Abstract
This study explored the role of Aphanocladium album (strain MX95), Pleurotus ostreatus (strain ALPO) and Pleurotus eryngii (strain AL142PE) as potential biological limiters. MX95, ALPO and AL142PE were screened under laboratory studies against Phytophthora nicotianae (PN), Fusarium oxysporum f. sp. radicis-lycopersici (FORL), Fusarium oxysporum f. sp. lycopersici (FOL), Fusarium solani (FS), Sclerotinia minor (SM), Sclerotinia sclerotiorum (SS), Athelia (Sclerotium) rolfsii (AR) and Verticillium dahliae (VD). The radial growth inhibition and the over-growth of potential antagonists on the target organisms were used to assess the interactions in the in vitro dual culture plate assay. The antagonistic ability of each challenge isolate was evaluated by calculating an index of the antagonism (AI) based on the interaction type in the dual cultures. MX95, reducing the growth of SS (20%) and FS (40%), displayed deadlock at mycelial contact against FOL and FORL, deadlock at distance versus VD and completely over-grew PN and SM. ALPO reduced (43 to 88%) the mycelial growth of tested pathogens except FORL and replaced PN and VD. AL142PE reducing (53 to 67%) SS, VD, FS and FOL mycelial growth and completely over-grew PN. AR showed combative ability against all the experienced biological limiters. Based on the results of the AI values, MX95 (AI = 16.5) was considered an active antagonist, while ALPO (AI = 11.5) and AL142PE (AI = 12.0) were moderately active antagonists. Strains MX95, ALPO and AL142PE were suitable as environment-friendly potential biocontrol agents to manage some of the main soil-borne agents of foot, root, soft rot and wilt diseases. These results are the first step in the assessment of the potential capacity of these organisms as biological limiters. Nevertheless, additional experiments should be performed for the translation to the field conditions in plant protection against soil-borne plant pathogens. In particular, the optimisation of dose and application time validation should be performed for a solid conclusion about the competitive ability of MX95, ALPO and AL142PE and the usefulness of potential biological limiters.
1. Introduction
Plants are the primary source of nutrition for livestock and provide over 80% of the food consumed by humans [1]. Pathogens, including fungi, bacteria, viruses and nematodes, damage plants and their products, cause relevant economic losses to growers, increase prices of products to consumers and produce direct or indirect damage to the environment [1,2,3]. Some diseases make plant products unfit for human or animal consumption by contaminating them with poisonous structures (e.g., ergot from the sclerotia produced by Claviceps purpurea) or harmful microbial-based toxins associated, for example, to species of Aspergillus, Penicillium, Fusarium, Trichothecium, Myrothecium, Stachybotrys and other fungi [4,5].
Quarantine measures, crop certification, use of pathogen-free propagating material and plant resistance are aimed to exclude the pathogen from the host plants. If the unwanted microorganism is just introduced, eradication could eliminate, destroy or inactivate the inoculum. When the pathogen is already present, the development of resistant varieties through plant breeding, genetically engineered plants, use of agrochemicals and physical methods (i.e., heat treatments, UV irradiation, modified or controlled atmosphere, cold storage and inducing resistance by applying elicitors) and good agronomic and horticultural practices alone or in an integrated disease management approach may limit the pathogen spread, its harmfulness and progress and keep disease development at an acceptable level [4,6]. Advancements in biotechnology, microbiology, bioinformatics as well as information and communication technology have given new strategies for plant-disease management [7].
Strategies found on chemicals management protect the host plant and/or eradicate an existing infection. Adversely, chemicals application may impact the health of agricultural workers and consumers and drive the development of pathogen resistance [8,9]. In the past recent years, large numbers of synthetic agrochemicals have been banned due to their toxicity in animals and humans. The development of environment-friendly and sustainable agriculture also improved the research focused on developing alternative inputs to chemicals for controlling the agents of damage and disease on plants. These alternatives included those referred to as “Biological controls” or “Biocontrol” [10,11].
The term “Biocontrol” has been used in different fields of biology, most notably entomology and plant pathology. In entomology, it describes the use of predatory insects, entomopathogenic nematodes or microbial pathogens to suppress populations of different insects [11,12]. In plant pathology, biocontrol applies microbial antagonists to suppress diseases development [2,11,13,14]. In both ambits, the organism that suppresses the pest or the pathogen is referred to as the biological control agent (BCA). More broadly, biological control comprised the natural products extracted or fermented from various sources able to reduce the effect of plant-pathogens action directly or through the activation of biological mechanisms. The microenvironment manipulation to favour the activity of antagonists was also enclosed. Cultural practices such as crop rotations and cropping disease-resistant cultivars, suppressing organisms causing plant diseases, would be included in the definition. More narrowly, biocontrol refers to the introduction of living organisms, other than disease-resistant host plants, to suppress the activities and populations of one or more plant pathogens [11].
Intensively studied BCAs agents are bacterial (species of the genera Bacillus, Burkholderia, Lysobacter, Pantoea, Pseudomonas and Streptomyces), fungal (species of the genera Ampelomyces, Coniothyrium, Dactylella, Gliocladium, Paecilomyces, Aspergillus, Penicillium, Trichoderma, avirulent strains of Fusarium oxysporum and binucleate Rhizoctonia-like fungi), the Chromist Pythium oligandrum and mycorrhizal fungi such as Pisolithus and Glomus spp. [10,11,15]. Ampelomyces quisqualis (AQ10™ Biogard, Italy) and Pseudozyma (Anthracocystis) flocculosa (Sporodex™ Plant Products Co. Ltd, Canada) control powdery mildew. Non-pathogenic Fusarium oxysporum (Fusaclean™ Natural Plant Production, France, Biofox C™ SIAPA, Italy) prevent Fusarium wilt diseases. Phlebiopsis gigantea (Rotstop™ Verdera, Finland) and Paraphaeosphaeria (Coniothyrium) minitans (Contans WG™ Bayer crop science, Italy; KONI™ Bioved Ltd, Hungary) reduce the incidence of root rot diseases by Heterobasidion annosum and watery soft rot caused by Sclerotinia species, respectively. Clonostachys rosea (syn. Gliocladium catenulatum) is effective against damping-off, seed rot, root, stem rot and wilt diseases (Primastop™ AgBio Development, Inc, USA), soil-borne and foliar diseases of greenhouse vegetables, herbs and ornamentals (Prestop™ Verdera, Finland). Yarrowia lipolytica (Aspire™ Ecogen, USA) is also effective against agents of post-harvest diseases. Several products use Trichoderma species as an active BCA. Remedier™ (ISAGRO S.p.A., Italy), based on Trichoderma asperellum and Trichoderma gamsii, and Binab T™ (BINAB Bio-Innovation AB, Sweden), containing Trichoderma harzianum and Trichoderma polysporum, are effective against root and collar diseases and protect wounds in ornamental, shade, forest and fruits trees. Remedier also prevents “Esca” and other trunk diseases of vine. T. harzianum as RootShield™ (BioWorks, Inc. USA), is specific against root and foot rot diseases, Pythium, Fusarium, Rhizoctonia, Thielaviopsis and Cylindrocladium species, while Trichodex™ (Makhteshim Chemical Works Ltd., Israel) works well against grey mould, Rhizoctonia, Sclerotinia and Colletotrichum species. Trichoderma viride (Trieco™ Ecosense Labs, India) and T. virens (SoilGard™ Certis, USA) fight soil-borne fungi. SoilGard is also specific against species of the genera Rhizoctonia and Pythium [2,11].
The present study aimed to investigate the antagonistic activity of three new biological limiters against seven soil-borne phytopathogens agents of the most important foot, root and wilt diseases of several plant species.
2. Materials and Methods
2.1. Strains and Media
The strains of potential BCA and target organisms listed in Table 1 were used in this study.

Table 1.
Potential biological control agents and phytopathogens strains used in this study.
Isolates of target organisms were stored at 5 ± 1 °C on sterile soil according to Toussoun and Nelson [16]. PN and the potential BCAs were stored on Potato Dextrose Agar (PDA) slant tubes in the culture collection of the Department of Soil, Plant and Food Sciences—Plant Pathology Section at the University of Bari Aldo Moro. All the strains were revitalised and routinely grown on PDA at 25 ± 1 °C in the dark.
2.2. Growth Rate
In determining the growth rate of mycelium, antagonists and target pathogens were singly grown in a 90 mm Petri dish containing 18 mL PDA. A plug (3 mm in diameter) of each isolate, collected from actively grown cultures, was placed 1 cm far from the border of the plate on the line of dish diameter. Inoculate plates were sealed with Parafilm and incubated at 25 ± 1 °C in darkness. Radius measurements were made every eight hours following the line of dish diameter. All isolates were tested in triplicates and the experiment was repeated at least two times.
2.3. In Vitro Dual Culture Bioassays
The antagonistic potential of selected biological limiters was tested in dual culture assay on PDA medium in 90 mm Petri dishes. Each plate was seeded with a 3 mm diameter disc cut from the edge side of an actively growing pure culture of pathogen and potential antagonist. Pathogen and antagonist plugs were placed together in the same plate on opposite sides, 1 cm far from the border of the Petri dish. As a control, pathogen and potential antagonist plugs were placed alone. Inoculate plates were sealed with Parafilm and incubated at 25 ± 1 °C, in the dark. All dual cultures were made in triplicates and repeated at least two times. The cultures were observed every eight hours to record the time of the first contact between the two mycelia.
Radial colony growth of the pathogen in the direction of the potential BCA and its growth on the control plate was also measured daily. The readings assessed at 10 days after inoculation were used to calculate the percentage of inhibition of radial growth (IRG) as , where R1 = average of radial growth (mm) of the pathogen in the control plates, R2 = average of radial growth (mm) of the pathogen in dual cultures.
The percentage of over-growth of antagonist (OA) on the target organism was calculated 18 days after co-inoculation using the formula , where OGA = average over-growth (mm) of the potential BCA on the target pathogen, R2 = average of radial growth (mm) of the pathogen in presence of the antagonist.
Mycelial interactions in dual culture were scored under a stereomicroscope every 2 days and were determined using the scale reported in Table 2.
Table 2.
Rating scale used to detect the antagonistic ability of each tested isolate.
For each tested BCA, the antagonism index (AI) [17,18] was calculated as , where n = of each type of reaction and i = corresponding score (Table 1). Tested BCAs were considered active antagonists (AI > 15), moderately active antagonists (AI = 15–10) and weak antagonists (AI < 10).
2.4. Statistical Analysis
Plates for growth rate, ICR and OA were allowed in a randomised design. Homogeneity of variances was analysed by Levene’s test. The data obtained for each set of experiments were subject to Analysis of Variance (ANOVA) using the statistical package SAS version 9.0 for Windows. The ICR and OA data were analysed as radius values and expressed as a percentage in the figures. The pairwise comparison of means was performed with the Fisher Least Significant Difference (LSD) test at p = 0.05.
3. Results
3.1. Growth Rate
All the tested strains developed a different growth rate according to the species considered. After 7 days survey, MX95, ALPO and AL142PE reached a radius of 44.00, 52.00 and 62.00 mm, respectively (Figure 1).
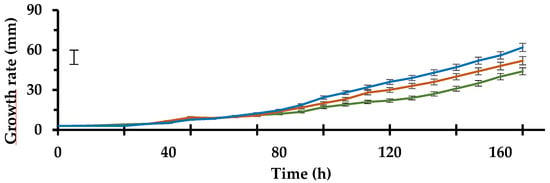
Figure 1.
Growth rate in 90 mm Petri dishes containing Potato Dextrose Agar of tested antagonists: MX95 (—), ALPO (—), AL142PO (—). Data are the means of six replicates ± standard deviations. The vertical bar indicates the Fisher’s LSD at p = 0.05. For acronym definitions, see Table 1.
Among the tested pathogens, SM and SS were the fastest and colonised the entire plate in under 96 h (Figure 2), while the lower growth rate was recorded for VD (23 mm during 4 days).
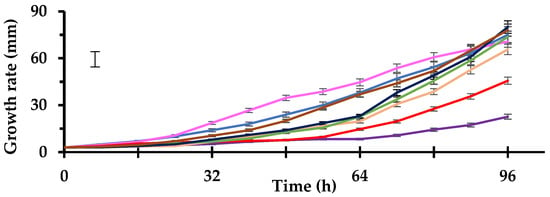
Figure 2.
Growth rate in 90 mm Petri dishes containing Potato Dextrose Agar of target pathogens: SS (—), SM (—), FS (—), FOL (—), FORL (—), AR (—), VD (—) and PN (—). Data are the means of six replicates ± standard deviations. The vertical bar indicates the Fisher’s LSD at p = 0.05. For acronym definitions, see Table 1.
3.2. In Vitro Dual Culture Bioassays
The time required for the first contact (Table 3) between the potential BCA and the target pathogen ranged from 56 h to the no contact associated with deadlock at distance.
Table 3.
Time (hours) required for the first contact between the tested antagonist Aphanocladium album (MX95), Pleurotus ostreatus (ALPO) or Pleurotus eryngii (AL142PE) and the tested phytopathogenic organisms.
Dual culture assays showed different types of interaction between the response and challenge fungal isolates (Figure 3, Figure 4 and Figure 5).
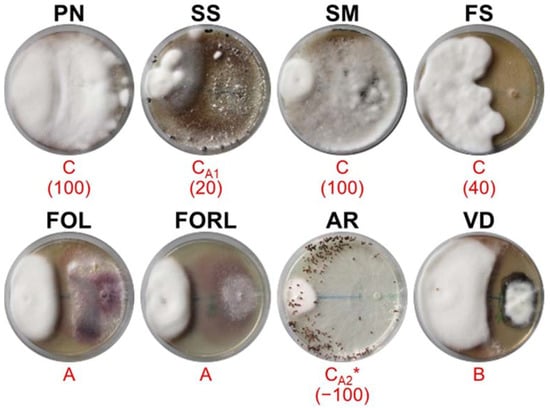
Figure 3.
Mycelial interactions after 28 days of co-incubation on PDA between the antagonist MX95 (left) and the target pathogens (right). For acronym definitions, see Table 1. Red letters show the type of interactions as described in Table 2. * indicates the partial or complete replacement of antagonist by the pathogen. In brackets is the percentage of over-growth 18 days after co-inoculation.
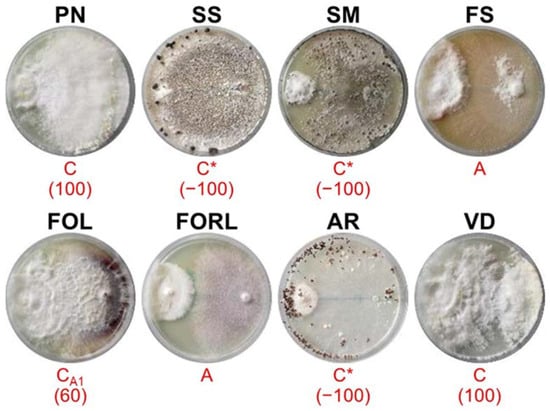
Figure 4.
Mycelial interactions after 28 days of co-incubation on PDA between the antagonist ALPO (left) and the target pathogens (right). For acronym definitions, see Table 1. Red letters show the type of interactions as described in Table 2. * indicates the partial or complete replacement of antagonist by the pathogen. In brackets is the percentage of over-growth 18 days after co-inoculation.
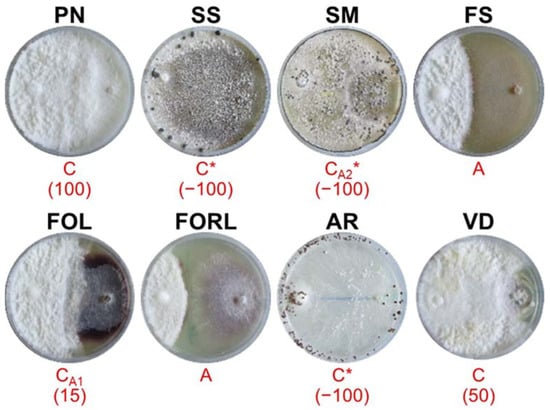
Figure 5.
Mycelial interactions after 28 days of co-incubation on PDA between the antagonist AL142PE (left) and the target pathogens (right). For acronym definitions, see Table 1. Red letters show the type of interactions as described in Table 2. * indicates the partial or complete replacement of antagonist by the pathogen. In brackets is the percentage of over-growth 18 days after co-inoculation.
MX95 (Figure 3) showed deadlock after mycelial contact during the interactions with FOL and FORL, while deadlock at a distance was exhibited with VD. MX95 completely over-grew (OA = 100%) SM and PN, while it partially grew on FS (OA = 40%) and SS (OA = 10%). AR in part replaced MX95.
ALPO (Figure 4) showed deadlock after mycelial contact during the interactions with FS and FORL, grew on PN (OA = 100%), FOL (OA = 60%) and VD (OA = 100%) and was completely replaced by SS, SM and AR.
AL142PE (Figure 5) showed deadlock after mycelial contact during the interactions with FS and FORL, over-grew PN (OA = 100%), VD (OA = 50%) and FOL (OA = 15%) colony surfaces and was completely replaced by SS, SM and AR.
Replacement of pathogen by the antagonist was more frequent (41.7%) than deadlock (29.2%). Furthermore, in the 29.2% of tested interactions, the target pathogen was able to replace partially or completely the tested antagonist (Table 4).
Table 4.
Frequency of type and subtype of interactions between mycelium of tested antagonist and target pathogen in dual culture experiments on Potato Dextrose Agar medium expressed as a percentage of the total number (144) of pairings tested (a).
MX95 reached an average IRG of 40.8% and was effective to inhibit the growth of FOL and PN, while was less effective against AR (Figure 6). ALPO, showing an average IRG of 56.3%, had a strong inhibitory capacity towards SM, SS and AR, while it was less effective against FORL (Figure 6). AL142PE showed a high efficacy to contrast AR, VD, SS and FS and achieved an average IRG of 53.1% (Figure 6).
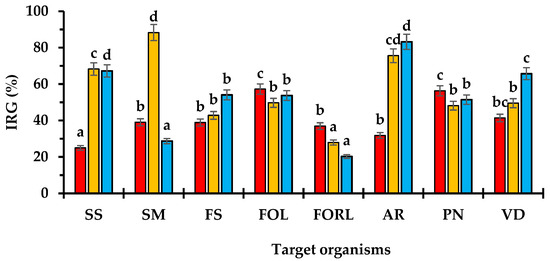
Figure 6.
Percentage of inhibition radial growth (IRG) assessed 10 days after co-inoculation of MX95 (■), ALPO (■) and AL142PE (■) against SS, SM, FS, FOL, FORL, AR, PN and VD. Data are the means of six replicates ± standard deviations. For each antagonist, values accompanied by the same letters are not significantly different (p ≤ 0.05) according to Fisher’s LSD test. For acronym definitions, see Table 1.
Based on the AI values, MX95 (AI = 16.5) was considered an active antagonist, while ALPO (AI = 11.5) and AL142PE (AI = 12.0) were moderately active antagonists.
4. Discussion
Soil-borne pathogens cause, every year, considerable agricultural crop losses, and their management is identified as one of the top farm management issues faced by farmers around the world [19]. The most common and destructive foot, root, soft rot and wilt diseases of cropped plant in the field and protected crops are associated with Phytophthora nicotianae, Fusarium oxysporum f. sp. radicis-lycopersici, Fusarium oxysporum f. sp. lycopersici, Fusarium solani, Sclerotinia minor, Sclerotinia sclerotiorum, Athelia rolfsii and Verticillium dahliae.
Agricultural chemicals are commonly used for the management of soil-borne pathogens. However, the high frequency of chemical use, non-target effects, development of pathogen resistance to chemical pesticides, risks to human health and the surrounding environment and phasing out of some effective soil fumigants such as methyl bromide have encouraged the development of alternative environmentally friendly methods for disease management [20,21,22,23].
A wide range of chromists, fungi, bacteria and viruses control plant parasitic nematodes, plant pathogens including fungi, oomycetes, bacteria and viruses and reduce disease development [10,11,12]. Hyperparasitism and hypovirulence are examples of mechanisms expressed by BCAs during direct antagonism. In contrast, antibiotics, lytic enzymes and other by-products of microbial life mediate BCA suppression effects during indirect antagonisms. The most abundant non-pathogenic plant-associated microbes protect the plant by rapid colonisation of space and exhausting the limited available substrates so that none are available for pathogens to grow in the rhizosphere and on the plant surface. Stimulation of plant host defence pathways by BCAs is a form of indirect antagonism. However, in the context of the natural environment, mechanisms associated with pathogen suppression will be modulated by the relative occurrence of other organisms in addition to the pathogen [11,24,25,26,27,28,29,30,31,32,33]. Contributing to disease control are organisms classified as competitive saprophytes, facultative plant symbionts and facultative hyperparasites. These can generally survive on dead plant material, but they colonise and express biocontrol activities while growing on plant tissues [11].
Most BCAs of plant pathogens are fungi because these organisms are self-propagating, have a high reproductive rate (sexually as well as asexually), have a short generation time, are target specific and, in the absence of the host, they can survive as saprotrophic [2].
In this study, the potential antagonistic capacity of Aphanocladium album strain MX95, Pleurotus ostreatus strain ALPO and Pleurotus eryngii strain AL142PE was evaluated in dual culture bioassays against the eight strains of phytopathogenic organisms tested.
The in vitro dual culture test excludes environmental factors that may impact practical biocontrol application, confirms the ability to show rapidly and clearly the mutual effects of the paired organisms and provides a preliminary screening of the interaction between antagonist and pathogen. In terms of effectiveness, the potential antagonists herein tested were mainly species dependent. The growth rate on the PDA plate of potential antagonist shares out a daily increase of 4.5 ± 0.5, 5.1 ± 0.4 and 5.8 ± 0.5 mm for MX95, ALPO and AL142PE, respectively.
Different behaviour in growth characterised these eight tested plant pathogens. VD was the slowest with a daily increase in growth calculated at 5.76 mm, while SM was faster reaching a daily increase in growth of 20 mm. SS, AR, PN, FORL, FS and FOL showed a daily increase in growth in the range 19.5–11.4 mm. These different growth abilities could support antagonists and pathogens during the interaction in dual cultures and explain the effects on inhibition of radial growth, over-growth of antagonist and pathogens, the type of interaction and, therefore, the antagonism index. The mycelial growth rate also influenced the time required for the first contact between the antagonist and the target pathogen. Low time for the first contact was associated with antagonist or target pathogens with fast mycelial growth.
Deadlock at distance or with initial contact, partial or complete replacement were the types of reaction observed in these studies as signs of antagonisms between tested organisms. The complete (CB1) or partial (CB2) replacement after an initial deadlock at a distance and complete replacement after initial deadlock with mycelial contact (CA2) never occurred. All dual culture plates exhibited signs of interaction between the antagonist species and the target pathogens. Of the tested pathogenic organisms, AR showed combative ability against AL142PE, ALPO and MX95.
A. album is an Ascomycota belonging to the Nectriaceae family characterised for its capacity to survive for a long time and to sporulate on poor substrates. It is considered a necrotrophic mycoparasite able to produce hydrolytic enzymes such as protease, gluconase and several chitinases involved in cell wall degradation of many phytopathogenic fungi [34]. This fungus can grow over and around uredia of the rusts Puccinia coronata, Puccinia hordei, Puccinia graminis f. sp. avenue and Puccinia recondita f. sp. triticina under very humid conditions [35]. The strain MX95 of A. album (patent MI2006A000503) was tested as a biological limiter against Golovinomyces (Oidium) lycopersici (agent of powdery mildew on tomato and squash), Podosphaera (Sphaerotheca) fusca (the agent of powdery mildew on cucumber) and other agents of foliar diseases [36,37]. Furthermore, on tomato, A. album MX95 was an efficient limiter of Pseudopyrenochaeta (Pyrenochaeta) lycopersici (the agent of Corky Root) and the root-knot nematode Meloidogyne incognita [38]. Moreover, A. album strain MX95 significantly decreased root gall formation by Meloidogyne javanica in infected tomato plants, improved plant fitness and increased rhizosphere microbial populations [39]. Finally, MX95 showed a satisfactory efficacy to control post-harvest rot diseases of grape in pre- and post-harvest [40].
A. album produced extracellular chitinase when grown in minimal medium with chitin (a linear polymer composed of repeating β(1,4)-N-acetylglucosamine) as the sole carbon source. Chitin is a biopolymer ingredient of the exoskeletons of arthropods and in the cell walls of fungi. This molecule is hydrolysed by endo-chitinases (chitin glucanohydrolase, EC 3.2.1.14) to β(1,4)-N-acetylglucosamine oligomers and chitobiose [41,42]. Chitinases are considered one of the chemicals produced by A. album anti-fungal activity [34,42]. Considering the low substrate colonisation rate of this organism, the results are encouraging and make it a potential biocontrol agent.
The oyster mushrooms (Pleurotus spp.) are in third place after the white button (Agaricus bisporus) and shiitake (Lentinula edodes) among world mushroom production [43]. Several strains of P. eryngii and P. ostreatus are extensively cultivated in the world due to their excellent gastronomical qualities and longer shelf life. The species of the Pleurotus genus are xylotrophic mushrooms widely distributed in nature. Strains of P. ostreatus showed a strong inhibitory activity on the mycelia growth of Ceratobasidium cereale (syn. Rhizoctonia cerealis), Gaeumannomyces tritici (formerly Gaeumannomyces graminis var. tritici), Fusarium culmorum and Bipolaris sorokiniana, reaching an antagonism index value of 18.0 [17]. P. ostreatus also had a strong combative ability against mycoparasitic fungi as Clonostachys rosea, T. harzianum, Tricoderma pseudokoningii and T. viride with an antagonism index value of 13.5 [18]. Furthermore, strains of P. ostreatus and P. eryngii were tested as BCAs of sugar beet nematode Heterodera schachtii [43]. Here, the strains ALPO and AL142PE, despite their low growth rate, showed a variable inhibitory activity depending on the target pathogen considered. These strains caused deadlock at mycelial contact against FS and FOL and high inhibitory activity against VD, PN and SS. Strains of SM and AR partially or completely replaced the two tested Pleurotus antagonists. The activity as BCAs of the two Pleurotus strains could be associated with laccases production as a defensive response against mycelial invasion [44].
Based on the AI values, fungal isolates can be divided into three categories according to Badalyan et al. [17,18]: (1) active, with AI > 15, (2) moderately active, with AI between 10 and 15, and (3) weakly active, with AI < 10. A lower index of antagonism is associated with a weaker inhibition response to the paired isolate [45]. AI is a qualitative measure defined as the ability of a fungus to dominate and compete with other species [46]. Higher AI denotes the higher competitive and inhibitory ability of paired isolate. Our experiment revealed MX95 (AI = 16.5) as an active antagonist, while ALPO (AI = 11.5) and AL142PE (AI = 12.0) were moderately active in inhibition according to the AI.
MX95 showed deadlock at mycelial contact against the tested strains of F. oxysporum f. sp. lycopersici and F. oxysporum f. sp. radicis-lycopersici and deadlock at distance with V. dahliae. Over-growth without initial deadlock was the effect of MX95 against the strains of S. minor, F. solani and P. nicotianae. Meanwhile, partial replacement after initial deadlock with mycelial contact was recorded in the interaction with S. sclerotiorum strain. The strain of A. rolfsii was instead the unique pathogen able to over-grow MX95.
The strains ALPO of P ostreatus and AL142PE of P. eryngii caused deadlock at mycelial contact against the tested strains of F. solani and F. oxysporum f. sp. lycopersici and high inhibitory activity against the tested strains of V. dahliae, P. nicotianae and S. sclerotiorum. The strains of S. minor and A. rolfsii partially or completely replaced the two tested Pleurotus antagonists.
MX95, ALPO and AL142PE are suitable as environment-friendly potential BCAs to manage some very destructive soil-borne pathogens of plants as an alternative to synthetic chemicals.
Finally, we would like to point out a drawback of this study in the light of the interpretation and generalisation of the obtained results. Because of the low number of repetitions and only one tested reference isolate of each target pathogen, the results should be treated with caution. Further tests are needed to verify the universality of the obtained findings, with a wider range of response isolates of S. sclerotiorum, S. minor, F. solani, F. oxysporum f. sp. lycopersici, F. oxysporum f. sp. radicis-lycopersici, V. dahliae, A. rolfsii and P. nicotianae. A. album MX95 treatments could improve plant fitness and the rhizosphere microbiome, increasing bacterial diversity of the rhizosphere [39]. The application of the Pleurotus ostreatus and P. eryngii exhausted cultivation residual biomass could manage soil-borne plant-pathogens, increase the soil organic matter content, nitrogen and other plant macro and micro-nutrients overall in semiarid and arid soils.
5. Conclusions and Future Trend
Against the eight strains of target pathogens, the three BCAs here tested revealed different types of interactions in dual cultures on PDA. A. album strain MX95 showed two types of competitive interactions: (i) deadlock, consisting of mutual inhibition after mycelial contact against FOL and FORL, (ii) deadlock at a distance against VD and (iii) replacement against PN, SS, SM and FS. Moreover, MX95 was completely replaced by AR. P. ostreatus strain ALPO and P. eryngii strain AL142PE demonstrated: (i) deadlock after mycelial contact against FS and FORL and (ii) replacement against PN, FOL and VD. Furthermore, ALPO and AL142PE were completely replaced by SS, SM and AR. The results of the antagonism index suggested that MX95 was the most competitive and had the highest inhibition of PN, SM, FS, FOL, FORL and VD growth, while ALPO and AL142PE were competitive with PN, FS, FOL, FORL and VD. These isolates are promising candidates for use as biological limiters, but additional experiments with different isolates of S. sclerotiorum, S. minor, F. solani, F. oxysporum f. sp. lycopersici, F. oxysporum f. sp. radicis-lycopersici, V. dahliae, A. rolfsii and P. nicotianae should be conducted for confirmation and clarification of our results. Based on the results of these preliminary studies, we can assume that A. album strain MX95, P. ostreatus strain ALPO and P. eryngii strain AL142PE can be favourably considered as a new BCAs suitable to use in plant protection for the control of soil-borne plant pathogens. Nevertheless, additional tests and supplementary experiments should be performed for a solid conclusion. However, further research is still needed to optimise product rates and methods of application (e.g., dose and time) along with field validation experiments.
Author Contributions
Conceptualisation, G.D., C.C., F.M. and G.L.B.; data curation, G.D. and G.L.B.; investigation, G.D., C.C., F.M. and G.L.B.; writing—original draft, G.D., C.C. and G.L.B.; writing—review and editing, G.D., C.C., F.M. and G.L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Tonia Gemma provided helpful discussions and skilful proof reading of the manuscript of this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rizzo, D.M.; Lichtveld, M.; Mazet, J.A.K.; Togami, E.; Miller, S.A. Plant health and its effects on food safety and security in a One Health framework: Four case studies. One Health Outlook 2021, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Ann. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: San Diego, CA, USA, 2005; p. 992. [Google Scholar]
- Kuhn, D.M.; Ghannoum, M.A. Indoor Mold, Toxigenic Fungi, and Stachybotrys chartarum: Infectious disease perspective. Clin. Microbiol. Rev. 2003, 16, 144–172. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Chawla, S. Cultural Practices: An eco-friendly innovative approaches in plant disease management. In Eco-Friendly Innovative Approaches in Plant Disease Management; Singh, V.K., Singh, Y., Singh, A., Eds.; International Book Publisher: Dehradun, India, 2012; pp. 1–20. [Google Scholar]
- Haq, I.U.; Ijaz, S. Plant Disease Management Strategies for Sustainable Agriculture through Traditional and Modern Approaches Series. Volume 13: Sustainability in Plant and Crop Protection; Springer Nature: Cham, Switzerland, 2020; p. 339. [Google Scholar]
- Ramakrishnan, B.; Venkateswarlu, K.; Sethunathan, N.; Megharaj, M. Local applications but global implications: Can pesticides drive microorganisms to develop antimicrobial resistance? Sci. Total Environ. 2019, 654, 177–189. [Google Scholar] [CrossRef]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef]
- Nega, A. Review on Concepts in Biological Control of Plant Pathogens. J. Biol. Agric. Health. 2014, 4, 33–55. [Google Scholar]
- Pal, K.K.; McSpadden Gardener, B. Biological Control of Plant Pathogens. Plant Health Inst. 2006, 10, 1117–1122. [Google Scholar] [CrossRef]
- Andrews, J.H. Biological control in the phyllosphere. Ann. Rev. Phytopathol. 1992, 30, 603–635. [Google Scholar] [CrossRef]
- Stirling, M.; Stirling, G. Disease Management: Biological Control. In Plant Pathogens and Plant Diseases; Brown, J., Ogle, H., Eds.; Rockvale Publications: Armidale, Australia, 1997; pp. 427–439. [Google Scholar]
- Baker, K.F. Evolving concepts of biological control of plant pathogens. Ann. Rev. Phytopathol. 1987, 25, 67–85. [Google Scholar] [CrossRef]
- Bubici, G. Streptomyces spp. as biocontrol agents against Fusarium species. CAB Rev. 2018, 13, 050. [Google Scholar] [CrossRef]
- Toussoun, T.A.; Nelson, P.E. A Pictorial Guide to the Identification of Fusarium Species according to the Taxonomic System of Snyder and Hansen; Pennsylvania State University Press: London, UK, 1968; p. 51. [Google Scholar]
- Badalyan, S.M.; Innocenti, G.; Garibyan, N.G. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Phytopathol. Mediterr. 2002, 41, 200–225. [Google Scholar] [CrossRef]
- Badalyan, S.M.; Innocenti, G.; Garibyan, N.G. Interactions between xylotrophic mushrooms and mycoparasitic fungi in dual culture. Phytopathol. Mediterr. 2004, 43, 44–48. [Google Scholar] [CrossRef]
- Katan, J. Interactions of soilborne pathogens with roots and aboveground plant organs. In Plant Roots: The Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Basel Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 949–959. [Google Scholar]
- Martin, F.N. Development of alternative strategies for management of soilborne pathogens currently controlled with methyl bromide. Annu. Rev. Phytopathol. 2003, 41, 325–350. [Google Scholar] [CrossRef]
- Kiss, L. How dangerous is the use of fungal biocontrol agents to nontarget organisms? New Phytol. 2004, 163, 453–455. [Google Scholar] [CrossRef]
- Whipps, J.M.; Lumsden, R.D. Commercial use of fungi as plant disease biological control agents: Status and prospects. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 9–22. [Google Scholar]
- Klokočar-Šmit, Z.D.; Lević, J.T.; Maširević, S.N.; Grozdanović-Varga, J.M.; Vasić, M.A.; Aleksić, S.R. Fusarium rot of onion and possible use of bioproduct. Proc. Matica Srp. Nat. Sci. 2008, 114, 135–148. [Google Scholar] [CrossRef]
- Lo, C.T. General mechanisms of action of microbial biocontrol agents. Plant Pathol. Bul. 1998, 7, 155–166. [Google Scholar]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Ann. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef]
- Islam, T.M.; Hashidoko, Y.; Deora, A.; Ito, T.; Tahara, S. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne peronosporomycetes. Appl. Environ. Microb. 2005, 71, 3786–3796. [Google Scholar] [CrossRef]
- Benhamou, N.; Chet, I. Cellular and molecular mechanisms involved in the intersection between Trichoderma harzianum and Pythium ultimum. Appl. Environ. Microb. 1997, 63, 2095–2099. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S. Induce systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Kilic-Ekici, O.; Yuen, G.Y. Induced resistance as a mechanism of biological control by Lysobacter enzymogenes strain C3. Phytopathology 2003, 93, 1103–1110. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wilhite, S.E.; Lunsden, R.D.; Strancy, D.C. Peptide synthetase gene in Trichoderma virens. Appl. Environ. Microbiol. 2001, 67, 5055–5062. [Google Scholar] [CrossRef] [PubMed]
- Menzler-Hokkanen, I. Socioeconomic significance of biological control. In An Ecological and Societal Approach to Biological Control; Eilenbergh, J., Hokkanen, M.T., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 13–25. [Google Scholar]
- He, P.; Chintamanani, S.; Chen, Z.; Zhu, L.; Kunkel, B.N.; Alfano, J.R.; Tang, X.; Zhou, J.M. Activation of a COI1-dependent pathway in Arabidopsis by Pseudomonas syringae type III effectors and coronatine. Plant J. 2004, 37, 589–602. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; De Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Kunz, C.; Sellam, O.; Bertheau, Y. Purification and characterization of a chitinase from the hyperparasitic fungus Aphanocladium album. Physiol. Mol. Plant Pathol. 1992, 40, 117–131. [Google Scholar] [CrossRef]
- Biali, M.; Dinoor, A.; Eshed, N.; Kenneth, R. Aphanocladium album, a fungus inducing teliospore production in rusts. Ann. Appl. Biol. 1972, 72, 37–42. [Google Scholar] [CrossRef]
- Ciccarese, F.; Longo, O.; Ambrico, A.; Schiavone, D. Aphanocladium album: Un promettente limitatore biologico dell’oidio del pomodoro e dello zucchino. La difesa delle colture in agricoltura biologica. Not. Mal. Piante 2001, 52, 69–71. [Google Scholar]
- Ciccarese, F.; Amenduni, M.; Schiavone, D.; Ambrico, A. Aphanocladium album, a new promising biocontrol agent against Oidium lycopersici. In Proceedings of the 10th Congress of the Mediterranean Phytopathological Union, Montpellier, France, 1–5 June 1997; pp. 559–562. [Google Scholar]
- Sasanelli, N.; Ciccarese, F.; Papajová, I. Aphanocladium album by via sub-irrigation in the control of Pyrenochaeta lycopersici and Meloidogyne incognita on tomato in a plastic-house. Helminthologia 2008, 45, 137–142. [Google Scholar] [CrossRef]
- Leoni, C.; Piancone, E.; Sasanelli, N.; Bruno, G.L.; Manzari, C.; Pesole, G.; Ceci, L.R.; Volpicella, M. Plant Health and Rhizosphere Microbiome: Effects of the Bionematicide Aphanocladium album in Tomato Plants Infested by Meloidogyne javanica. Microorganisms 2020, 8, 1922. [Google Scholar] [CrossRef]
- Ciccarese, F.; Longo, O.; Ambrico, A.; Schiavone, D.; Ziadi, T. Use of Aphanocladium album (isolate MX-95) in control of grape rot diseases in pre- and post-harvest [Vitis vinifera L.; Apulia]. In Atti Delle Giornate Fitopatologiche (Italy) Vol. II, Riccione, Italy, 27–29 March 2006; FAO: Rome, Italy, 2008; pp. 443–447. [Google Scholar]
- Studer, M.; Flück, K.; Zimmermann, W. Production of chitinases by Aphanocladium album grown on crystalline and colloidal chitin. FEMS Microbiol. Lett. 1992, 99, 213–216. [Google Scholar] [CrossRef]
- Sivastava, A.K.; Défago, G.; Boiler, T. Secretion of chitinase by Aphanocladium album, a hyperparasite of wheat rust. Experientia 1985, 41, 1612–1613. [Google Scholar] [CrossRef]
- Palizi, P.; Goltapeh, E.M.; Pourjam, E.; Safaie, N. Potential of Oyster mushrooms for the biocontrol of sugar beet nematode (Heterodera schachtii). J. Plant Prot. Res. 2009, 49, 27–33. [Google Scholar] [CrossRef]
- Savoie, J.M.; Mata, G.; Mamoun, M. Variability in brown line formation and extracellular laccase production during interaction between white-rot basidiomycetes and Trichoderma harzianum biotype Th2. Mycologia 2001, 93, 243–248. [Google Scholar] [CrossRef]
- Yuen, T.K.; Hyde, K.D.; Hodgkiss, I.J. Interspecific Interactions among Tropical and Subtropical Freshwater Fungi. Microb. Ecol. 1999, 37, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Morón-Ríos, A.; Gómez-Cornelio, S.; Ortega-Morales, B.O.; De la Rosa-García, S.; Partida-Martínez, L.P.; Quintana, P.; Alayón-Gamboa, A.; Cappello-García, S.; Gonzáles-Gómez, S. Interactions between abundant fungal species influence the fungal community assemblage on limestone. PLoS ONE 2017, 12, e0188443. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).