
Spatiotemporal Variation in Gross Primary Productivity and Their Responses to Climate in the Great Lakes Region of Sub-Saharan Africa during 2001–2020

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
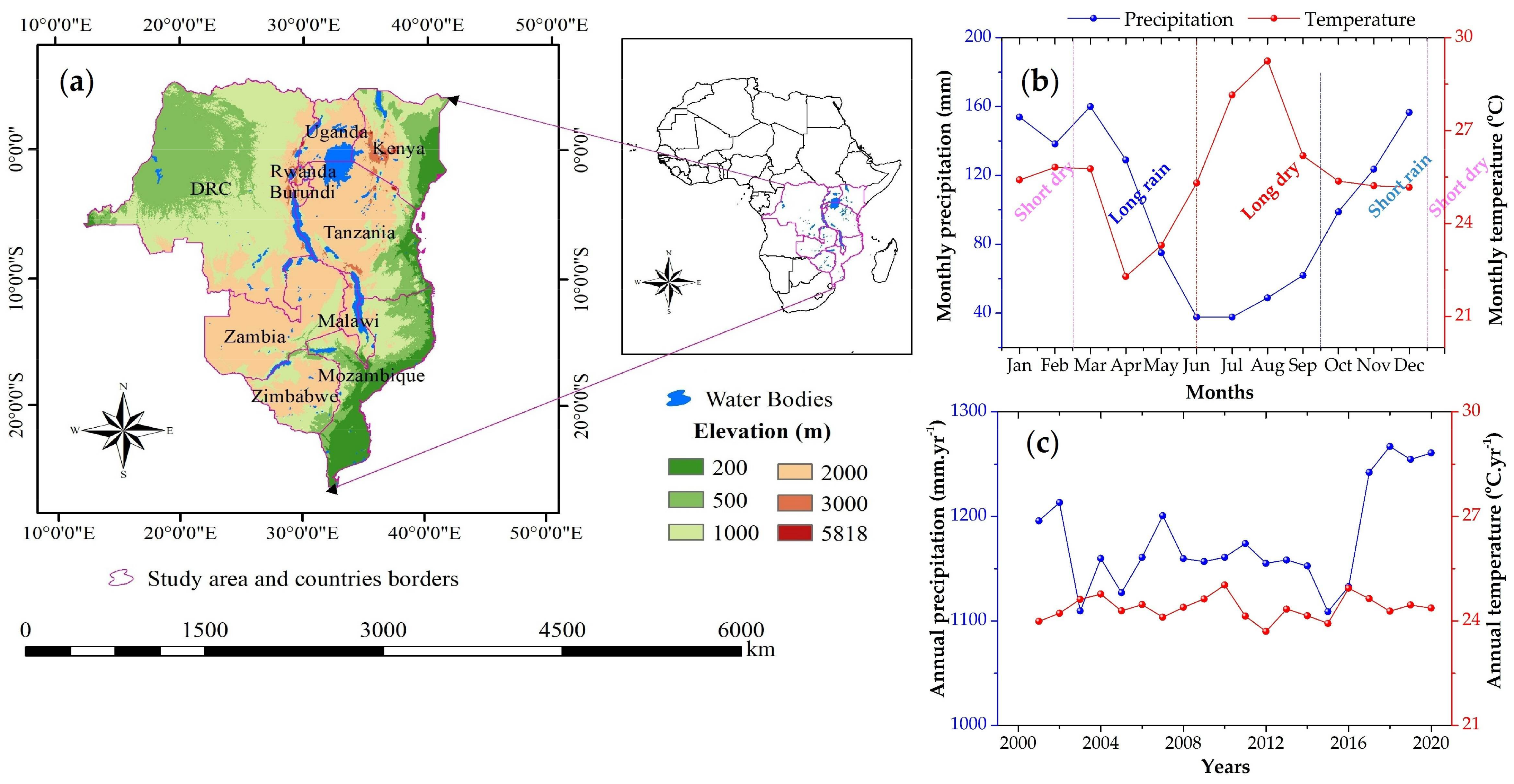
2.1. Study Area
2.2. Datasets
2.2.1. Vegetation Indices and Related Datasets
2.2.2. Climate Data
2.2.3. Field-Based GPP Data for Models Performance Validation
2.3. Methods
2.3.1. Data Processing
2.3.2. Descriptions of GPP Models
The CASA Model
The VPM Model
2.3.3. Estimation of Aridity Index
2.3.4. Evaluation of Model Performance
2.3.5. The Analysis of Spatial Relationship between GPP with Climate Factors
3. Results
3.1. The Spatial Patterns of GPP
3.2. Model Validation and GPP Simulation Stability
3.3. Seasonal and Inter-Annual Variations in GPP during 2001–2016
3.4. Analysis of GPP Variations among Individual Ecosystem Function Types (Biomes)
3.5. Spatial Variations of GPP in Response to Climate Factors
4. Discussion
4.1. Spatiotemporal Dynamics in GPP and GPP-LUE Models’ Performance Reliability
4.2. Seasonal and Inter-Annual Variability of GPP and Its Relationship with Climate
4.3. GPP Variation and Its Relationship with Climate Zones
4.4. Uncertainties and Future Directions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, Y.; Piao, S.; Li, X.; Chen, A.; Qin, D. Global patterns of vegetation carbon use efficiency and their climate drivers deduced from MODIS satellite data and process-based models. Agric. For. Meteorol. 2018, 256, 150–158. [Google Scholar] [CrossRef]
- Niu, S.; Fu, Z.; Luo, Y.; Stoy, P.C.; Keenan, T.F.; Poulter, B.; Zhang, L.; Piao, S.; Zhou, X.; Zheng, H.; et al. Interannual variability of ecosystem carbon exchange: From observation to prediction. Glob. Ecol. Biogeogr. 2017, 26, 1225–1237. [Google Scholar] [CrossRef]
- Lansø, A.S.; Smallman, T.L.; Christensen, J.H.; Williams, M.; Pilegaard, K.; Sørensen, L.L.; Geels, C. Simulating the atmospheric CO2 concentration across the heterogeneous landscape of Denmark using a coupled atmosphere-biosphere mesoscale model system. Biogeosciences 2019, 16, 1505–1524. [Google Scholar] [CrossRef] [Green Version]
- Golkar, F.; Shirvani, A. Spatial and temporal distribution and seasonal prediction of satellite measurement of CO2 concentration over Iran. Int. J. Remote Sens. 2020, 41, 8889–8907. [Google Scholar] [CrossRef]
- Madani, N.; Kimball, J.S.; Ballantyne, A.P.; Affleck, D.L.; Bodegom, P.M.; Reich, P.B.; Kattge, J.; Sala, A.; Nazeri, M.; Jones, M.O.; et al. Future global productivity will be affected by plant trait response to climate. Sci. Rep. 2018, 8, 2870. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Gowda, P.H.; Xiao, X.; Anup, K. Parameterizing ecosystem light use efficiency and water use efficiency to estimate maize gross primary production and evapotranspiration using MODIS EVI. Agric. For. Meteorol. 2016, 222, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tan, K.; Chen, B.; Du, P. Assessing the spatiotemporal variation and impact factors of net primary productivity in China. Sci. Rep. 2017, 7, 44415. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Yi, C.; Fang, W.; Hendrey, G. A global study of GPP focusing on light-use efficiency in a random forest regression model. Ecosphere 2017, 8, e01724. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Liu, X. Directly estimating diurnal changes in GPP for C3 and C4 crops using far-red sun-induced chlorophyll fluorescence. Agric. For. Meteorol. 2017, 232, 1–9. [Google Scholar] [CrossRef]
- Wu, Z.; Boke-Olen, N.; Fensholt, R.; Ardö, J.; Eklundh, L.; Lehsten, V. Effect of climate dataset selection on simulations of terrestrial GPP: Highest uncertainty for tropical regions. PLoS ONE 2018, 13, e0199383. [Google Scholar] [CrossRef]
- Cramer, W.; Kicklighter, D.W.; Bondeau, A.; Iii, B.M.; Churkina, G.; Nemry, B.; Ruimy, A.; Schloss, A.L. Comparing global models of terrestrial net primary productivity (NPP): Overview and key results. Glob. Change Biol. 1999, 5, 1–15. [Google Scholar] [CrossRef]
- Sannigrahi, S. Modeling terrestrial ecosystem productivity of an estuarine ecosystem in the Sundarban Biosphere Region, India using seven ecosystem models. Ecol. Model. 2017, 356, 73–90. [Google Scholar] [CrossRef]
- Kayiranga, A.; Chen, B.; Trisurat, Y.; Ndayisaba, F.; Sun, S.; Tuankrua, V.; Wang, F.; Karamage, F.; Measho, S.; Nthangeni, W.; et al. Water Use Efficiency-Based Multiscale Assessment of Ecohydrological Resilience to Ecosystem Shifts Over the Continent of Africa During 1992–2015. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005749. [Google Scholar] [CrossRef]
- Kayiranga, A.; Chen, B.; Guo, L.; Measho, S.; Hirwa, H.; Liu, S.; Bofana, J.; Sun, S.; Wang, F.; Karamage, F.; et al. Spatiotemporal variations of forest ecohydrological characteristics in the Lancang-Mekong region during 1992–2016 and 2020–2099 under different climate scenarios. Agric. For. Meteorol. 2021, 310, 108662. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, H.; Lin, A.; Zou, L.; Qin, W.; Du, Q. Evaluation of the latest MODIS GPP products across multiple biomes using Global Eddy Covariance Flux Data. Remote Sens. 2017, 9, 418. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Zhang, Y.; Jin, C.; Xiao, X. Comparison of solar-induced chlorophyll fluorescence, light-use efficiency, and process-based GPP models in maize. Ecol. Appl. 2016, 26, 1211–1222. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xiao, X.; Wagle, P.; Ma, S.; Baldocchi, D.; Carrara, A.; Zhang, Y.; Dong, J.; Qin, Y. Canopy and climate controls of gross primary production of Mediterranean-type deciduous and evergreen oak savannas. Agric. For. Meteorol. 2016, 226, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, X.; Li, R.; Long, F.; Zhang, L.; Zhang, Q.; Li, J. Water-use efficiency of an old-growth forest in lower subtropical China. Sci. Rep. 2017, 7, 42761. [Google Scholar] [CrossRef] [PubMed]
- Czubaszek, R. Exchange of Carbon Dioxide Between the Atmosphere and the Maize Field Fertilized with Digestate from Agricultural Biogas Plant. J. Ecol. Eng. 2019, 20, 145–151. [Google Scholar] [CrossRef]
- Measho, S.; Chen, B.; Trisurat, Y.; Pellikka, P.; Guo, L.; Arunyawat, S.; Tuankrua, V.; Ogbazghi, W.; Yemane, T. Spatio-Temporal Analysis of Vegetation Dynamics as a Response to Climate Variability and Drought Patterns in the Semiarid Region, Eritrea. Remote Sens. 2019, 11, 724. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.; Hao, X.; Qu, J.J. Monitoring Extreme Agricultural Drought over the Horn of Africa (HOA) Using Remote Sensing Measurements. Remote Sens. 2019, 11, 902. [Google Scholar] [CrossRef] [Green Version]
- ESA-CCI-LC. Land Cover CCI Product User Guide Version 2.0, Document Ref: CCI-LC-PUGV2. Available online: http://maps.elie.ucl.ac.be/CCI/viewer/download/ESACCI-LC-Ph2-PUGv2_2.0.pdf (accessed on 20 January 2020).
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations-the CRU TS3. 10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahayo, L.; Habiyaremye, G.; Kayiranga, A.; Kalisa, E.; Mupenzi, C.; Nsanzimana, D.F. Rainfall Variability and Its Impact on Rain-Fed Crop Production in Rwanda. Am. J. Soc. Sci. Res. 2018, 4, 9–15. [Google Scholar]
- Ndayisaba, F.; Guo, H.; Bao, A.; Guo, H.; Karamage, F.; Kayiranga, A. Understanding the spatial temporal vegetation dynamics in Rwanda. Remote Sens. 2016, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Karamage, F.; Zhang, C.; Fang, X.; Liu, T.; Ndayisaba, F.; Nahayo, L.; Kayiranga, A.; Nsengiyumva, J.B. Modeling rainfall-runoff response to land use and land cover change in Rwanda (1990–2016). Water 2017, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Dewitte, O.; Jones, A.; Spaargaren, O.; Breuning-Madsen, H.; Brossard, M.; Dampha, A.; Deckers, J.; Gallali, T.; Hallett, S.; Jones, R.; et al. Harmonisation of the soil map of Africa at the continental scale. Geoderma 2013, 211, 138–153. [Google Scholar] [CrossRef] [Green Version]
- Kayiranga, A.; Ndayisaba, F.; Nahayo, L.; Karamage, F.; Nsengiyumva, J.; Mupenzi, C.; Nyesheja, E. Analysis of climate and topography impacts on the spatial distribution of vegetation in the Virunga Volcanoes massif of east-central Africa. Geosciences 2017, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, K.; Sesha Sai, M.; Roy, P.; Dwevedi, R. Land Surface Water Index (LSWI) response to rainfall and NDVI using the MODIS Vegetation Index product. Int. J. Remote Sens. 2010, 31, 3987–4005. [Google Scholar] [CrossRef]
- USGS-NASA. Combined MODIS, NASA Satellite Data. Available online: https://lpdaac.usgs.gov/tools/data-pool/ (accessed on 9 December 2019).
- Engine, C. Cloud Computing of Climate and Remote Sensing Data. Available online: https://app.climateengine.org/ (accessed on 9 May 2019).
- Climate Database. Available online: https://climate.northwestknowledge.net/TERRACLIMATE/index_directDownloads.php (accessed on 1 May 2019).
- Jung, M.; Reichstein, M.; Margolis, H.A.; Cescatti, A.; Richardson, A.D.; Arain, M.A.; Arneth, A.; Bernhofer, C.; Bonal, D.; Chen, J.; et al. Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Koirala, S.; Weber, U.; Ichii, K.; Gans, F.; Camps-Valls, G.; Papale, D.; Schwalm, C.; Tramontana, G.; Reichstein, M.; et al. The FLUXCOM ensemble of global land-atmosphere energy fluxes. Sci. Data 2019, 6, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MTE-GPPEC. Available online: https://www.bgc-jena.mpg.de/geodb/projects/Home.php (accessed on 5 May 2019).
- Pötscher, B.M.; Preinerstorfer, D. Controlling the size of autocorrelation robust tests. J. Econom. 2018, 207, 406–431. [Google Scholar] [CrossRef] [Green Version]
- Polzehl, J.; Tabelow, K. Adaptive Smoothing of Digital Images: The R Package Adimpro; WIAS: Berlin, Germany, 2006. [Google Scholar]
- Zhang, Y.; Xiao, X.; Wu, X.; Zhou, S.; Zhang, G.; Qin, Y.; Dong, J. A global moderate resolution dataset of gross primary production of vegetation for 2000–2016. Sci. Data 2017, 4, 170165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, F.; Yokozawa, M.; Zhang, Z.; Xu, Y.; Hayashi, Y. Remote sensing of crop production in China by production efficiency models: Models comparisons, estimates and uncertainties. Ecol. Model. 2005, 183, 385–396. [Google Scholar] [CrossRef]
- Monteith, J.; Unsworth, M. Principles of Environmental Physics: Plants, Animals, and the Atmosphere; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Wu, J.; Guan, K.; Hayek, M.; Restrepo-Coupe, N.; Wiedemann, K.T.; Xu, X.; Wehr, R.; Christoffersen, B.O.; Miao, G.; da Silva, R.; et al. Partitioning controls on Amazon forest photosynthesis between environmental and biotic factors at hourly to interannual timescales. Glob. Chang. Biol. 2017, 23, 1240–1257. [Google Scholar] [CrossRef]
- Jin, C.; Xiao, X.; Merbold, L.; Arneth, A.; Veenendaal, E.; Kutsch, W.L. Phenology and gross primary production of two dominant savanna woodland ecosystems in Southern Africa. Remote Sens. Environ. 2013, 135, 189–201. [Google Scholar] [CrossRef]
- Wagle, P.; Xiao, X.; Torn, M.S.; Cook, D.R.; Matamala, R.; Fischer, M.L.; Jin, C.; Dong, J.; Biradar, C. Sensitivity of vegetation indices and gross primary production of tallgrass prairie to severe drought. Remote Sens. Environ. 2014, 152, 1–14. [Google Scholar] [CrossRef]
- Kalfas, J.L.; Xiao, X.; Vanegas, D.X.; Verma, S.B.; Suyker, A.E. Modeling gross primary production of irrigated and rain-fed maize using MODIS imagery and CO2 flux tower data. Agric. For. Meteorol. 2011, 151, 1514–1528. [Google Scholar] [CrossRef] [Green Version]
- Aber, J.D.; Reich, P.B.; Goulden, M.L. Extrapolating leaf CO2 exchange to the canopy: A generalized model of forest photosynthesis compared with measurements by eddy correlation. Oecologia 1996, 106, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.S. Evaluation of MODIS LAI, fAPAR and the relation between fAPAR and NDVI in a semi-arid environment using in situ measurements. Remote Sens. Environ. 2004, 91, 490–507. [Google Scholar] [CrossRef]
- Sedano, F.; Randerson, J. Multi-scale influence of vapor pressure deficit on fire ignition and spread in boreal forest ecosystems. Biogeosciences 2014, 11, 3739–3755. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, A.; Liu, W.; Ma, X.; Dong, G. Variation of aridity index and the role of climate variables in the Southwest China. Water 2017, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. FAO Rome 1998, 300, D05109. [Google Scholar]
- Prăvălie, R.; Bandoc, G. Aridity variability in the last five decades in the Dobrogea region, Romania. Arid. Land Res. Manag. 2015, 29, 265–287. [Google Scholar] [CrossRef]
- Trajkovic, S. Hargreaves versus Penman-Monteith under humid conditions. J. Irrig. Drain. Eng. 2007, 133, 38–42. [Google Scholar] [CrossRef]
- Mondal, A.; Khare, D.; Kundu, S. Change in rainfall erosivity in the past and future due to climate change in the central part of India. Int. Soil Water Conserv. Res. 2016, 4, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Su, Y.-C.; Shang, J.; Sha, J.; Li, X.; Sun, Y.-Y.; Ji, J.; Jin, B. Geographically Weighted Regression Effects on Soil Zinc Content Hyperspectral Modeling by Applying the Fractional-Order Differential. Remote Sens. 2019, 11, 636. [Google Scholar] [CrossRef] [Green Version]
- Puth, M.-T.; Neuhäuser, M.; Ruxton, G.D. Effective use of Pearson’s product–moment correlation coefficient. Anim. Behav. 2014, 93, 183–189. [Google Scholar] [CrossRef]
- O’Hara, R.; Merilä, J. Bias and precision in QST estimates: Problems and some solutions. Genetics 2005, 171, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Hao, Y.; Cui, X.; Chen, H.; Huang, S.; Du, Y.; Li, W.; Kardol, P.; Xiao, X.; Cui, L.; et al. Variability and changes in climate, phenology, and gross primary production of an alpine wetland ecosystem. Remote Sens. 2016, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Larsen, C.P. Effects of forest type and environmental factors on forest carbon use efficiency assessed using MODIS and FIA data across the eastern USA. Int. J. Remote Sens. 2013, 34, 8425–8448. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, G.; Yang, J.; Wimberly, M.C.; Zhang, X.; Tao, J.; Jiang, Y.; Zhu, J. Climate-driven global changes in carbon use efficiency. Glob. Ecol. Biogeogr. 2014, 23, 144–155. [Google Scholar] [CrossRef]
- Kayiranga, A.; Chen, B.; Zhang, H.; Nthangeni, W.; Measho, S.; Ndayisaba, F. Spatially explicit and multiscale ecosystem shift probabilities and risk severity assessments in the greater Mekong subregion over three decades. Sci. Total Environ. 2021, 798, 149281. [Google Scholar] [CrossRef] [PubMed]
- Giardina, C.P.; Ryan, M.G.; Binkley, D.; Fownes, J.H. Primary production and carbon allocation in relation to nutrient supply in a tropical experimental forest. Glob. Chang. Biol. 2003, 9, 1438–1450. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.M. Plant respiration in productivity models: Conceptualisation, representation and issues for global terrestrial carbon-cycle research. Funct. Plant Biol. 2003, 30, 171–186. [Google Scholar] [CrossRef]
- Jeong, S.-H.; Eom, J.-Y.; Lee, J.-H.; Lee, J.-S. Effect of rainfall events on soil carbon flux in mountain pastures. J. Ecol. Environ. 2017, 41, 37. [Google Scholar] [CrossRef] [Green Version]
- Laidler, K.J. A glossary of terms used in chemical kinetics, including reaction dynamics (IUPAC Recommendations 1996). Pure Appl. Chem. 1996, 68, 149–192. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Wu, W.; Liu, R. An overview of distributed activation energy model and its application in the pyrolysis of lignocellulosic biomass. Renew. Sustain. Energy Rev. 2014, 36, 236–246. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Guanter, L.; Zhou, S.; Ciais, P.; Joiner, J.; Sitch, S.; Wu, X.; Nabel, J.; Dong, J.; et al. Precipitation and carbon-water coupling jointly control the interannual variability of global land gross primary production. Sci. Rep. 2016, 6, 39748. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Piao, S.; Huang, M.; Ciais, P.; Zeng, Z.; Cheng, L.; Li, X.; Zhang, X.; Mao, J.; Peng, S.; et al. Global patterns and climate drivers of water-use efficiency in terrestrial ecosystems deduced from satellite-based datasets and carbon cycle models. Glob. Ecol. Biogeogr. 2016, 25, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Mawere, M. Underdevelopment, Development and the Future of Africa; Langaa Rpcig: Bamenda, Cameroon, 2017. [Google Scholar]
- Su, B.; Huang, J.; Fischer, T.; Wang, Y.; Kundzewicz, Z.W.; Zhai, J.; Sun, H.; Wang, A.; Zeng, X.; Wang, G.; et al. Drought losses in China might double between the 1.5 C and 2.0 C warming. Proc. Natl. Acad. Sci. USA 2018, 115, 10600–10605. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Sources | Datasets | Factors | Resolutions | |
|---|---|---|---|---|
| Spatial | Temporal | |||
| ISR | PAR | Vegetation status | 0.5° × 0.5° | daily |
| MODIS | FPAR | Vegetation status | 500 m | 8 days |
| NDVI | Vegetation indices | 500 m | 16 days | |
| EVI | Vegetation indices | 500 m | 8 days | |
| Land cover | Vegetation types | 500 m | yearly | |
| LSWI | Soil water content | 500 m | 8 days | |
| LAI | Vegetation canopy | 500 m | 16 days | |
| GPP (17A2H) | Carbon flux | 500 m | 8 days | |
| Global FLUXNET | GPPEC | Carbon flux | 0.5° × 0.5° | daily |
| TERRACLIMATE | Vapor pressure deficit | Soil water content | 0.5° × 0.5° | monthly |
| Soil moisture | Climate | 0.5° × 0.5° | monthly | |
| Wind speed | Climate | 0.5° × 0.5° | monthly | |
| Shortwave radiation | Climate | 0.5° × 0.5° | monthly | |
| Longwave radiation | Climate | 0.5° × 0.5° | monthly | |
| Land surface air temperature | Climate | 0.5° × 0.5° | monthly | |
| Solar radiation | Climate | 0.5° × 0.5° | monthly | |
| Soil heat density | Climate | 0.5° × 0.5° | monthly | |
| Evapotranspiration | Climate | 0.5° × 0.5° | monthly | |
| CLIMATEENGINE | Precipitation | Climate | 0.5° × 0.5° | monthly |
| Air temperature | Climate | 0.5° × 0.5° | monthly | |
| (Tmean, Tmin, Tmax) | Climate | 0.5° × 0.5° | monthly | |
| Biomes and Other LULC | Ratio (σ) | LUE (ε) Max | Tmin | Tmax | Topt |
|---|---|---|---|---|---|
| Evergreen needleleaf (EVGNL) | 0.5853 | 0.985 | 10 | 40 | 20 |
| Evergreen broadleaf (EVGB) | 0.4125 | 0.485 | 10 | 48 | 28 |
| Deciduous needleleaf (DecNL) | 0.5488 | 0.692 | 10 | 40 | 20 |
| Deciduous broadleaf (DECB) | 0.5488 | 0.542 | 10 | 40 | 20 |
| Grass and crop (Grass) | 0.5523 | 0.542 | 10 | 48 | 30 |
| Nonvegetated | na | 0.542 | 10 | 48 | 27 |
| Urban | na | 0.542 | 10 | 48 | 27 |
| Water bodies | na | 0.389 | 10 | 40 | 27 |
| Climate Zones | UNESCO (1979) | UNEP (1992) |
|---|---|---|
| Penman Method | Thornthwaite Method | |
| Hyper-Arid | <0.03 | <0.03 |
| Arid | 0.03–0.20 | 0.03–0.20 |
| Semi-Arid | 0.20–0.50 | 0.20–0.50 |
| Sub-Humid | 0.50–0.75 | 0.50–0.65 |
| Humid | >0.75 | >0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayiranga, A.; Chen, B.; Wang, F.; Nthangeni, W.; Dilawar, A.; Hategekimana, Y.; Zhang, H.; Guo, L. Spatiotemporal Variation in Gross Primary Productivity and Their Responses to Climate in the Great Lakes Region of Sub-Saharan Africa during 2001–2020. Sustainability 2022, 14, 2610. https://doi.org/10.3390/su14052610
Kayiranga A, Chen B, Wang F, Nthangeni W, Dilawar A, Hategekimana Y, Zhang H, Guo L. Spatiotemporal Variation in Gross Primary Productivity and Their Responses to Climate in the Great Lakes Region of Sub-Saharan Africa during 2001–2020. Sustainability. 2022; 14(5):2610. https://doi.org/10.3390/su14052610
Chicago/Turabian StyleKayiranga, Alphonse, Baozhang Chen, Fei Wang, Winny Nthangeni, Adil Dilawar, Yves Hategekimana, Huifang Zhang, and Lifeng Guo. 2022. "Spatiotemporal Variation in Gross Primary Productivity and Their Responses to Climate in the Great Lakes Region of Sub-Saharan Africa during 2001–2020" Sustainability 14, no. 5: 2610. https://doi.org/10.3390/su14052610