 91–95.55% similarity with the isolates from Iraq (LC714840), France (MK73247), Turkey (KT851447), Albanian (KF681514), Romania (KY433325), Italy (MT34185), and Egypt (MN223723). The phylogenetic analysis revealed four clades. Nigeria isolates formed clade II. However, one of our isolates formed a monophyletic relationship with clade I from Saudi Arabia (Figure 1).
4. Discussion
Microscopy methods have been the major technique to determine the prevalence of B. ovis infection in Nigeria [22,26]. However, studies on the high sensitivity and specificity favored the PCR technique over microscopy [31,36]. In this study, we utilized both microscopy and PCR methods in determining and comparing the prevalence of Babesia infection in Nigerian sheep. Microscopy and PCR were used to investigate the prevalence of B. ovis infection, while the PCR technique was used to determine the molecular characteristics of B. ovis infection of sheep in Abeokuta, Southwestern Nigeria, and Abuja, North Central Nigeria, in order to better understand the Babesia species circulating in the ovine population.
The greater prevalence of Babesia infection (61.1%) detected using microscopy compared to the molecular technique (36.9%) was consistent with Naderi et al. [37], who also found a higher prevalence when using microscopy. This might be due to a variety of factors, including artifacts, particularly when stains are stale, glass slides are greasy or dirty, or when the lenses of light microscopes are old, dirty, or faulty. Young, inexperienced scientists tend to miss the parasite if they are not careful, and time, focus, and patience are necessary attributes to imbibe when identifying parasites. Misdiagnosis is a common error, as several parasites might be confused for B. ovis, especially Theileria spp. and Anaplasma spp. as they are both intra-erythrocytic. All of these factors are commonly linked to microscopic detection [38]. The overall prevalence of B. ovis infection using the PCR method was 36.9%, which closely agrees with Shahzad et al. [35], who recorded a 34% prevalence of B. ovis infection using PCR, and a higher prevalence of 50% was also reported by Iqbal et al. [39], both of which were studies conducted in Pakistan. However, our report contrasts previously recorded prevalences of 0%, 8.25%, 16.27%, and 17.4% reported by Onyeche et al. [40] in Nigeria, Aktas et al. [41] and Altay et al. [42] in Turkey, and Rjeibi et al. [43] in Tunisia, respectively. It is possible that the cause for such disparities in prevalence is related to variances in geographical areas, different primer sets, and the climatic conditions in these regions. There has been evidence of cytoadherence and sequestration in Babesia bovis infection [44], and while these factors could influence the infection status of the animal via the blood collection sites, there has not been any reported evidence of such occurrences with B. ovis infection. Thus, further in vitro and in vivo studies on these factors (cytoadherence and sequestration) will be helpful.
In the present study, we observed that sheep aged more than 1.5 years and up to 3 years exhibited a higher degree (45.8%) of infection than any other age group. This is consistent with an earlier report by Rjeibi et al. [43] but disagrees with the report of Iqbal et al. [39], which states that sheep younger than one year old had more infections. The reason for such a high prevalence among this age group (>1.5–3 years) of animals from this study could be attributed to increased exposure to free-range grazing of sheep around farmlands, bushes, and so on, having gained maturity and thought to be at high productivity at such an age range, thus increasing the risk of exposure to the vector causing B. ovis infection. The lower prevalence (27.1%) observed among animals less than 1.5 years might be related to a more intense management approach applied to such animals due to their sensitivity, fragility, and potential for future productivity.
Male animals exhibited a greater degree of infection (37.5%) than female animals (36.5%), which fits with studies from Iqbal et al. [39], who found more B. ovis infection in males, but differs from reports by Rjeibi et al. [43] who found more B. ovis infection in females. This might simply be due to the number of samples collected for this study.
No significant difference was observed among breeds of sheep sampled relative to Babesia infection. However, the Balami breed had a higher infection rate (40.0%) than any other breed. This is in agreement with the reports of Adua et al. [45], who recorded a high prevalence (71.43%) of B. ovis infection in the Balami breed of sheep. The low sample size from our study could be a contributing factor to this relatively high infection rate; however, in Nigeria; there is also a scarcity of data on the effect of breed on the infection status (B. ovis) of sheep.
Sheep with different coat colors were also observed in relation to their infection status. Those with white coat colors were seen to have a higher degree (44.4%) of infection than any other category of coat colors. The reason for this could be the fact that white colors tend to reflect heat better than darker colors, and ticks, which are the vectors involved in the transmission of B. ovis infection, tend to thrive in a more conducive environment (less heat) which facilitates breeding and multiplying [46]. Therefore, white coat colors tend to maintain optimal temperatures for the replication of vectors. However, there has been no information on the effect of coat color relative to Babesia infection of sheep in Nigeria.
The body score evaluation had no significant difference regarding the B. ovis infection; however, emaciated animals had the highest degree (75.0%) of infection compared to others, which may be due to the pathogenesis of how B. ovis cause its deleterious effect on animals causing anorexia, progressive loss of weight, and emaciation. This is in agreement with the reports of Bello et al. [47] who reported a 16.4% prevalence of hemoparasitic infection in poor-condition sheep. Ngole et al. [48] and Masiga et al. [49] also reported poor feeding and emaciation as a consequence of hemoparasitic infection.
There was no significant difference observed between the infection status of sheep with Babesia and the two locations; however, Abeokuta had a higher prevalence (38.4%) of infection than Abuja (35.4%), which could be due to the sample size of sheep in the different locations of this study.
The packed cell volume (PCV) was categorized in this study using a benchmark of 27–45%, as <27% has been regarded as anemia and >45% is regarded as polycythemia due to increased dehydration. From this study, it was noted that polycythemic animals had a higher rate of infection than any other category of animals, which could be attributed to the sample size of the present study. This is in agreement with the report of Ademola and Onyiche [50] who found no significant difference between the PCV values and the infection status but in disagreement with the reports of Adejinmi et al. [23] and Bello et al. [47]. Babesia parasites are constantly under threat by the host’s immune system, thereby causing an upshot of genetic variants [51]. Several studies have reported the genetic polymorphism of Babesia spp. with the 18S rRNA gene [52,53,54].
In the phylogenetic tree, B. ovis Nigeria isolate 3 from this study formed a well-supported clade with Theileria spp. isolates from Saudi Arabia, showing some degree of diversity. However, B. ovis Nigeria isolates 1 and 2 formed a sub-cluster (Clade II) suggesting that isolates in Clade II are slightly divergent from isolates in Clade I even though they share a common ancestral origin. The introduction of this strain could be through the importation of sheep and/or vectors carrying the parasites into the country; further studies investigating strain diversity and vector distribution amongst ruminant animals in Nigeria would shed more light on this. Isolates from Iraq, Italy, and Egypt all formed a well-supported clade (Clade III), suggesting that the Nigerian isolates are more divergent from the Northern African type and are distantly related to isolates from clades I and II. This divergence could be attributed to evolutionary changes that promote environmental adaptation for survival in varied climatic environments. A re-sequencing of African strains would better enhance our understanding of the evolutionary footprint responsible for this divergence. Furthermore, isolates from Turkey, Albania, and Romania also formed a well-supported clade (Clade IV) and are more distantly related to Clades I, II, and III. This further suggests that isolates from this study are clearly distinct from isolates found in other regions and indicative of several strains being circulated amongst ruminant animals in Africa and Asia.
Limitation
The limitation of this study can be attributed to the relatively low number of samples used, and this may have been the reason for not having statistically significant values amongst the different variables. This may form a basis for further studies to significantly increase their sample size amongst the study population to better reflect findings in a named population.
5. Conclusions
This study reveals the endemic occurrence of B. ovis in sheep breeds of different age groups, sexes, and coat colors. This is Nigeria’s first research conducted to molecularly detect and characterize B. ovis in sheep. The current findings suggest two distinct strains of B. ovis are circulating in the different sheep breeds in Nigeria. Our findings described the prevalence of the parasite in sheep of different sexes, ages, and coat colors. The findings of the study suggest that although the prevalence of B. ovis varies in sheep populations, the prevalence is not significantly affected by sex, age, coat color, blood parameters, or body score condition. However, the higher the prevalence, the less fit the animals. Our findings also showed that sheep are key players in harboring and distributing tick-borne pathogens. As the role of hygiene in sheep production was not a part of this study, it is therefore difficult to clarify whether animals in which a lower prevalence was recorded enjoyed better hygiene or not. However, this could form the basis for future studies. Further studies on the genetics of circulating strains of B. ovis and their diversity are recommended for a comprehensive picture at the national level for policy formulation and management in curbing the spread and dissemination of B. ovis infection amongst the ruminant population in Nigeria.
6. Recommendation
Further research into the tick species implicated in the transmission of Babesia ovis in sheep should be conducted. The genetic diversity within species should also be researched in order to better comprehend the organism’s polymorphic traits. The government should provide the appropriate information to farmers about the economic relevance of B. ovis in sheep and its public health significance, as this assists in providing the blueprint for reaching the pre-elimination stage of this disease in Nigeria.
Author Contributions
Conceptualization, T.S.A., M.I.T. and M.O.; methodology, T.S.A.; soft-ware, T.S.A. and M.O.; validation, M.O., M.I.T., F.A.A. and A.O.S.; formal analysis, T.S.A., M.O. and M.I.T.; investigation, T.S.A., M.I.T., M.O., F.A.A. and A.O.S.; resources, T.S.A. and M.O.; data curation, T.S.A., M.O. and M.I.T.; writing—original draft preparation, T.S.A.; writing—review and editing, T.S.A., M.O. and M.I.T.; visualization, M.O.; supervision, M.O., M.I.T., F.A.A. and A.O.S.; project administration, M.O. and M.I.T.; funding acquisition, T.S.A., M.I.T. and A.O.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Tertiary Education Trust Fund, Federal Republic of Nigeria, grant number TETFUND 2021.
Institutional Review Board Statement
This study was approved by the Institutional Review Board (IRB) of the College of Veterinary Medicine, Federal University of Agriculture, Abeokuta, and the protocols used in this study were approved by the Ethical Committee on Research, College of Veterinary Medicine, Federal University of Agriculture, Abeokuta with approval number FUNAAB/COLVET/CREC/2021/02/01.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, M.O., upon reasonable request. Nucleotide sequences from this study have been deposited into the GenbankTM database repository.
Acknowledgments
The authors want to acknowledge the support of the staff of the Department of Veterinary Parasitology and Entomology, College of Veterinary Medicine, Federal University of Agriculture Abeokuta, and Discipline of Genetics (Genomics and Bioinformatics Group, Westville Campus), School of Life Sciences, University of Kwazulu-Natal, South Africa.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Iangopathy, M.; Palavesam, A.; Amaresan, S.; Muthusamy, R. Economic Impact of Gastrointestinal Nematodes in Sheep on Meat Production. Int. J. Livest. Res. 2019, 9, 44–48. [Google Scholar]
- Kasozik, I.; Matovu, E.; Tayebwa, D.S.; Natuhwera, J.; Mugezi, I.; Muhero, M. Epidemiology of increasing hemoparasite burden in Ugandan cattle. Open J. Vetrinary Med. 2014, 4, 220–231. [Google Scholar] [CrossRef]
- Akande, F.A.; Takeet, M.I.; Makanju, O. Haemoparasites of cattle in Abeokuta, South-west, Nigeria. Sci. World J. 2010, 5, 19–21. [Google Scholar]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B. Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 9102–9114. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, J.S.; Luo, J.; Schnittger, L.; Seitzer, U.; Jongejan, F.; Yin, H. Phylogenetic position of small ruminant infecting piroplasms. Ann. N. Y. Acad. Sci. 2006, 1081, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Sevinc, F.; Sevinc, M.; Ekici, O.D.; Yıldız, R.; Isık, N.; Aydogdu, U. Babesia ovis infections: Detailed clinical and laboratory observations in the pre- and post-treatment periods of 97 field cases. Vet. Parasitol. 2013, 191, 35–43. [Google Scholar] [CrossRef]
- Sevinc, F.; Xuan, X. Major tick-borne parasitic diseases of animals: A frame of references in Turkey. Eurasian J. Vet. Sci. 2015, 31, 132–142. [Google Scholar] [CrossRef]
- Zahid, I.A.; Latif, M.; Baloch, K.B. Incidence and treatment of theileriasis and Babesiosis. Pak. Vet. J. 2005, 25, 137–140. [Google Scholar]
- Walker, A.R.; Bouattour, A.; Camicas, J.L.; Estrada-Peria, A.; Horak, I.; Latif, A.; Pegram, R.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003. [Google Scholar]
- Lopes, L.B.; Nicolino, R.; Capanema, R.O.; Oliveira, C.S.F.; Haddad, J.P.A.; Eckstein, C. Economic impacts of parasitic diseases in cattle. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Res. 2015, 10, 1–10. [Google Scholar] [CrossRef]
- Huruma, G.; Abdurhaman, M.; Gebre, S.; Deresa, B. Identification of tick species and their prevalence in and around Sebeta town, Ethiopia. J. Parasitol. Vector Biol. 2015, 7, 1–8. [Google Scholar]
- Marufu, M.C. Prevalence of Ticks and Tick-borne Diseases in Cattle on Communal Rangelands in the Highland Areas of the Eastern Cape Province, South Africa. Master’s Thesis, Department of Livestock and Pasture Science Faculty of Science and Agriculture, University of Fort Hare, Alice, South Africa, 2008; pp. 1–134. [Google Scholar]
- Yiwombe, K. An Investigation to Determine the Resistance of the Boophilus Tick (Blue Tick) to Amitraz in Selected Areas of Zimbabwe. Bachelor’s Thesis, Midlands State University, Gweru, Zimbabwe, 2013. [Google Scholar]
- Esmaeilnejad, B.; Tavassoli, M.; Asri-Rezaei, S.; Dalir-Naghadeh, B.; Mardani, K.; Jalilzadeh-Amin, G.; Golabi, M.; Arjmand, J. PCR-Based Detection of Babesia ovis in Rhipicephalus bursa and Small Ruminants. J. Parasitol. Res. 2014, 2014, 294704. [Google Scholar] [CrossRef] [PubMed]
- Onoja, I.I.; Malachy, P.; Mshelia, W.P.; Okaiyeto, S.O.; Danbirni, S.; Kwananshie, G. Prevalence of Babesiosisin cattle and goats at Zaria abbatoir. Niger. J. Vet. Adv. 2013, 3, 211–214. [Google Scholar]
- Fakhar, M.; Hajihasani, A.; Masoufi, S.; Alizadeh, H.; Shirzad, H.; Piri, F.; Pagheh, A.S. An epidemiological survey on bovjne and ovine Babesiosis in Kurdistan province, Western Iran. Trop. Anim. Health Prod. 2012, 44, 319–322. [Google Scholar] [CrossRef]
- Ijaz, M.; Rehman, A.; Ali, M.M.; Umar, M.; Khalid, S.; Mehmoo, K.; Hanif, A. Clinico-epidemiology and therapeutical trials on Babesiosis in sheep and goats in Lahore, Pakistan. J. Anim. Plant Sci. 2013, 23, 666–669. [Google Scholar]
- Urquhart, G.M.; Armour, J.; Duncan, J.L.; Dunn, A.M.; Jennings, F.W. Veterinary Parasitology, 2nd ed.; Blackwell Science Incorporated: Hoboken, NJ, USA, 1996; pp. 242–253. [Google Scholar]
- Radostits, O.M.; Gay, G.C.; Hinchiff, K.W.; Constable, P.O. Veterinary Medicine: A Text Book of the Diseases of Cattle, Sheep, Goat, Pigs and Horses, 10th ed.; Saunders Elsevier: London, UK, 2007; pp. 1110–1489, 1527–1530. [Google Scholar]
- Rahbari, S.; Nabian, S.; Khaki, Z.; Alidadi, N.; Ashrafihelan, J. Clinical hematologic and pathologic aspects of experimental ovine Babesiosis in Iran. Iran. J. Vet. Res. 2008, 9, 58–64. [Google Scholar]
- Takeet, M.I.; Akande, F.A.; Abakpa, S.A.V. Haemoprotozoan parasites of sheep in Abeokuta, Nigeria. Niger. J. Parasitol. 2009, 30, 142–146. [Google Scholar]
- Adejinmi, J.O.; Sadiq, N.A.; Fashanu, S.O.; Lasisi, O.T.; Ekundayo, S. Studies on the blood parasites of sheep in Ibadan, Nigeria. Afr. J. Biomed. Res. 2004, 7, 41–43. [Google Scholar] [CrossRef]
- Jatau, I.D.; Abdulganiyu, A.; Lawal, A.I.; Okubanjo, O.O.; Yusuf, K.H. Gastrointestinal and Haemoparasitism of sheep and goats at slaughtered Abattoir in Kano, Northern Nigeria. Sahel J. Vet. Sci. 2011, 9, 7–11. [Google Scholar]
- Samaila, A.B.; Musa, B.L. Prevalence of Haemoparasite of sheep and goats slaughtered in Bauchi Abattoir. Int. J. Appl. Biol. Res. 2012, 4, 128–133. [Google Scholar]
- Opara, M.N.; Santali, A.; Mohammed, B.R.; Jegede, O.C. Prevalence of Haemoparasites of small Ruminants in Lafia Nassarawa State. A Guinea Savannah zone in Nigeria. J. Vet. Adv. 2016, 6, 1251–1257. [Google Scholar]
- Egbe-Nwiyi, T.N.; Sheriff, G.A.; Paul, B.T. Prevalence of tick-borne haemoparasiticdiseases (TBHDS) and Hematological changes in sheep and goats in Maiduguri Abattoir. J. Vet. Med. Anim. Health 2018, 10, 28–33. [Google Scholar] [CrossRef]
- Takeet, M.I.; Oyewusi, A.J.; Abakpa, S.A.V.; Daramola, O.D.; Peters, S.O. Genetic diversity among Babesia rossi detected in naturally infected dog in Abeokuta, Nigeria, based 18S rRNA gene sequences. Acta Parasitol. 2017, 62, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.M.G.; Paul, J.; Zaman, V. Medical and Veterinary Protozoology, an Illustrated Guide, Revised ed.; Churchill Livingstone: Edinburgh, UK; London, UK, 1977; pp. 32–49. [Google Scholar]
- Cheesbrough, M. Destrict Laboratory Practice in Tropical Countries; Cambridge University Press: Cambridge, UK, 2005; pp. 64–67. [Google Scholar]
- Nagore, D.; García-Sanmartín, J.; García-Pírez, A.L.; Juste, R.A. Hurtado Identification, genetic diversity and prevalence of Theileria and Babesia species in a sheep population from Nortern Spain. Int. J. Parasitol. 2004, 34, 1059–1067. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy; W. H. Freeman & Co.: New York, NY, USA, 1973. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Tajima, F.; Nei, M. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1984, 1, 269–285. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Shahzad, W.; Noor, H.; Ahmad, M.U.; Munir, R.; Saghar, M.S.; Hassan, M.M.; Ahmad, N.; Akbar, G.; Mehmood, F. Prevalence and Molecular Diagnosis of Babesia ovis and Theileria ovis in Lohi Sheep at Livestock Experiment Station (LES), Bahadurnagar, Okara, Pakistan. Iran. J. Parasitol. 2013, 8, 570–578. [Google Scholar]
- Naderi, A.; Nayebzadeh, H.; Gholami, S. Detection of Babesia infection among human, goats and sheep using microscopic and molecular methods in the city of Kuhdasht in Lorestan Province, West of Iran. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2017, 41, 837–842. [Google Scholar] [CrossRef]
- Dumanli, N.; Aktas, M.; Cetinkaya, B.; Cakmak, A.; Koroglu, E.; Saki, C.E. Prevalence and distribution of tropical theileriosis in eastern Turkey. Vet. Parasitol. 2005, 127, 9–15. [Google Scholar] [CrossRef]
- Iqbal, F.; Fatima, M.; Shahnawaz, S.; Naeem, M.; Shaikh, R.S.; Shaikh, A.S.; Aktas, M.; Ali, M. A Study on the determination of risk factors associated with babesiosis and prevalence of Babesia sp., by PCR amplification, in small ruminants from Southern Punjab (Pakistan). Parasite 2011, 18, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; Mofokeng, L.S.; Thekisoe, O.; MacLeod, E.T. Molecular survey for tick-borne pathogens and associated risk factors in sheep and goats in Kano Metropolis, Nigeria. Vet. Parasitol. Reg. Stud. Rep. 2022, 33, 100753. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Dumanli, N. Determination of prevalence and risk factors for infection with Babesia ovis in small ruminants from Turkey by polymerase chain reaction. Parasitol. Res. 2007, 100, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Aktas, M.; Dumanli, N. Detection of Babesia ovis by PCR in Rhipicephalus bursa collected from naturally infested sheep and goats. Res. Vet. Sci. 2008, 85, 116–119. [Google Scholar] [CrossRef]
- O’Connor, R.M.; Long, J.A.; Allred, D.R. Cytoadherence of Babesia bovis-infected erythrocytes to bovine brain capillary endothelial cells provides an in vitro model for sequestration. Infect. Immun. 1999, 67, 3921–3928. [Google Scholar] [CrossRef]
- Rjeibi, M.R.; Gharbi, M.; Mhadhbi, M.; Mabrouk, W.; Ayari, B.; Nasfi, I.; Jedidi, M.; Sassi, L.; Rekik, M.; Darghouth, M.A. Prevalence of piroplasms in small ruminants in North-West Tunisia and the first genetic characterisation of Babesia ovis in Africa. Parasite 2014, 21, 23. [Google Scholar] [CrossRef]
- Adua, M.M.; Idahor, K.O.; Panda, A.I.; Omeje, J.N. Prevalence of haemoparasites (Babesia species) in sheep under the traditional system of management in Lafia metropolis Nasarawa state, Nigeria. NSUK J. Sci. Technol. 2016, 6, 17–21. [Google Scholar]
- Carr, A.L.; Salgado, V.L. Ticks home in on body heat: A new understanding of Haller’s organ and repellent action. PLoS ONE 2019, 14, e0221659. [Google Scholar] [CrossRef]
- Bello, A.M.; Lawal, J.R.; Dauda, J.; Wakili, Y.; Mshellia, E.M.; Abubakar, M.I.; Biu, A.A. Prevalence of haemoparasite in balami sheep from Miduguri, Northeastern, Nigeria. Direct Res.J. Vet. Med. Anim. Sci. 2017, 2, 28–35. [Google Scholar]
- Ngole, I.U.; Ndamukong, K.J.; Mbuh, J.V. Internal parasites and haematological values in cattle slaughtered in Buea subdivision in Cameroun. Trop. Anim. Health Prod. 2003, 35, 409–413. [Google Scholar] [CrossRef]
- Masiga, D.K.; Okech, G.; Irungu, P.; Ouma, J.; Wekesa, S.; Ouma, B.; Guya, S.O.; Ndung’u, J.M. Growth and mortality in sheep and goats under high tsetse challenge in Kenya. Trop. Anim. Health Prod. 2002, 34, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Ademola, I.O.; Onyiche, T.E. Haemoparasites and haematological parameters of slaughtered Ruminants and pigs at Bodija Abattoir, Ibadan, Nigeria. Afr. J. Biomed. Res. 2013, 16, 101–105. [Google Scholar]
- Pedroni, M.J.; Sondgeroth, K.S.; Gallego-Lopez, G.M.; Echaide, I.; Lau, A.O. Comparative transcriptome analysis of geographically distinct virulent and attenuated Babesia bovis strains reveals similar gene expression changes through attenuation. BMC Genom. 2013, 14, 763. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, R.K.; Nimisha, M.; Sruthi, M.K.; Vidya, P.; Amrutha, B.R.; Kurbet, P.S.; Kumar, K.G.A.; Varghese, A.; Deepa, C.K.; Dinesh, C.N. Molecular characterization of South Indian field isolates of bovine Babesia spp. and Anaplasma spp. Parasitol. Res. 2018, 118, 617–630. [Google Scholar] [CrossRef]
- Laha, R.; Mondal, B.; Biswas, S.K.; Chand, K.; Das, M.; Sarma, D.; Goswami, A.; Sen, A. Detection of Babesia bigemina infection in cattle from North-Eastern India by polymerase chain reaction and its genetic relatedness with other isolates. Trop. Anim. Health Prod. 2015, 47, 633–636. [Google Scholar] [CrossRef]
- Lack, J.B.; Reichard, M.V.; van den Bussche, R.A. Phylogeny and evolution of the piroplasmida as inferred from 18S rRNA sequences. Int. J. Parasitol. 2012, 42, 353–363. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).