
Intense Pasture Management in Brazil in an Integrated Crop-Livestock System Simulated by the DayCent Model

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
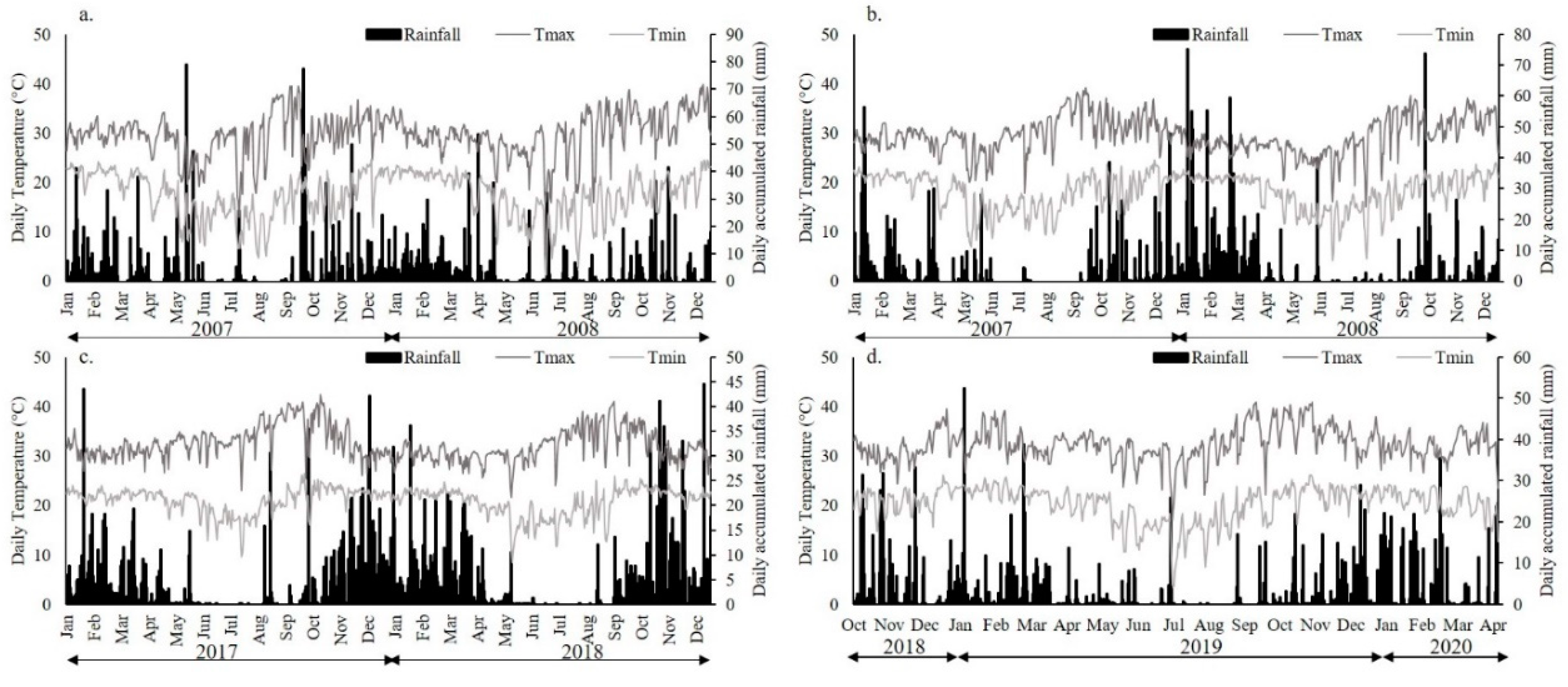
2.1. Field Data
2.2. SOC Reference Data from the Literature
2.3. Model Description and Simulation Settings
2.4. Calibration and Validation Procedures
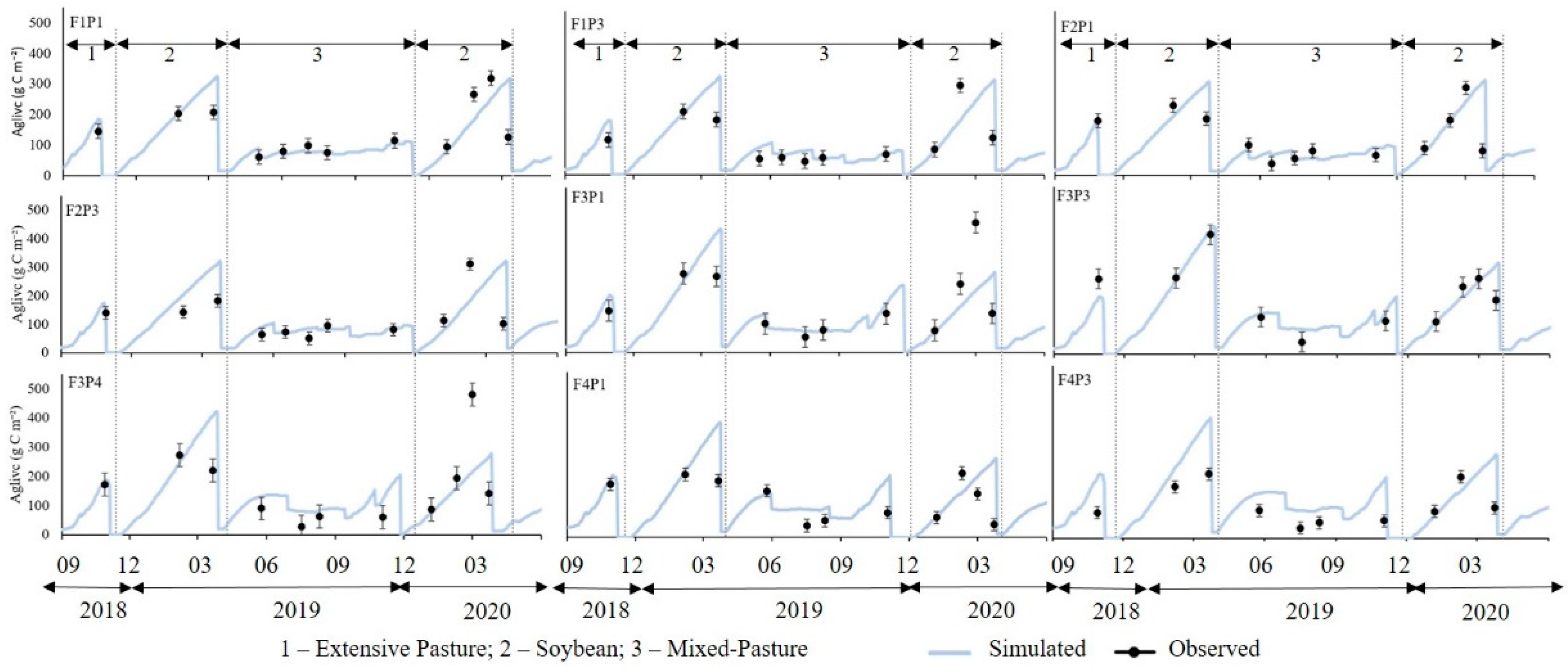
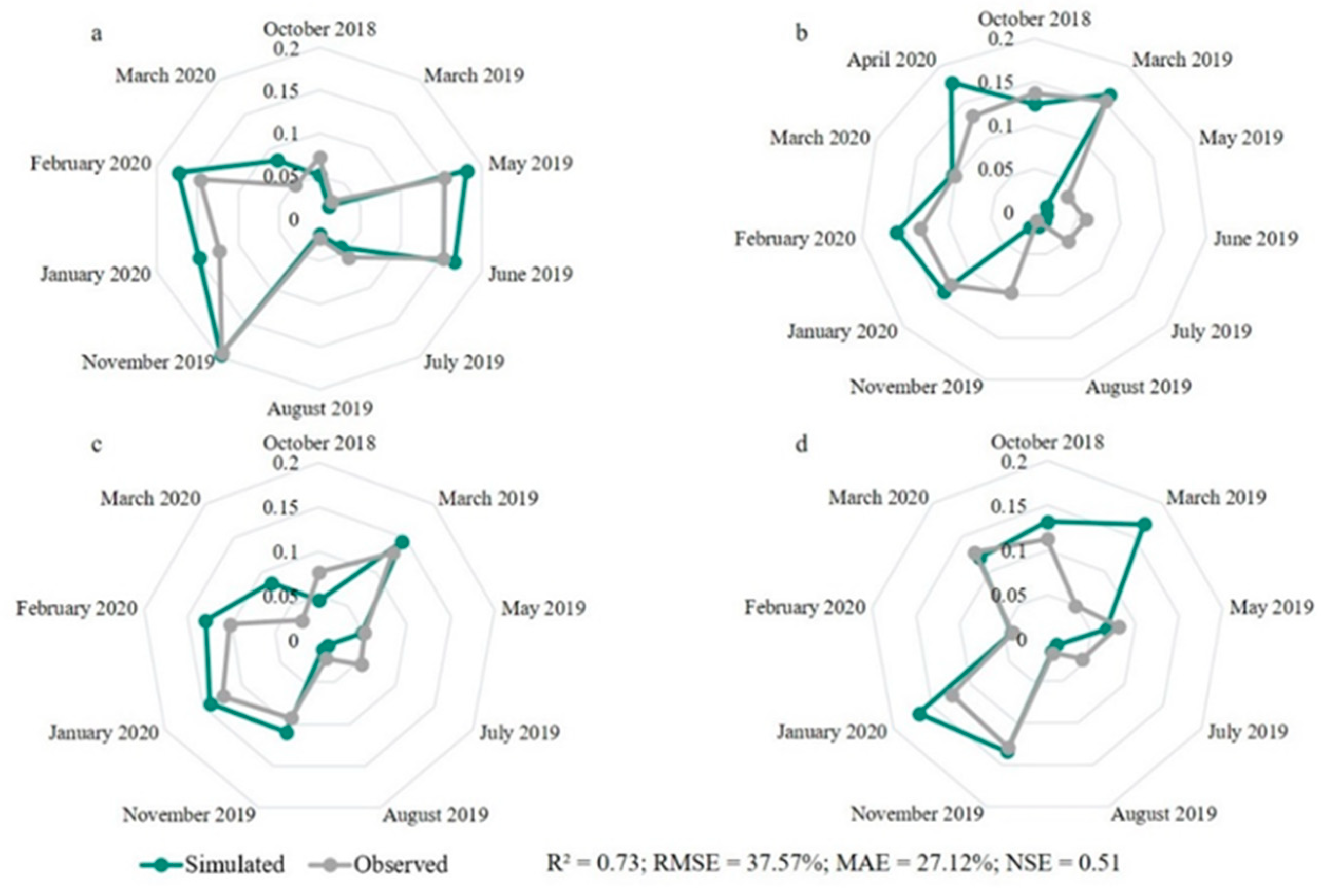
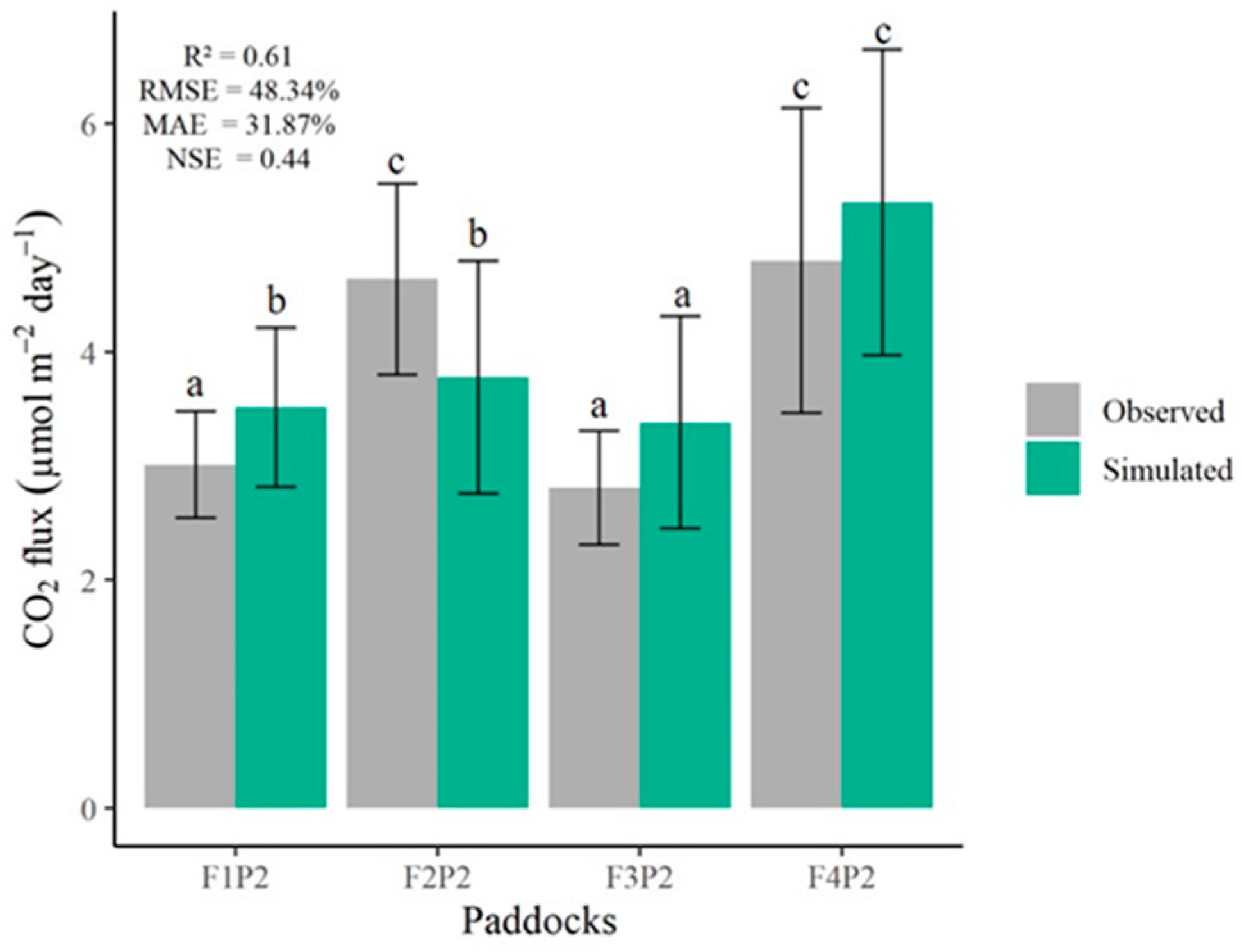
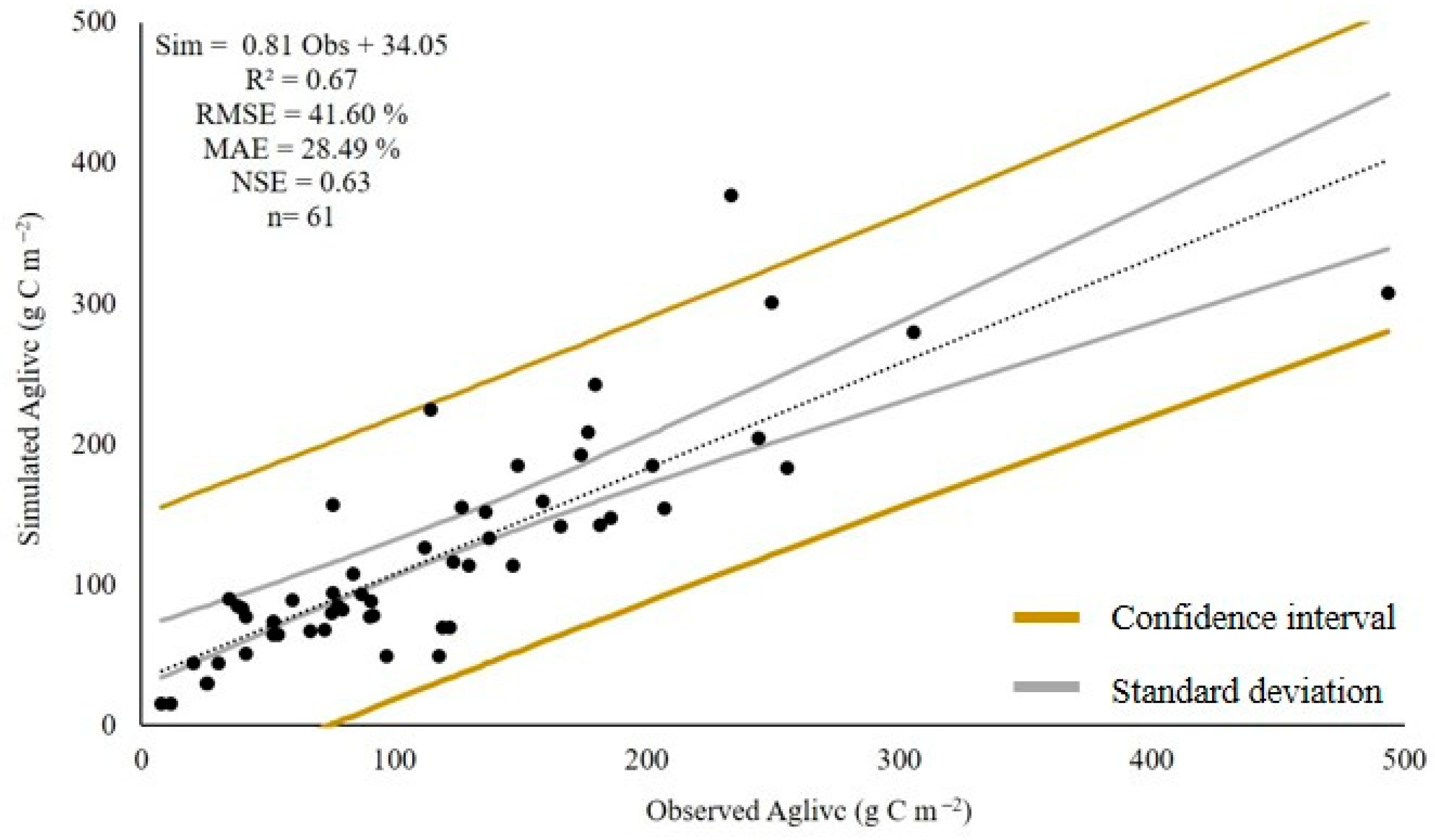
3. Results
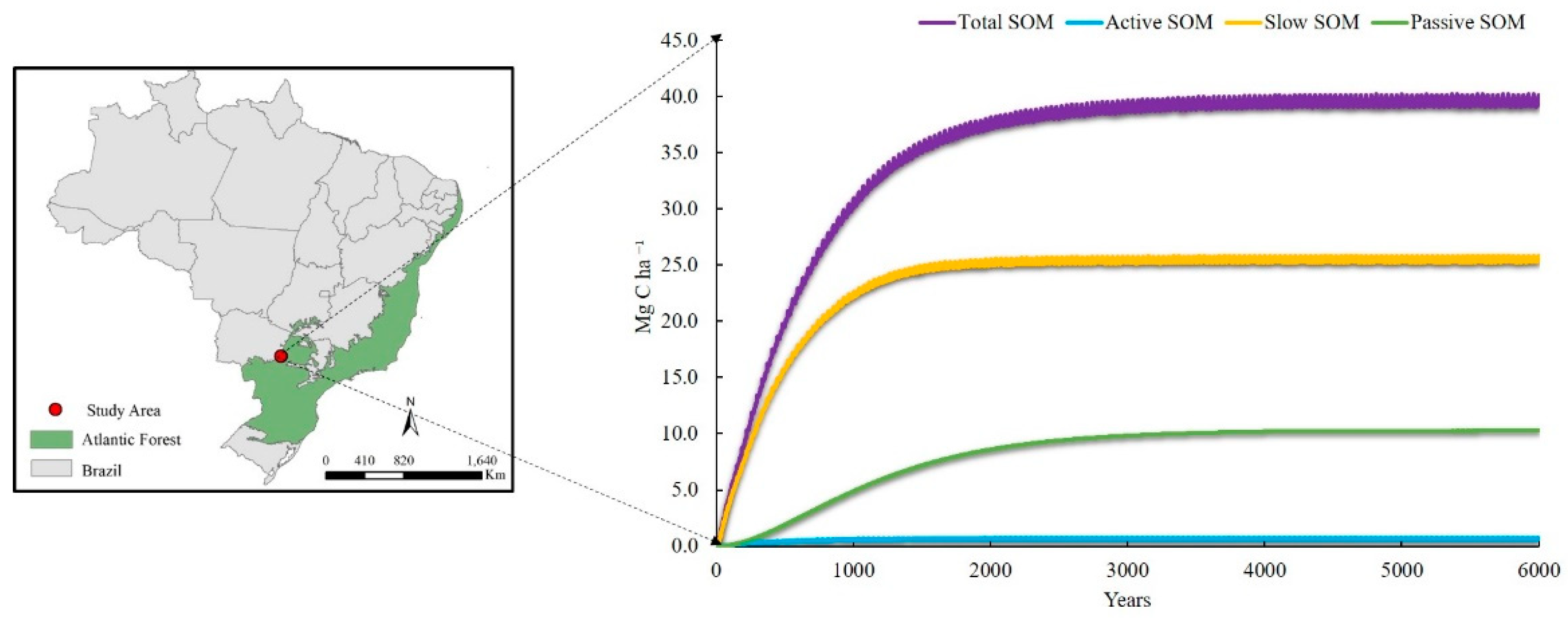
3.1. Equilibrium Modeling
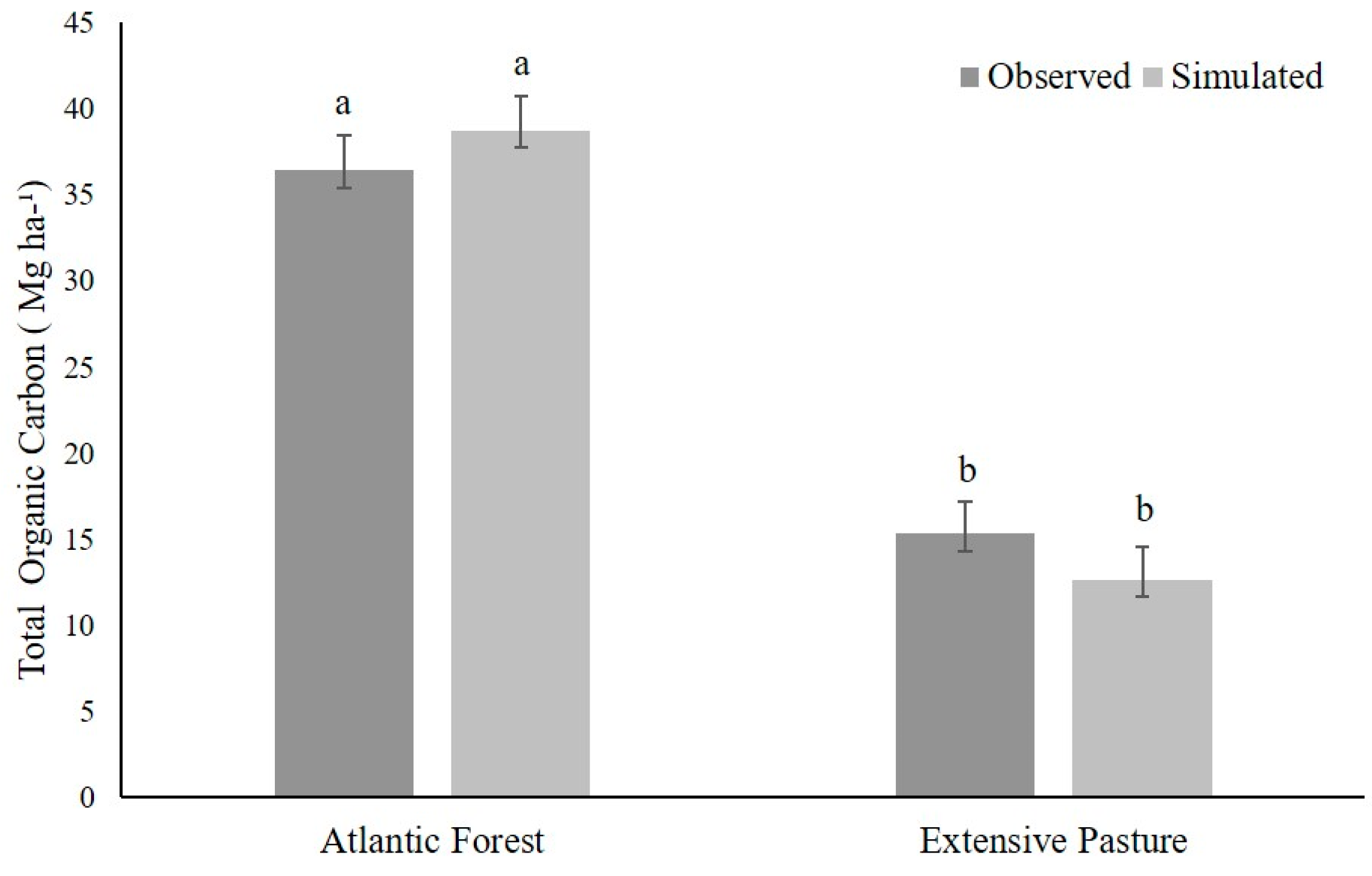
3.2. Model Calibration
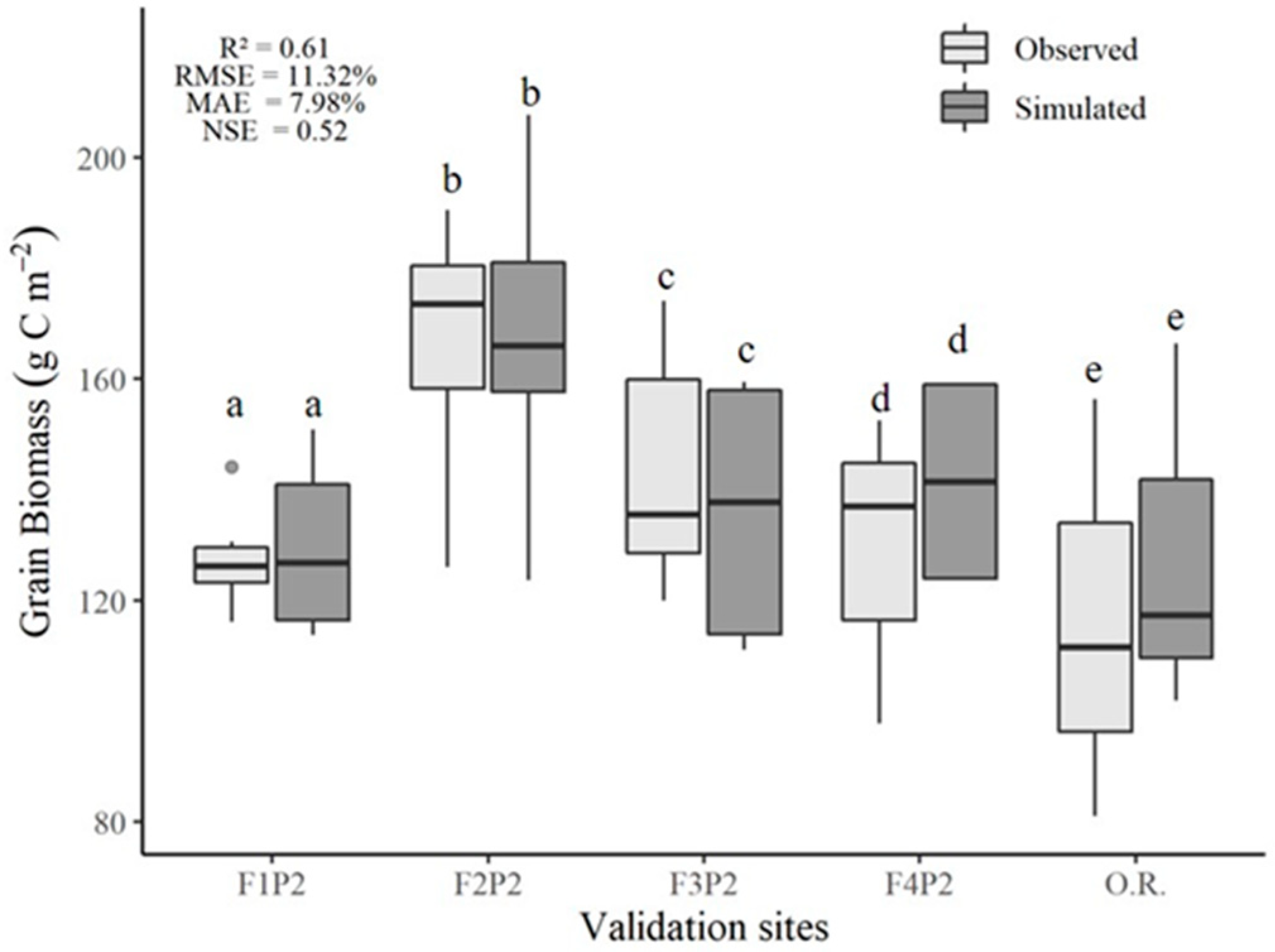
3.3. Model Validation
4. Discussion
4.1. Equilibrium
4.2. Calibrated Model
4.3. Validated Model
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season Managements (Crop Phase) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Soybean Cycle 2018/2019 | Soybean Cycle 2019/2020 | |||||||||
| Field | Pad. | Cultivar | Sowing date | Harvest date | Cultivar | Sowing date | Harvest date | |||
| 1 | 1, 2, 3 | BRS7380 RR | 18 November 2018 | 02 April 2019 | NS6700 | 12 December 2019 | 07 April 2020 | |||
| 2 | 1, 2, 3 | AS3730 IPRO | 20 November 2018 | 30 March 2019 | NS6700 | 11 December 2019 | 04 April 2020 | |||
| 3 | 1, 2, 3, 4 | NS6700 | 21 November 2018 | 04 April 2019 | AS3730 IPRO | 13 December 2019 | 08 April 2020 | |||
| 4 | 1, 2, 3 | NS6700 | 22 November 2018 | 05 April 2019 | BRS7380 RR | 15 December 2019 | 09 April 2020 | |||
| Off-season managements in 2019 (mixed-pasture phase) | ||||||||||
| Field | Pad. | Mixed-pasture | Sowing date | 1° Grazing | 2° Grazing | 3° Grazing | Events | |||
| Period | Stocking rate | Period | Stocking rate | Period | Stocking rate | Mowing | ||||
| 1 | 1 | Millet + ruzigrass | 02/04 | 17/05–26/05 | 152 | 14/07–21/07 | 59 | 17/08–28/08 | 103 | 15/06 |
| 2 | 26/05–04/06 | 21/07–28/07 | 28/08–08/09 | |||||||
| 3 | 04/06–12/06 | 28/07–03/08 | 08/09–20/09 | |||||||
| 2 | 1 | Millet + ruzigrass | 04/04 | 17/05–25/05 | 205 | 14/07–21/07 | 57 | 17/08–28/08 | 103 | 14/06 |
| 2 | 25/05–02/06 | 21/07–28/07 | 28/08–08/09 | |||||||
| 3 | 02/06–09/06 | 28/07–03/08 | 08/09–20/09 | |||||||
| 3 | 1 | Millet + ruzigrass | 05/04 | 27/05–07/06 | 146 | 17/08–26/08 | 143 | 09/10–14/10 | 121 | - |
| 2 | 07/06–18/06 | 26/08–05/09 | 14/10–19/10 | |||||||
| 3 | 18/06–29/06 | 05/09–14/09 | 19/10–24/10 | |||||||
| 4 | 29/06–11/07 | 14/09–22/09 | 24/10–31/10 | |||||||
| 4 | 1 | Millet + ruzigrass | 06/04 | 31/05–14/06 | 163 | 17/08–29/08 | 78 | - | - | - |
| 2 | 14/06–28/06 | 29/08–10/09 | ||||||||
| 3 | 28/06–11/07 | 10/09–22/09 | ||||||||
| Sampling Fields Collects Period | Land Use | System Phase | Collected Data | Sampled Fields |
|---|---|---|---|---|
| 28 October 2018 | Extensive pasture | Before ICLS | Soil samplings for determination of soil organic carbon and pH, aboveground biomass, volumetric soil water content, bulk density, total soil carbon | 1, 2, 3, 4 |
| 5 February 2019 | Soybean | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 21 March 2019 | Soybean | ICLS | Aboveground biomass, volumetric soil water content, grain biomass | 1, 2, 3, 4 |
| 18 April 2019 | Mixed-pasture | ICLS | Soil respiration | 1, 2, 3, 4 |
| 01 May 2019 | Mixed-pasture | ICLS | Soil respiration | 1, 2, 3, 4 |
| 11 May 2019 | Mixed-pasture | ICLS | Soil respiration | 1, 2, 3, 4 |
| 17 May 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 1 and 2 |
| 25 May 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 3 and 4 |
| 16 June 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 1 and 2 |
| 17 July 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 10 August 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 2 November 2019 | Mixed-pasture | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 8 November 2019 | Mixed-pasture | ICLS | Soil respiration | 1, 2, 3 |
| 15 November 2019 | Mixed-pasture | ICLS | Soil respiration | 1, 2, 3, 4 |
| 6 January 2020 | Soybean | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 8 January 2020 | Soybean | ICLS | Soil respiration | 1, 2, 3, 4 |
| 9 February 2020 | Soybean | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 14 February 2020 | Soybean | ICLS | Soil respiration | 1, 2, 3, 4 |
| 1 March 2020 | Soybean | ICLS | Aboveground biomass, volumetric soil water content | 1, 2, 3, 4 |
| 1 March 2020 | Soybean | ICLS | Soil respiration | 1, 2, 3 |
| 23 March 2020 | Soybean | ICLS | Aboveground biomass, volumetric soil water content, grain biomass | 1, 2, 3, 4 |
| 23 March 2020 | Soybean | ICLS | Soil respiration | 1, 2, 3, 4 |
| Initials | Descriptions |
|---|---|
| AGLREM | Fraction of aboveground live which will not be affected by harvest operations |
| BGLREM | Fraction of belowground live which will not be affected by harvest operations |
| BIOMAX | Aboveground biomass level above which the minimum and maximum C/E ratios of new shoot increments equal pramn(*,2) and pramx(*,2), respectively |
| CLAYPG | Number of soil layers that crop roots can occupy. The value used as CLAYPG for annual plants will vary from 1 on the day that plant growth starts to CLAYPG as read from the CROP option on day FRTC(3) of plant growth |
| EFRGRN(1) | Fraction of aboveground N which goes to grain |
| FDGREM | Fraction of standing dead (stdedc) removed by a grazing event over a one-month period. The daily removal rate is approximately fdgrem/30 |
| FERAMT(1) | Amount of N to be added |
| FERAMT(2) | Amount of P to be added |
| FLGREM | Fraction of live shoots (aglivc) removed by a grazing event over a one-month period. The daily removal rate is approximately flgrem/30 |
| FULCAN | Value of aboveground live C (aglivc) at full canopy cover, above which potential production is not reduced (above which there is no restriction on seedling growth) |
| PPDF(3) | Left curve shape for parameterization of a Poisson Density Function curve to simulate temperature effect on growth |
| PPDF(4) | Right curve shape for parameterization of a Poisson Density Function curve to simulate temperature effect on growth |
| PRAMN(1,1) | Minimum aboveground C/N ratio with zero biomass |
| PRAMN(1,2) | Minimum aboveground C/N ratio with biomass > biomax |
| PRAMX(1,1) | Maximum aboveground C/N ratio with zero biomass |
| PRAMX(1,2) | Maximum aboveground C/N ratio with biomass > biomax |
| PRDX(1) | Coefficient for calculating total monthly potential production as a function of solar radiation outside the atmosphere. It functions as a radiation use efficiency scalar on potential production. It reflects the relative genetic potential of the plant; larger PRDX(1) values indicate greater growth potential |
| PRDX(2) | Coefficient for calculating total monthly potential production as a function of solar radiation outside the atmosphere. It functions as a radiation use efficiency scalar on potential production. It reflects the relative genetic potential of the plant; larger PRDX(2) values indicate greater growth potential |
| SNFXMX | Maximum symbiotic N fixation for forest (actual symbiotic N fixation will be less if available mineral N is sufficient for growth) |
References
- Moraes, A.; Carvalho, P.C.F.; Anghinoni, I.; Lustosa, S.B.C.; Costa, S.E.V.G.A.; Kunrath, T.R. Integrated crop—Livestock systems in the Brazilian subtropics. Eur. J. Agron. 2014, 57, 4–9. [Google Scholar] [CrossRef]
- Embrapa. ILPF em Número, 12. 2016. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/158636/1/2016-cpamt-ilpf-em-numeros.pdf (accessed on 1 August 2021). (In Portuguese).
- Bonaudo, T.; Bendahan, A.B.; Sabatier, R.; Ryschawy, J.; Bellon, S.; Leger, F.; Magda, D.; Tichit, M. Agroecological principles for the redesign of integrated crop-livestock systems. Eur. J. Agron. 2014, 57, 43–51. [Google Scholar] [CrossRef]
- Wruck, F.J.; Behling, M.; Antonio, D.B.A. Sistemas Integrados em Mato Grosso e Goiás. In Sistemas Agroflorestais: A Agropecuária Sustentável, 1st ed.; Alves, F.V., Laura, V.A., Almeida, R.G., Eds.; Embrapa: Brasilia, Brazil, 2015; pp. 169–194. (In Portuguese) [Google Scholar]
- Silva, L.S.; Laroca, J.V.S.; Coelho, A.P.; Gonçalves, E.C.; Gomes, R.P.; Pacheco, L.P.; Carvalho, P.C.F.; Pires, G.C.; Oliveira, R.L.; Souza, J.M.A.; et al. Does grass-legume intercropping change soil quality and grain yield in integrated crop-livestock systems? Appl. Soil Ecol. 2022, 170, 104257. [Google Scholar] [CrossRef]
- Jones, J.W.; Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Batchelor, W.D.; Hunt, L.A.; Wilkens, P.W.; Singh, U.; Gijsman, A.J.; Ritchie, J.T. The DSSAT cropping system model. Eur. J. Agron. 2003, 18, 3–4. [Google Scholar] [CrossRef]
- Zhang, Y.; Hansen, N.; Trout, T.; Nielsen, D.; Paustian, K. Modeling Deficit Irrigation of Maize with the DayCent Model. Agron. J. 2018, 110, 1754–1764. [Google Scholar] [CrossRef]
- Nehbandani, A.; Soltani, A.; Hajjarpoor, A.; Dadrasi, A.; Nourbakhsh, F. Comprehensive yield gap analysis and optimizing agronomy practices of soybean in Iran. J. Agric. Sci. 2020, 158, 739–747. [Google Scholar] [CrossRef]
- Zhang, Y.; Gurung, R.; Marx, E.; Williams, S.; Ogle, S.M.; Paustian, K. DayCent model predictions of NPP and grain yields for agricultural lands in the contiguous U.S. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005750. [Google Scholar] [CrossRef]
- Lemma, B.; Williams, S.; Paustian, K. Long term soil carbon sequestration potential of smallholder croplands in southern Ethiopia with Daycent model. J. Environ. Manag. 2021, 294, 112893. [Google Scholar] [CrossRef]
- Necpálová, M.; Anex, R.P.; Fienen, M.N.; Del Grosso, S.J.; Castellano, M.J.; Sawyer, J.E.; Iqbal, J.; Pantoja, J.L.; Barker, D.W. Understanding the Daycent model: Calibration, sensitivity, and identifiability through inverse modeling. Environ. Model. Softw. 2015, 66, 110–130. [Google Scholar] [CrossRef] [Green Version]
- Frolking, S.E.; Moiser, A.R.; Ojima, D.S.; Li, C.; Parton, W.J.; Potter, C.S.; Priesack, E.; Stenger, R.; Haberbosch, C.; Dorsch, P. Comparison of N2O Emissions from Soils at Three Temperate Agricultural Sites: Simulations of year-round measurements by four models. Nutr. Cycl. Agroecosyst. 1998, 52, 77–105. [Google Scholar] [CrossRef]
- Del Grosso, S.J.; Mosier, A.R.; Parton, W.J.; Ojima, D.S. Daycent model analysis of past and contemporary soil N2O and net greenhouse gas flux for major crops in the USA. Soil Tillage Res. 2005, 83, 9–24. [Google Scholar] [CrossRef]
- Parton, W.J.; Hartman, M.; Ojima, D.; Schimel, D. Daycent description and testing. Glob. Planet. Chang. 1998, 19, 35–48. [Google Scholar] [CrossRef]
- Weiler, D.A.; Tornquist, C.G.; Parton, W.; Santos, H.P.; Santi, A.; Bayer, C. Crop Biomass, Soil Carbon, and Nitrous Oxide as Affected by Management and Climate: A Daycent Application in Brazil. Soil Sci. Soc. Am. J. 2017, 81, 945–955. [Google Scholar] [CrossRef]
- Damian, J.M.; Matos, E.S.; Pedreira, B.C.; Carvalho, P.C.F.; Premazzi, L.M.; Williams, S.; Paustian, K.; Cerri, C.E.P. Predicting soil C changes after pasture intensification and diversification in Brazil. Catena 2021, 202, 105238. [Google Scholar] [CrossRef]
- Laroca, J.V.S.; Souza, J.M.A.; Pires, G.C.; Pires, G.J.C.; Pacheco, L.P.; Silva, F.D.; Wruck, F.J.; Carneiro, M.A.C.; Silva, L.S.; Souza, E.D. Soil quality and soybean productivity in crop-livestock integrated system in no-tillage. Pesqui. Agropecu. Bras. 2018, 53, 1248–1258. [Google Scholar] [CrossRef]
- FAO. Measuring and Modelling Soil Carbon Stocks and Stock Changes in Livestock Production Systems. 2018. Available online: http://www.fao.org/3/I9693EN/i9693en.pdf (accessed on 15 September 2021).
- Fitton, N.; Bindi, M.; Brilli, L.; Cichota, R.; Dibari, C.; Fuchs, K.; Huguenin-elie, O.; Klumpp, K.; Lieffering, M.; Lüscher, A.; et al. Modelling biological N fixation and grass-legume dynamics with process-based biogeochemical models of varying complexity. Eur. J. Agron. 2019, 106, 58–66. [Google Scholar] [CrossRef]
- Baslam, M.; Mitsui, T.; Hodges, M.; Priesack, E.; Herritt, M.T.; Aranjuelo, I.; Sanz-Sáez, Á. Photosynthesis in a changing global climate: Scaling up and scaling down in crops. Front. Plant Sci. 2020, 11, 882. [Google Scholar] [CrossRef]
- Keohane, N.; Petsonk, A.; Hanafi, A. Toward a club of carbon markets. Clim. Chang. 2015, 144, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Del Grosso, S.J.; Parton, W.J.; Adler, P.R.; Davis, S.C.; Keough, C.; Marx, E. Daycent model simulations for estimating soil carbon dynamics and greenhouse gas fluxes from agricultural production systems. In Managing Agricultural Greenhouse Gases; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 241–250. [Google Scholar]
- Hartman, M.; Parton, W.; Del Grosso, S.; Easter, M.; Hendryx, J.; Hilinski, T.; Kelly, R.; Keough, C.; Killian, K.; Lutz, S.; et al. The Daily Century Ecosystem, Soil Organic Matter, Nutrient Cycling, Nitrogen Trace Gas, and Methane Model: User Manual, Scientific Basis, and Technical Documentation. In Natural Resource Ecology Laboratory; Colorado State University: Fort Collins, CO, USA, 2018. [Google Scholar]
- United Nations General Assembly. Transforming Our World: The 2030 Agenda for Sustainable Development. 2015. Available online: http://www.un.org/ga/search/view_doc.asp?symbol=A/RES/70/1&Lang=E (accessed on 20 January 2022).
- Gil, J.D.B.; Reidsma, P.; Giller, K.; Todman, L.; Whitmore, A.; Van Ittersum, M. Sustainable development goal 2: Improved targets and indicators for agriculture and food security. Ambio 2019, 48, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.M.; Hansen, J.; Rioux, J.; Stirling, C.M.; Twomlow, S.; Wollenberg, E.L. Urgent action to combat climate change and its impacts (SDG 13): Transforming agriculture and food systems. Curr. Opin. Environ. Sustain. 2018, 34, 13–20. [Google Scholar] [CrossRef]
- UN. SDG-15; Life on Land: Why It Matters. United Nations Sustainable Development Goals: New York, NY, USA, 2016.
- Rolim, G.S.; Camargo, M.B.P.; Grosselilania, D.; Moraes, J.F.L. Climatic classification of Köppen and Thornthwaite sistems and their applicability in the determination of agroclimatic zonning for the state of São Paulo, Brazil. Bragantia 2007, 66, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Battisti, R.; Bender, F.D.; Sentelhas, P.C. Assessment of different gridded weather data for soybean yield simulations in Brazil. Theor. Appl. Climatol. 2019, 135, 237–247. [Google Scholar] [CrossRef]
- Dias, H.B.; Sentelhas, P.C. Assessing the performance of two gridded weather data for sugarcane crop simulations with a process-based model in Center-South Brazil. Int. J. Biometeorol. 2021, 65, 1881–1893. [Google Scholar] [CrossRef]
- Bender, F.D.; Sentelhas, P.C. Solar Radiation Models and Gridded Databases to Fill Gaps in Weather Series and to Project Climate Change in Brazil. Adv. Meteorol. 2018, 2018, 6204382. [Google Scholar] [CrossRef] [Green Version]
- Folfaro, V.J.; Neto, F.S. Ensaio de caracterização estratigrafica do cretaceo no estado de São Paulo: Grupo bauru. Rev. Bras. Geociênc 1976, 10, 177–185. (In Portuguese) [Google Scholar]
- Moniz, A.C.; Carvalho, A. Sequência de evolução de solos derivados do arenito Bauru e de rochas básicas da região noroeste do estado de São Paulo. Bragantia 1973, 32, 309–335. (In Portuguese) [Google Scholar] [CrossRef]
- Rossi, M. Mapa Pedológico do Estado de São Paulo: Revisado e Ampliado; Instituto Florestal: Sao Paulo, Brazil, 2017. (In Portuguese)
- Schoeneberger, P.; Wysocki, D.; Busskohl, C.; Libohova, Z. Landscapes, Geomorphology, and Site Description; USDA Natural Resources Conservation Service Soils: Waverley, IA, USA, 2017; Volume 18, pp. 21–80.
- Esquerdo, J.C.D.M.; Antunes, J.F.G.; Coutinho, A.C.; Speranza, E.A.; Kondo, A.A.; Santos, J.L. SATVeg: A web-based tool for visualization of MODIS vegetation indices in South America. Comput. Electron. Agric. 2020, 175, 105516. [Google Scholar] [CrossRef]
- Sato, A.; Tsuyuzaki, T.; Seto, M. Effect of Soil Agitation, Temperature or Moisture on Microbial Biomass Carbon of a Forest and an Arable Soil. Microbes Environ. 2000, 15, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Embrapa. Manual de Métodos de Análise de Solo; Embrapa CNPS: Rio de Janeiro, Brazil, 1997. (In Portuguese) [Google Scholar]
- FAO. Knowledge Reference for National Forest Assessments—Modeling for Estimation and Monitoring. 2005. Available online: http://www.fao.org/forestry/17111/en/ (accessed on 15 September 2021).
- Rigolin, I.M.; Santos, C.H.; Calonego, J.C.; Tiritan, C.S. Estoque De Carbono Do Solo Em Sistemas Vegetais Com Manejo Agrícola Diferenciado No Oeste Paulista. Colloq. Agrar. 2013, 9, 16–29. (In Portuguese) [Google Scholar] [CrossRef]
- Souza, M.C. Consorciação de braquiária, milheto e crotalária em safrinha na produção de fitomassa e cobertura do solo. In 35 f. Trabalho de Conclusão de Curso (Graduação em Engenharia Agrícola e Ambiental); Universidade Federal de Mato Grosso, Instituto de Ciências Agrárias e Tecnológicas: Rondonopolis, Brazil, 2018. (In Portuguese) [Google Scholar]
- Machado, L.A.Z. Misturas de forrageiras anuais e perenes para sucessão à soja em sistemas de integração lavoura pecuária. Pesqui. Agropecu. Bras. 2012, 47, 629–636. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Del Grosso, S.J.; Parton, W.J.; Mosier, A.R.; Hartman, M.D.; Brenner, J.; Ojima, D.S.; Schimel, D.S. Simulated interaction of carbon dynamics and nitrogen trace gas fluxes using the Daycent model. In Modeling Carbon and Nitrogen Dynamics for Soil Management; Schaffer, M., Ma, L., Hansen, S., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 303–332. [Google Scholar]
- Parton, W.J.; Schimel, D.S.; Cole, C.V.; Ojima, D.S. Analysis of factors controlling soil organic levels of grasslands in the Great Plains. Soil Sci. Soc. Am. J. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Parton, W.J.; Stewart, J.W.B.; Cole, C.V. Dynamics of C, N, P and S in grassland soils: A model. Biogeochemistry 1988, 5, 109–131. [Google Scholar] [CrossRef]
- Wieder, W.R.; Hartman, M.D.; Sulman, B.N.; Wang, Y.-P.; Koven, C.D.; Bonan, G.B. Carbon cycle confidence and uncertainty: Exploring variation among soil biogeochemical models. Glob. Chang. Biol. 2018, 24, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating generalized soil-water characteristics from texture. Soil Sci. Soc. Am. J. 1986, 50, 1031–1036. [Google Scholar] [CrossRef]
- Garrison, M.V.; Batchelor, W.D.; Kanwar, R.S.; Ritchie, J.T. Evaluation of the CERES-Maize water and nitrogen balances under tile-drained conditions. Agric. Syst. 1999, 62, 189–200. [Google Scholar] [CrossRef]
- Chen, L.; Wang, K.; Baoyin, T. Effects of grazing and mowing on vertical distribution of soil nutrients and their stoichiometry (C:N:P) in a semi-arid grassland of North China. Catena 2021, 206, 105507. [Google Scholar] [CrossRef]
- Gilmullina, A.; Rumpel, C.; Blagodatskaya, E.; Chabbi, A. Management of grasslands by mowing versus grazing—Impacts on soil organic matter quality and microbial functioning. Appl. Soil Ecol. 2020, 156, 103701. [Google Scholar] [CrossRef]
- Heggenstaller, A.H.; Moore, K.J.; Liebman, M.; Anex, R.P. Nitrogen influences biomass and nutrient partitioning by perennial, warm-season grasses. Agron. J. 2009, 101, 1363–1371. [Google Scholar] [CrossRef]
- Sainju, U.M.; Allen, B.L.; Lenssen, A.W.; Ghimire, R.P. Root biomass, root/shoot ratio, and soil water content under perennial grasses with different nitrogen rates. Field Crop. Res. 2017, 210, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wang, Q.; Hong, J.; Chen, W.; Qi, C.; Ye, L. Life cycle assessment of diammonium-and monoammonium-phosphate fertilizer production in China. J. Clean. Prod. 2017, 141, 1087–1094. [Google Scholar] [CrossRef]
- Anghinoni, G.; Anghinoni, F.B.G.; Tormena, C.A.; Braccini, A.L.; Mendes, I.C.; Zancanaro, L.; Lal, R. Conservation agriculture strengthen sustainability of Brazilian grain production and food security. Land Use Policy 2021, 108, 105591. [Google Scholar] [CrossRef]
- Soares, D.S.; Ramos, M.L.G.; Marchão, R.L.; Maciel, G.A.; Oliveira, A.D.; Malaquias, J.V.; Carvalho, A.M. How diversity of crop residues in long-term no-tillage systems affect chemical and microbiological soil properties. Soil Tillage Res. 2019, 194, 104316. [Google Scholar] [CrossRef]
- Nascente, A.S.; Li, Y.C.; Crusciol, C.A.C. Cover crops and no-till effects on physical fractions of soil organic matter. Soil Tillage Res. 2013, 130, 52–57. [Google Scholar] [CrossRef]
- Teixeira, R.A.; Soares, T.G.; Fernandes, A.R.; Braz, A.M.S. Grasses and legumes as cover crop in no-tillage system in northeastern Pará Brazil. Acta Amaz. 2014, 44, 411–418. [Google Scholar] [CrossRef]
- Yang, J.M.; Yang, J.Y.; Liu, S.; Hoogenboom, G. An evaluation of the statistical methods for testing the performance of crop models with observed data. Agric. Syst. 2014, 127, 81–89. [Google Scholar] [CrossRef]
- Priesack, E.; Gayler, S.; Hartmann, H. The impact of crop growth sub-model choice on simulated water and nitrogen balances. Nutr. Cycl. Agroecosyst. 2006, 75, 1–13. [Google Scholar] [CrossRef]
- Moriasi, D.N.; Arnold, J.G.; Liew, M.W.V.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. ASABE 2007, 50, 885–900. [Google Scholar] [CrossRef]
- Ihaka, R. Gentleman R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.C.G.; Foloni, J.S.S.; Fabris, L.B.; Tiritan, C.S. Fitomassa e relação C/N em consórcios de sorgo e milho com espécies de cobertura. Pesqui. Agropecu. Bras. 2009, 44, 1504–1512. (In Portuguese) [Google Scholar] [CrossRef]
- Ferreira, A.O.; Moraes Sá, J.C.; Harms, M.G.; Miara, S.; Briedis, C.; Netto, C.Q.; Santos, J.B.; Canalli, L.B. Carbon balance and crop residue management in dynamic equilibrium under a no-till system in campos gerais. Rev. Bras. Ciência Solo 2012, 36, 1583–1590. [Google Scholar] [CrossRef] [Green Version]
- Mazurana, M.; Fink, J.R.; Camargo, E.; Schmitt, C.; Andreazza, R.; Camargo, F.A.O. Estoque de carbono e atividade microbiana em sistema de plantio direto consolidado no Sul do Brasil. Rev. Ciências Agrárias 2013, 36, 288–296. (In Portuguese) [Google Scholar]
- Rangel, O.J.P.; Silva, C.A.; Guimarães, P.T.G.; Melo, L.C.A.; Oliveira Junior, A.C. Carbono orgânico e nitrogênio total do solo e suas relações com os espaçamentos de plantio de cafeeiro. Rev. Bras. Ciência Solo 2008, 32, 2051–2059. (In Portuguese) [Google Scholar] [CrossRef]
- Bortolon, E.S.O.; Mielniczuk, J.; Tornquist, C.G.; Lopes, F.; Bergamaschi, H. Validation of the Century model to estimate the impact of agriculture on soil organic carbon in Southern Brazil. Geoderma 2011, 167, 156–166. [Google Scholar] [CrossRef]
- Sant-Anna, S.A.C.; Jantalia, C.P.; Vilela, J.M.S.L.; Marchão, R.L.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M. Changes in soil organic carbon during 22 years of pastures, cropping or integrated crop/livestock systems in the Brazilian Cerrado. Nutr. Cycl. Agroecosyst. 2016, 180, 101–120. [Google Scholar] [CrossRef] [Green Version]
- Sarker, J.R.; Singh, B.P.; Warwick, J.; Dougherty, W.J.; Fang, Y.; Badgery, W.; Hoyle, F.C.; Dalal, R.C.; Cowie, A.L. Impact of agricultural management practices on the nutrient supply potential of soil organic matter under long-term farming systems. Soil Tillage Res. 2018, 175, 71–81. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Moinet, M.; Hunt, J.E.; Rumpel, C.; Chabbi, A.; Millard, P. Temperature sensitivity of decomposition decreases with increasing soil organic matter stability. Sci. Total Environ. 2020, 704, 135460. [Google Scholar] [CrossRef]
- Tiecher, T. Manejo e Conservação do solo e da água em Pequenas Propriedades Rurais No sul do Brasil: Práticas Alternativas de Manejo Visando a Conservação do solo e da Água; UFRGS: Porto Alegre, Brazil, 2016; 186p. (In Portuguese) [Google Scholar]
- Jarecki, M.; Kariyapperuma, K.; Deen, B.; Graham, J.; Bazrgar, A.B.; Vijayakumar, S.; Thimmanagari, M.; Gordon, A.; Voroney, P.; Thevathasan, N. The potential of Switchgrass and Miscanthus to Enhance Soil Organic Carbon Sequestration—Predicted by Daycent Model. Land 2020, 9, 509. [Google Scholar] [CrossRef]
- McClelland, S.C.; Paustian, K.; Williams, S.; Meagan, E.; Schipanski, M.E. Modeling cover crop biomass production and related emissions to improve farm-scale decision-support tools. Agric. Syst. 2021, 191, 103151. [Google Scholar] [CrossRef]
- Salton, J.C.; Mielniczuk, J.; Bayer, C.; Fabrício, A.C.; Macedo, M.C.M.; Broch, D.L. Teor e dinâmica do carbono no solo em sistemas de integração lavoura-pecuária. Pesqui. Agropecu. Bras. 2011, 46, 1349–1356. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Martin, G.; Durand, J.L.; Duru, M.; Gastal, F.; Julier, B.; Litrico, I.; Louarn, G.; Médiène, S.; Moreau, D.; Valentin-Morison, M.; et al. Role of ley pastures in tomorrow’s cropping systems. A review. Agron. Sustain. Dev. 2020, 40, 17. [Google Scholar] [CrossRef]
- Priori, S.; Pellegrini, S.; Vignozzi, N.; Costantini, E.A.C. Soil Physical-Hydrological Degradation in the Root-Zone of Tree Crops: Problems and Solutions. Agronomy 2021, 11, 68. [Google Scholar] [CrossRef]
- Wiesner, S.; Duff, A.J.; Desai, A.R.; Panke-Buisse, K. Increasing Dairy Sustainability with Integrated Crop-Livestock Farming. Sustainability 2020, 12, 765. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.E.; Johnson, J.M.F.; Jin, V.L.; Lehman, M.; Osborne, S.L.; Varvel, G.E.; Paustian, K. Assessing the soil carbon, biomass production and nitrous oxide emission impact of corn stover management for bioenergy feedstock production using Daycent. BioEnergy Res. 2014, 7, 491–502. [Google Scholar] [CrossRef]
- Weiler, D.A.; Tornquist, C.G.; Szchomack, T.; Ogle, S.M.; Carlos, F.S.; Bayer, C. Daycent simulation of methane emissions, grain yield, and soil organic carbon in a subtropical paddy rice system. Rev. Bras. Cienc. Solo 2018, 42, 1–12. [Google Scholar] [CrossRef]
- Qin, X.; Wang, H.; He, Y.K.; Li, Y.; Li, Z.; Gao, Q.; Wan, Y.; Qian, B.; McConkey, B.; DePauw, R.; et al. Simulated adaptation strategies for spring wheat to climate change in a northern high latitude environment by DAYCENT model. Eur. J. Agron. 2018, 95, 45–56. [Google Scholar] [CrossRef]
- Boote, K.J.; Jones, J.W.; Hoogenboom, G.; Pickering, N.B. The CROPGRO Model for Grain Legumes. In Understanding Options for Agricultural Production; Tsuji, G., Hoogenboom, G., Thornton, P.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 99–128. [Google Scholar]
- Lai, L.; Kumar, S.; Chintala, R.; Owens, V.N.; Clay, D.; Schumacher, J.; Nizami, A.S.; Lee, S.S.; Rafique, R. Modeling the impacts of temperature and precipitation changes on soil CO2 fluxes from a Switchgrass stand recently converted from cropland. J. Environ. Sci. 2016, 43, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Senapati, N.; Chabbi, A.; Giostri, A.F.; Yeluripati, J.B.; Smith, P. Modelling nitrous oxide emissions from mown-grass and grain-cropping systems: Testing and sensitivity analysis of DailyDayCent using high frequency measurements. Sci. Total Environ. 2016, 572, 955–977. [Google Scholar] [CrossRef] [Green Version]
- Scheer, C.; Del Grosso, S.J.; Parton, W.J.; Rowlings, D.W.; Grace, P.R. Modeling nitrous oxide emissions from irrigated agriculture: Testing Daycent with high-frequency measurements. Ecol. Appl. 2014, 24, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Bianco, L.F.; Trevizan, F.H.; Nicolino Filho, C.J.; Oliveira, T.B.M.; Neiverth, W.; Crusiol, L.G.T.; Rio, A.; Sibaldelli, R.N.R.; Carvalho, J.F.C.; Ferreira, L.C.; et al. Algumas características das cultivares de soja Embrapa 48 e BR 16 em diferentes regimes hídricos. In VIII Jornada Acadêmica da Embrapa Soja; Embrapa Soja, Documentos, 339; Resumos Expandidos: Londrina, Brazil, 2013; Volume 8, pp. 137–141. (In Portuguese) [Google Scholar]
- La Scala, N.; Lopes, A.; Spokas, K.; Bolonhezi, D.; Archer, D.W.; Reicosky, D.C. Short-term temporal changes of soil carbon losses after tillage described by a first-order decay model. Soil Tillage Res. 2008, 99, 108–118. [Google Scholar]
- Schenato, R.B. Simulação de Fluxos de Gases de Efeito Estufa em Sistemas de Manejo do solo no Sul do Brasil; Lume: Porto Alegre, Brazil, 2013; Volume 139. (In Portuguese) [Google Scholar]
- Phillips, C.L.; Bond-Lamberty, B.; Desai, A.R.; Lavoie, M.; Risk, D.; Tang, J.; Todd-Brown, K.; Vargas, R. The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling. Plant Soil 2017, 413, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.M.S.; Williams, S.; Cerri, C.E.P.; Paustian, K. Predicting soil C changes over sugarcane expansion in Brazil using the Daycent model. GCB Bioenergy 2017, 9, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Del Grosso, S.J.; Parton, W.J.; Ojima, D.S.; Keough, C.A.; Riley, T.H.; Mosier, A.R. Daycent simulated effects of land use and climate on county level N loss vectors in the USA. Nitrogen Environ. 2008, 8, 571–595. [Google Scholar]
- Piva, J.T.; Dieckow, J.; Bayer, C.; Pergher, M.; Alburquerque, M.A.; Moraes, A.; Pauletti, V. No-tillage and crop-livestock with silage production impact little on carbon and nitrogen in the short-term in a subtropical Ferralsol. Rev. Bras. Ciências Agrárias 2020, 15, 7057. [Google Scholar] [CrossRef]
- Stehfest, E.; Heistermann, M.; Priess, J.A.; Ojima, D.S.; Alcamo, J. Simulation of global crop production with the ecosystem model Daycent. Ecol. Model. 2007, 209, 203–219. [Google Scholar] [CrossRef]
- Cordeiro, L.A.M.; Kluthcouski, J.; Silva, J.R.; Rojas, D.C.; Omote, H.S.G.; Moro, E.; Silva, P.C.G.; Tiritan, C.S.; Longen, A. Integração Lavoura-Pecuária em Solos Arenosos: Estudo de caso da Fazenda Campina No Oeste Paulista; Embrapa Cerrados-Documentos: Planaltina, Brazil, 2020. (In Portuguese) [Google Scholar]
- Lee, J.; Pedroso, G.; Linquist, B.A.; Putnam, D.; Van Kessel, C.; Six, J. Simulating switchgrass biomass production across ecoregions using the Daycent model. GCB Bioenergy 2012, 4, 521–533. [Google Scholar] [CrossRef]
- Prather, A. The Impact of Integrated Crop-Livestock Systems: A Review of the Components and Barriers of the Classic Farming Approach. Master’s Thesis, Department of Diagnostic Medicine/Pathobiology, College of Veterinary Medicine, Kansas State University, Olathe, KS, USA, 2022. [Google Scholar]
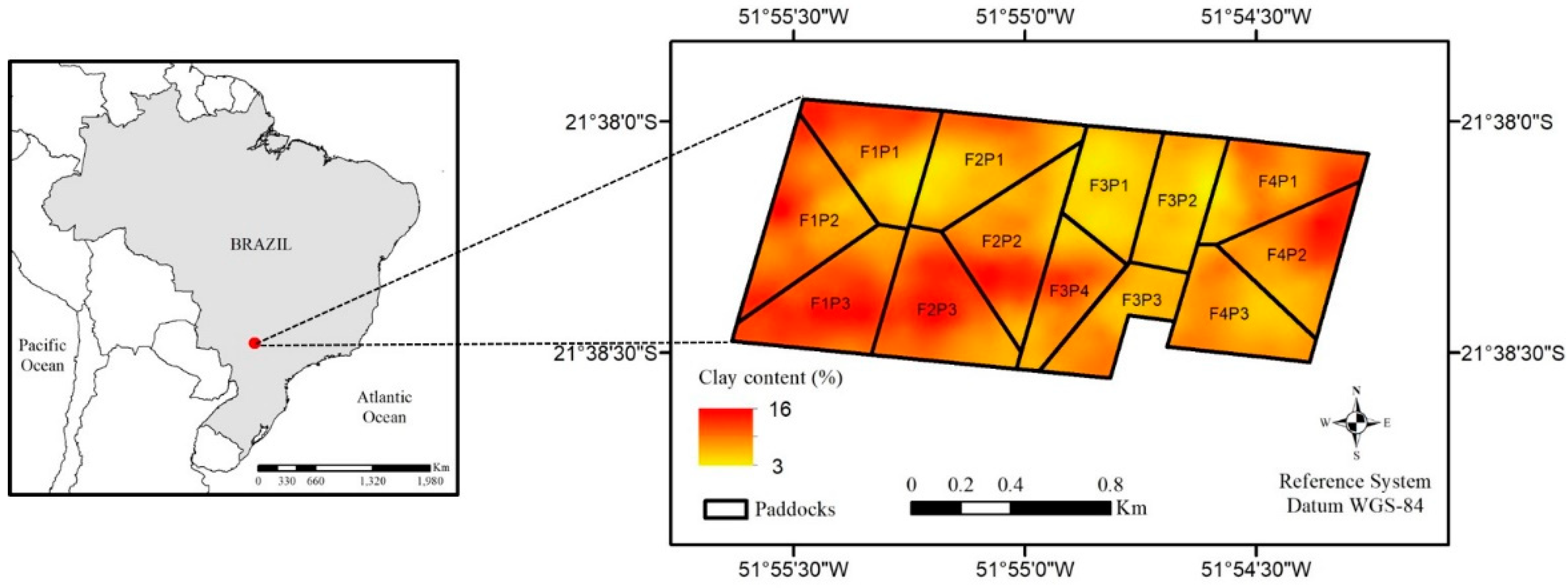





| Field | Paddock | Clay 1 (%) | Sand 1 (%) | Total C 1 (g kg−1) | pH 2 | TSWA 3 (mm mm−1) |
|---|---|---|---|---|---|---|
| 1 | 1 | 8.3 | 81.1 | 6.2 | 5.1 | 0.16 |
| 2 | 10.1 | 79.2 | 5.8 | 5.1 | 0.16 | |
| 3 | 10.3 | 78.0 | 5.4 | 5.1 | 0.16 | |
| 2 | 1 | 7.3 | 84.4 | 4.7 | 5.0 | 0.15 |
| 2 | 7.9 | 82.1 | 5.2 | 5.1 | 0.16 | |
| 3 | 9.9 | 79.9 | 4.8 | 5.1 | 0.16 | |
| 3 | 1 | 4.6 | 86.6 | 4.9 | 5.1 | 0.15 |
| 2 | 4.8 | 82.6 | 4.4 | 5.0 | 0.16 | |
| 3 | 6.2 | 82.8 | 4.0 | 5.0 | 0.16 | |
| 4 | 7.9 | 81.1 | 4.3 | 5.1 | 0.16 | |
| 4 | 1 | 9.7 | 81.4 | 6.0 | 4.9 | 0.16 |
| 2 | 9.1 | 79.1 | 4.7 | 4.9 | 0.16 | |
| 3 | 6.9 | 81.9 | 4.0 | 4.9 | 0.16 |
| Steps | Submodel | Modified Parameters | Value | ||
|---|---|---|---|---|---|
| Default | Calibrated | ||||
| Equilibrium | Tree | MT | PRDX(2) | 0.1–5.0 | 1.50 |
| Pre-Experiment | Fert | NPI3 | FERAMT(1) | 0–9999 | 0.80 |
| FERAMT(2) | 0–9999 | 1.83 a | |||
| NPI4 | FERAMT(1) | 0–9999 | 6.00 | ||
| FERAMT(2) | 0–9999 | 1.31 b | |||
| RRPNL | FERAMT(2) | 0–9999 | 0.0356 c | ||
| RRPL | FERAMT(2) | 0–9999 | 1.96 d | ||
| Crop | TKNBM | PRDX(1) | 0.1–5.0 | 1.60 | |
| TKNBM/TKNZ1 | PPDF(3) | 0–1.0 | 1.00 | ||
| PPDF(4) | 0–10.0 | 2.50 | |||
| BIOMAX | 0–1000 | 400 | |||
| PRAMN(1,1) | 1–100.0 | 30.00 | |||
| TKNZ1 | PRDX(1) | 0.1–5.0 | 1.20 | ||
| Graz | GH | FLGREM | 0–1.0 | 0.35 | |
| FDGREM | 0–1.0 | 0.15 | |||
| ICLS | Fert | MAP | FERAMT(1) | 0–9999 | 2.75 |
| FERAMT(2) | 0–9999 | 5.67 | |||
| Crop | MMBR | PRDX(1) | 0.1–5.0 | 1.60 | |
| PRAMN(1,1) | 1–100.0 | 30.00 | |||
| PRAMN(1,2) | 1–200.0 | 90.00 | |||
| PRAMX(1,1) | 1–200.0 | 35.00 | |||
| PRAMX(1,2) | 1–400.0 | 95.00 | |||
| SNFXMX | 0–1.0 | 0.05 | |||
| MMBR/SYBRS/SYAS/SYNS | BIOMAX | 0–1000 | 400 | ||
| MMBR | CLAYPG | 1–9.0 | 6.00 | ||
| SYBRS | PRDX(1) | 0.1–5.0 | 1.15 | ||
| SYAS/SYNS/SYBRS | FULCAN | 50–200 | 200 | ||
| EFRGRN(1) | 0–1.0 | 0.70 | |||
| PRAMN(1,1) | 1–100.0 | 5.00 | |||
| PRAMN(1,2) | 1–200.0 | 15.00 | |||
| PRAMX(1,1) | 1–200.0 | 15.00 | |||
| PRAMX(1,2) | 1–400.0 | 30.00 | |||
| SNFXMX | 0–1.0 | 0.065 | |||
| SYAS | PRDX(1) | 0.1–5.0 | 1.10 | ||
| SYNS | PRDX(1) | 0.1–5.0 | 1.20 | ||
| Harv | MOWTD | AGLREM | 0–1.0 | 0.05 | |
| BGLREM | 0–1.0 | 0.50 | |||
| MOWC | AGLREM | 0–1.0 | 0.82 | ||
| BGLREM | 0–1.0 | 0.80 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, Y.F.; Valadares, R.V.; Dias, H.B.; Cuadra, S.V.; Campbell, E.E.; Lamparelli, R.A.C.; Moro, E.; Battisti, R.; Alves, M.R.; Magalhães, P.S.G.; et al. Intense Pasture Management in Brazil in an Integrated Crop-Livestock System Simulated by the DayCent Model. Sustainability 2022, 14, 3517. https://doi.org/10.3390/su14063517
Silva YF, Valadares RV, Dias HB, Cuadra SV, Campbell EE, Lamparelli RAC, Moro E, Battisti R, Alves MR, Magalhães PSG, et al. Intense Pasture Management in Brazil in an Integrated Crop-Livestock System Simulated by the DayCent Model. Sustainability. 2022; 14(6):3517. https://doi.org/10.3390/su14063517
Chicago/Turabian StyleSilva, Yane Freitas, Rafael Vasconcelos Valadares, Henrique Boriolo Dias, Santiago Vianna Cuadra, Eleanor E. Campbell, Rubens A. C. Lamparelli, Edemar Moro, Rafael Battisti, Marcelo R. Alves, Paulo S. G. Magalhães, and et al. 2022. "Intense Pasture Management in Brazil in an Integrated Crop-Livestock System Simulated by the DayCent Model" Sustainability 14, no. 6: 3517. https://doi.org/10.3390/su14063517