Abstract
The forest community structure and dynamic structure provide basic data for in-depth study of the community maintenance mechanisms and succession processes of urban forest ecosystems. Moreover, the study on the dynamics of suburban forest communities can provide a reference for the succession process of forest communities under severe human disturbance. In this context, this study analyzes the species composition and community structure of a 6 ha subtropical secondary evergreen broad-leaved forest plot in Wuchaoshan National Forest Park, Hangzhou, Eastern China. There are 36,757 existing woody plants (with DBH ≥ 1 cm), belonging to 94 species (45 evergreen species, and 48 deciduous species) in 63 genera of 34 families, with a plant density of 6126 plants/ha. The overall size–class distribution shows an obvious inverted “J” shape. Species with the top-five important values in the sample are Schima superba, Camellia fraternal, Symplocos anomala, Cyclobalanopsis glauca, Eurya rubiginosa, accounting for 57.3% of all individuals within the sample. In addition, different species have different habitat preferences and different distribution areas. The Wuchaoshan forest is in the middle and late stages of the succession. Different interference histories in the community succession process are demonstrated through studies of secondary evergreen broad-leaved forests. Long-term community monitoring contributes to indicating the direction and potential of the secondary forest succession.
1. Introduction
The construction and development of urban forests not only enhances the urban image and comprehensive competitiveness but also functions as a significant measure to improve the carrying capacity of biodiversity and promote the sustainable development of the regional economy [1,2,3,4]. Researchers at home and abroad have conducted various research on the ecological functions, growth and development rules, biodiversity differences, and structural changes (tree species composition, plaque composition, landscape composition, and seasonal changes) of different types of plants in urban forests communities [2,3,4,5,6,7]. The study of suburban forests can provide valuable suggestions and theoretical support for urban forest construction, urban ecological restoration, and improvement of the human settlement environment. Jin et al. [8] studied the species composition and functional diversity of urban forest communities. The closer the distance to the forest edge, the greater the impact on the functional traits of the urban forest, and the less the impact on species diversity. Therefore, the study highlights the importance of preserving forest integrity by protecting inner habitats at the edges of new forests and increasing biodiversity conservation buffers [8]. The study on the construction law and succession of the suburban forest ecosystem community process can optimize the distribution and pattern of tree species in urban forests, improve the protection and restoration of suburban forests, enhance the construction of ecological corridors, etc. [8,9,10]. Currently, there is still a lack of systematic research on suburban forests. Therefore, the establishment of a large sample of urban forests helps study species coexistence or community-building, as well as provides a platform for interdisciplinary and comprehensive scientific biodiversity research on urban forests.
Species composition and community structure, to a certain extent, determine the structure and function of forest ecosystems, such as wind protection and sand fixation, water conservation, biodiversity conservation, and nutrient circulation [11]. Different forest types and succession stages have diverse forest ecosystem functions. For example, broad-leaf forests have high soil conservation value and water conservation function [11]. The greater the difference in the utilization of resources between different species in species-rich mixed forest communities, the stronger the carbon sink capacity, thus effectively maintaining the ecosystem stability [9,10]. In addition, the forest community structure affects the evapotranspiration and energy exchange between the forest and atmosphere in the urban green space ecosystem, which plays an important role in the regulation of the urban thermal environment [12,13]. The dynamic changes in community structure and population determine the function of forest division, reflecting the ability of plants to use resources and the stability of the community [14]. The radial structure of the population displays the age of the forest community, demonstrates the community renewal dynamics and the population adaptation to the habitat, and can predict the population development direction and succession trend [12,13,14]. The spatial distribution patterns of species partly reflect the integrated responses of factors, such as habitat preferences and life history strategies of different populations [15,16]. Each species has specific optimal habitat conditions, occupies different niche spaces, and exerts diverse ecological functions [17,18]. Therefore, research based on forest dynamic monitoring samples can help to monitor the long-term dynamic changes in species composition and community structure and provide a theoretical basis for studying the spatial and temporal patterns, succession processes, and forest ecosystem maintenance mechanisms [19]. The forest-monitoring plot can contain more tree species composition and community structure, which provides a platform for studying the community-building mechanism and ecosystem function [19].
The mid-subtropical evergreen broad-leaved forest is one of the most typical forest vegetation types mainly distributed in China, with complex species composition, diverse community structure, rich biodiversity, and significant ecosystem service functions [11], which plays a key role in maintaining ecosystem security and stability [20]. However, the subtropical evergreen broad-leaved aged forest in China has been damaged by long-term artificial disturbance. The secondary broad-leaved forest is the main forest type in Zhejiang Province, but its community structure is incomplete, and the ecological function needs to be improved [21]. There are many influencing factors in the recovery process of secondary forest communities. Community diversity and structural dynamics are jointly affected by environmental factors, such as the interference history and topography. Moreover, the recovery speed and direction show significant differences in different succession stages [18,22]. This study was conducted in the Wuchaoshan National Forest Park in Hangzhou Province between October 2020 and June 2021 according to the CTFS (center for tropical forest science) forest dynamic monitoring sample construction standard [16,23], and a 6 ha central subtropical evergreen forest dynamic monitoring prototype was established for the first time in the suburbs of Hangzhou. By analyzing the species composition, species diversity, and community radial structure of the sample site, a community survey and dynamic monitoring and assessment were also carried out. In addition, the forest dynamics and the community-construction process in the forest succession process under artificial interference were revealed, guiding biodiversity conservation and vegetation restoration in secondary evergreen broad-leaf forests.
2. Study Area and Methods
2.1. Overview of the Study Area
Wuchaoshan National Forest Park belongs to the veins of Tianmu Mountain, located in Xianlin Town, Yuhang District, Hangzhou City, Zhejiang Province (33°41′ N, 120°00′ E), China. It is about 20 km away from Hangzhou city, with a total area of 522 ha, an average altitude of 264 m, the highest altitude being 494.7 m. As a natural forest ecosystem in the suburbs of the city, Wuchaoshan National Forest Park is located in the central subtropical zone, with superior water resources and heat conditions. The annual average temperature is 16.1 °C, the average temperature of the coldest month in January is 3.6 °C, and the average temperature of the hottest month in August is 38.4 °C. Moreover, the annual sunshine hours reach 1970.6 h, and the plant growth period is 311 days; red soil and yellow soil are the main soil types. The forest coverage rate of Zhejiang Wuchaoshan National Forest Park is as high as 93% with rich plant species. Evergreen broad-leaved forest is the regional vegetation in the area, and trees that appear more frequently were Cyclobalanopsis glauca, Castanopsis chinensis, Schima superba, Castanopsis sclerophylla, etc. Before the 1970s, the whole reserve experienced human interference such as felling. In the past 40 years, the forest has been completely closed for afforestation.
2.2. Sample Setting and Community Survey
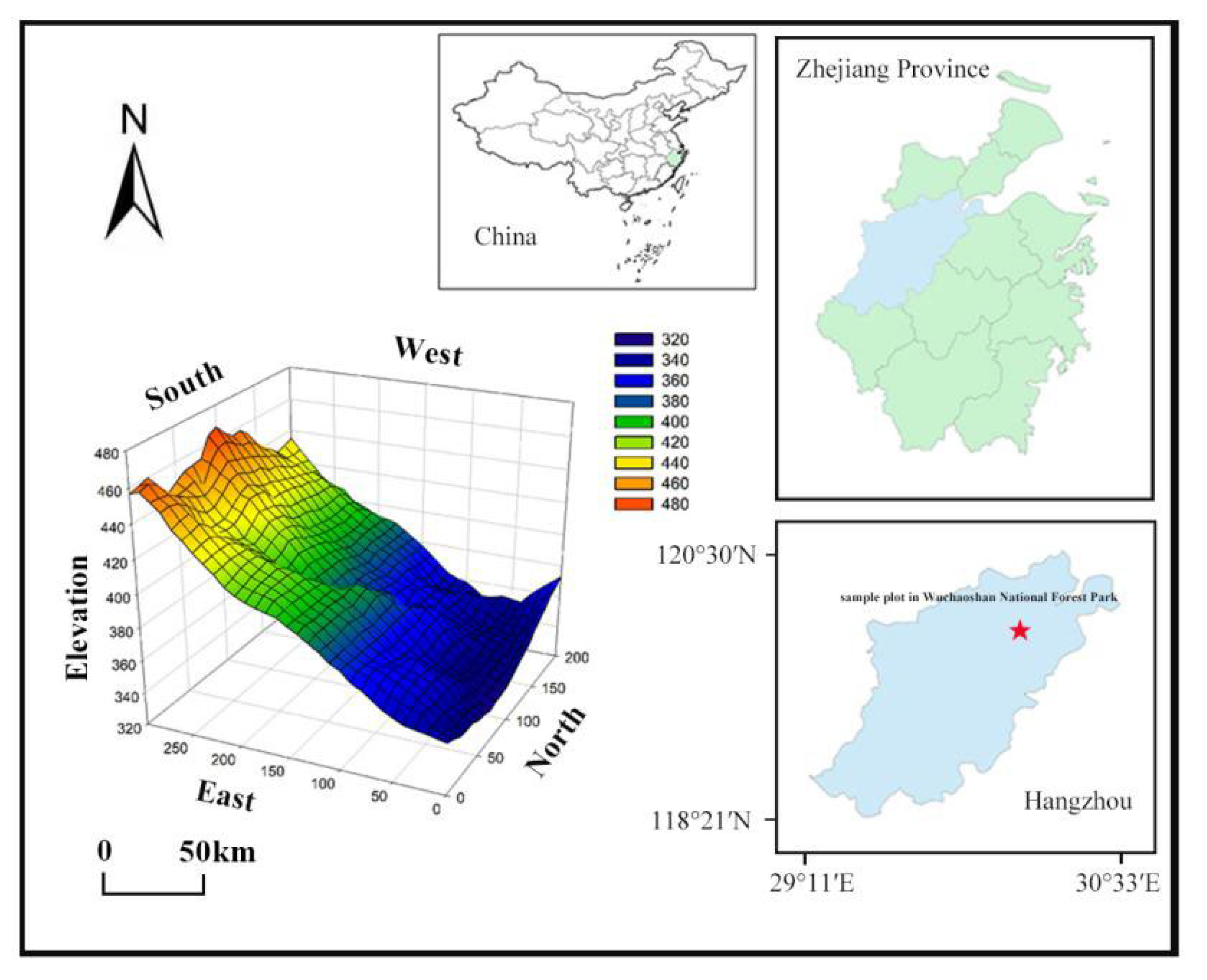
From October 2020 to June 2021, a well-recovered subtropical evergreen hardwood forest community was selected, and a 6 ha (200 m × 300 m) forest dynamic monitoring site was established according to the construction standard of CTFS (center for tropical forest science) dynamic monitoring sample. The 6 ha site was divided into 150 sample plots (each 20 m × 20 m), permanently marked with stainless steel tubes at 4 corners of each sample square (Figure 1). The origin coordinates of the sample site are 30°11′11.37″ N, 120°0′0.60″ E, and the elevation range is from 340.1 to 467.4 m. Each 20 m × 20 m sample square was divided into 16 sample squares (each 5 m × 5 m) to conduct a comprehensive survey of the sample plants. The individuals with surviving woody plants with a diameter at breast height of no less than 1 cm (with DBH ≥ 1 cm) in each sample were numbered using aluminum plates and marked with red paint at 1.3 m above the tree height. The species names, DBH, tree height, coordinates, branching, and initiation status were recorded for all labeled woody plant individuals [16,23].
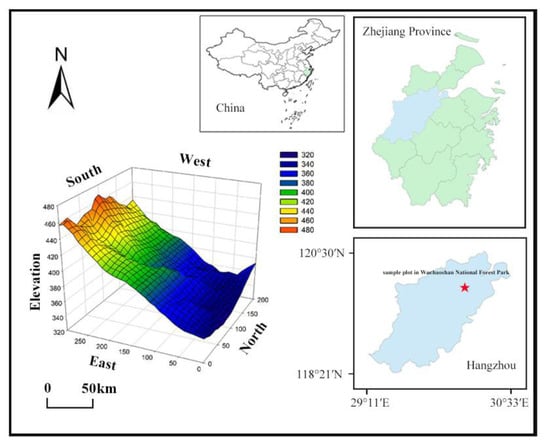
Figure 1.
Location and topographic map of forest dynamic monitoring plot in Wuchaoshan National Forest Park.
2.3. Data Analysis
The dominance of the species was measured by the importance value (IV). IV = (relative density + relative frequency + relative dominance)/3.
The species–area curve was plotted based on the sampled area of the sample and the corresponding number of species. Species abundance and species abundance rank distribution curves were plotted according to the correspondence between individual abundance and species. Data were processed using the “vegan” package of R software [24].
3. Result
3.1. Number of Individuals and Species Composition
A total of 36,757 surviving individual woody plants (DBH ≥ 1 cm) were recorded in the 6 ha dynamic monitoring sample of Wuchaoshan National Forest Park. The number of independent individuals was 25,390 with 5213 branches and 6154 sprouts. The individual density of the sample was 6126 units/ha (including branches and sprouts) or 4230 units/ha (excluding branches and sprouts). A total of 94 species of woody plants were recorded in 34 families and 63 genera. Among them, four species of gymnosperms were recorded in four families, which were Cupressaceae, Taxaceae, Taxodiaceae, Pinaceae respectively.
There were 45 species of evergreen woody plants (a total of 30,755 plants), accounting for 47.4% and 83.7% of the total number of species and individuals, respectively. There were 48 species of deciduous woody plants (a total of 6002 plants), representing 51.6% and 16.3% of the total number of species and individuals, respectively. The most common families in the sample were Fagaceae (10 species in 6 genera), Lauraceae (9 species in 5 genera), Rosaceae (8 species in 6 genera), Ericaceae (6 species in 3 genera), Aquifoliaceae (6 species in 1 genus), Theaceae (5 species in 4 genera), Symplocaceae (5 species in 1 genus). The number of species in these seven families accounted for 52.1% of the total number of samples, and that of plants (30,880 in total) reached 84.0%. The total basal area for the entire 6 ha sample was 184.43 m2, and the average basal area was 30.74 m2/ha. The species in the largest basal area of the sample was Schima superba, exceeding 82.90 m2.
3.2. The Species–Area, Species–Abundance, and Species–Abundance Rank Distribution Curves
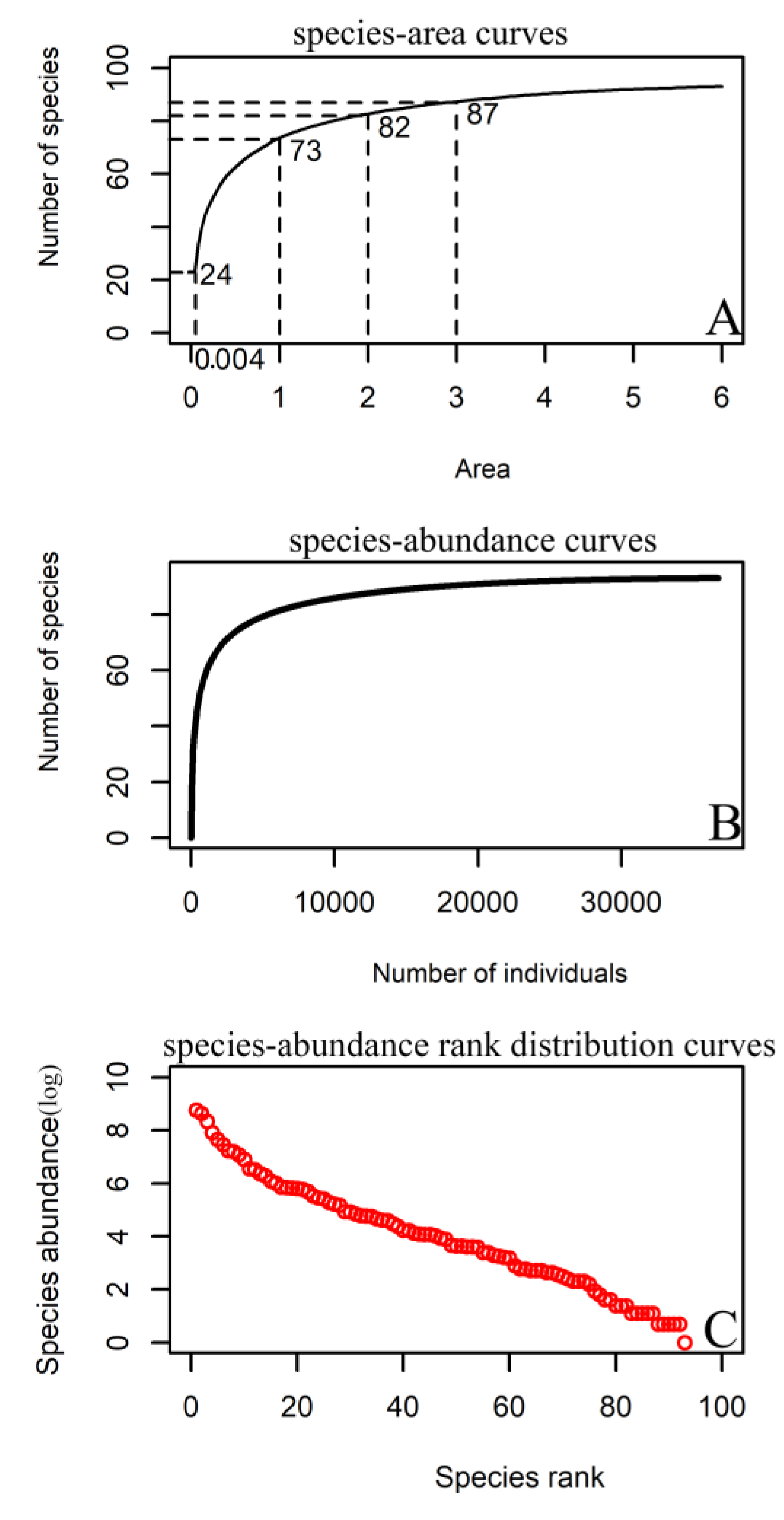
The species–area curve of the 6 ha sample (Figure 2A) showed that when the sampled area was 1 ha, the number of species was 73, accounting for 77.4% of the total number of sampled species. When the sampled area was 2 ha, the species number was 82, representing 87.2% of the total species number. When the sampled area reached 3 ha, the number of species was 87, representing 92.6% of the total species number. The species–abundance curve (Figure 2B) demonstrated that the number of species increased rapidly with the number of sampled individuals, and the rate gradually slowed as the number of sampled individuals increased. When the number of individuals sampled reached 10,000, the number of species was 86, representing 91.5% of the total species number. When the number of individuals sampled reached 20,000, the number of species was 91, representing 96.8% of the total species number. The species–abundance rank distribution curves (Figure 2C) displayed that the samples had obvious multiple dominant species and more rare species. There were 39 species with no more than 30 individuals in the sample and 19 species with fewer than 10 individuals in the sample.
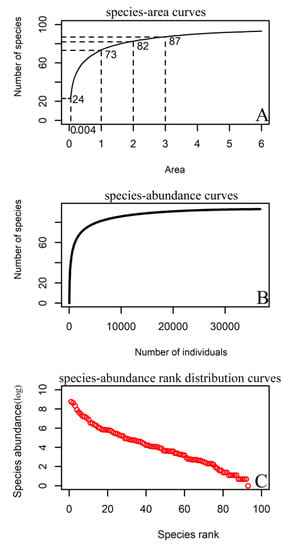
Figure 2.
The species–area (A), species–abundance (B), and species–abundance rank distribution (C) curves for woody plants in the 6 ha urban forest dynamics sample of Wuchaoshan, Zhejiang Province.
3.3. Community Radial Structure
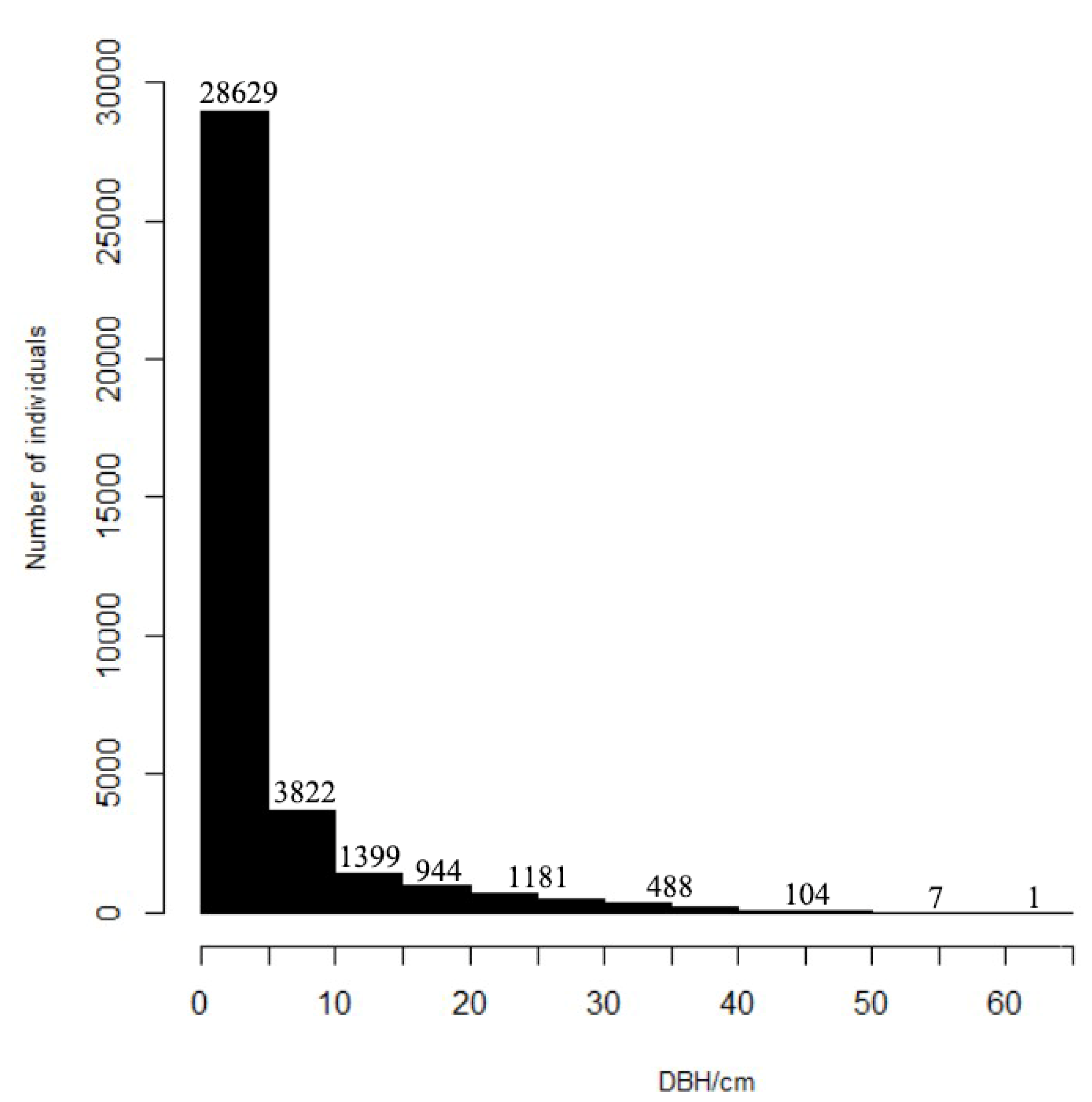
The average diameter at breast height of all woody plants in the sample was 4.76 cm, and the overall size–class distribution showed an obvious inverted “J” shape (Figure 3). The small-diameter plant individuals (1.0 cm ≤ DBH ≤ 5 cm) represent the highest proportion of the total number of individuals (Table 1), with 28811 plants, accounting for 78.38% of the total plant number. There were 32,633 plants with DBH < 10 cm, representing 88.78% of the total plant number. There were relatively few large-diameter individuals. To be specific, there were 1781 plants with DBH > 20 cm, accounting for 4.85% of the total plant number, and there were only 600 plants with DBH > 30 cm, accounting for 1.63% of the total plant number.
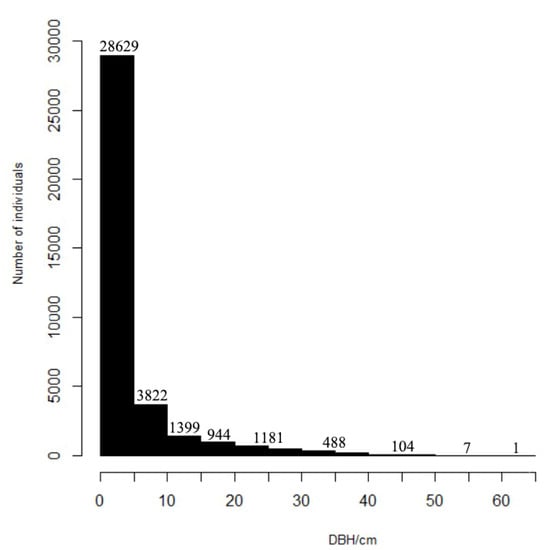
Figure 3.
Size–class distribution of diameter at breast height (DBH) ≥ 1 cm of woody plants in the 6 ha urban forest dynamics sample of Wuchaoshan, Zhejiang Province.

Table 1.
The number of stems, species, genera, and families with different minimum diameters at breast height (DBH) in the 6 ha urban forest dynamics sample of Wuchaoshan, Zhejiang Province.
3.4. Importance Values and Spatial Distribution of the Dominant Tree Species
There were 21 species with importance values greater than 1 (Table 2), representing 88.4% and 89.0% of the total number of individuals and the total basal area, respectively. The top-five species with important values and multiple individuals in the sample were completely consistent, in turn, which were Schima superba, Camellia fraternal, Symplocos anomala, Cyclobalanopsis glauca, Eurya rubiginosa, accounting for 57.3% of all individual abundance in the sample. The top-five species with the largest basal area were Schima superba, Cyclobalanopsis glauca, Cunninghamia lanceolata, Castanopsis sclerophylla, Quercus serrata, which accounted for 69.6% of the total basal area in the sample.
Table 2.
Dominant species of woody plants in the 6 ha urban forest dynamics sample of Wuchaoshan, Zhejiang Province.
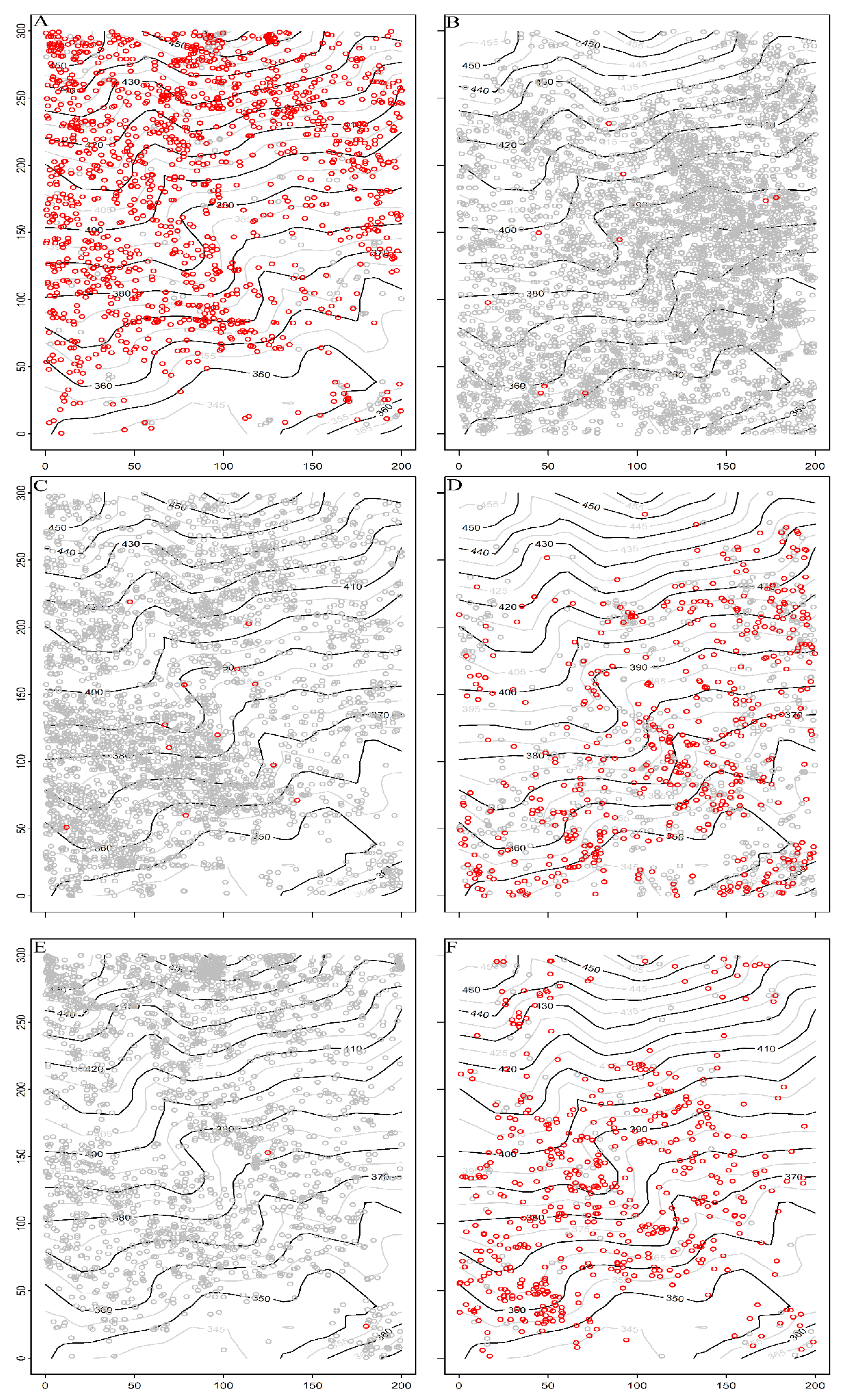
The distribution patterns of dominant species of Schima superba, Camellia fraternal, Symplocos anomala, Cyclobalanopsis glauca, Eurya rubiginosa, Cunninghamia lanceolata were significantly different, and the spatial distribution of large-diameter individuals (DBH > 10 cm) were more obvious (Figure 4). Schima superba tended to be distributed in clusters on slopes at higher altitudes, with large-diameter individuals of Schima superba mainly concentrated on the uphill position (Figure 4A). Camellia fraternal, Symplocos anomala and Eurya rubiginosa were small shrubs. Camellia fraternal mostly accumulated at low altitudes and rarely grew at higher altitudes (Figure 4B). Symplocos anomala always occupied the relatively gentle position of the low sea and was less distributed in the mountain valley (Figure 4C). The growth of Eurya rubiginosa on high-altitude slopes may be related to competition for resources such as light (Figure 4E). The distribution patterns of Cyclobalanopsis glauca and Cunninghamia lanceolata were exactly opposite to that of Schima superba. These two species were usually distributed in a small number of Schima superba, and large-diameter individuals were scattered at relatively low altitudes and relatively gentle terrain (Figure 4D,F).
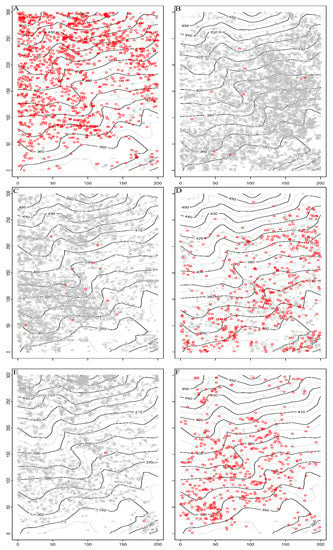
Figure 4.
Spatial distribution map of six dominant species in the 6 ha urban forest dynamics sample of Wuchaoshan, Zhejiang Province. Note: gray circles represent stems DBH < 10 cm, red circles represent stems ≥10 cm DBH. (A) Schima superba; (B) Camellia fraterna; (C) Symplocos anomala; (D) Cyclobalanopsis glauca; (E) Eurya rubiginosa; (F) Cunninghamia lanceolata. Note: Black and gray solid lines in the figure represent contour lines. The x-axis and y-axis indicate the length and width of the plot.
4. Discussion
The 6 ha sample of Wuchaoshan National Forest Park in Zhejiang Province is the first large forest dynamic monitoring sample to monitor the energy flow and material circulation of the urban forest ecosystem in China, hoping to supplement the long-term urban forest monitoring data on the China Forest Biodiversity Monitoring Network (CForBio) lacks. Studies on the basic ecological processes, biodiversity maintenance mechanisms, and forest dynamics of peri-urban forests can help clarify the different environmental gradients and response mechanisms of species adaptation and disturbance systems under climate change, enrich forest biodiversity, as well as protect and restore urban forest natural ecosystems [23]. This study helps to provide a scientific basis for sustainable urban development and ecological protection of physical and mental health in urbanized areas [3,4].
4.1. Species Composition and Structure of Urban Forest Communities
Species grouping and structural dynamics of forest communities demonstrate ecosystem processes and maintenance mechanisms [25], which reflects the direction and speed of the forest succession process [26]. Compared with the secondary evergreen broad-leaved forest in the 6 ha typical forest dynamic monitoring sample in Wuchaoshan National Forest Park, its species composition is similar to that of Gutian Mountain in Zhejiang [15,27,28], reflecting that the community species composition of low altitude zonal vegetation in Zhejiang province is similar [15,16]. The community had a small average diameter (the average diameter at breast height was 4.76 cm), and the large-diameter individuals were small. The average diameter at breast height is similar to the forest dynamic monitoring sample in other subtropical areas in China, such as Badagong Mountain samples in Hunan (5.41 cm) [29], Gutian Mountain (5.21 cm) [28], and Hubei Qizimei Mountain (4.38 cm). The whole sample size–class distribution presents an obvious inverted “J” type distribution. With the increase in diameter order, the number of individuals gradually decreases. After reaching a certain diameter order, the number of strains tends to stabilize [11], demonstrating the good regeneration status of the whole stand. As a typical aging forest structure, the main dominant population is naturally well updated and belongs to the growth type [11], indicating that the community can achieve self-renewal and facilitate stable development of the community [1,4]. These community features suggest that this secondary evergreen broad-leaved forest sample is in the middle and late stage of natural recovery after disturbance, which is usually one of the most diverse forest species [30]. At this stage, there are a large number of succession pioneer species (such as Sassafras tzumu, Alniphyllum fortunei), and relatively stable top community species (e.g., Castanopsis sclerophylla, Cyclobalanopsis glauca, etc.). It is believed that Schima superba and Castanopsis sclerophylla are the main dominant species in the subtropical evergreen broad-leaved forest, and Castanopsis sclerophylla occupies a larger proportion and more important value in the aged forest, while Schima superba is the main tree species in secondary forests. The evergreen species such as Schima superba, Cyclobalanopsis glauca, and Castanopsis sclerophylla have significant advantages in the important value and multiple degree proportion in the Wuchaoshan mountain sample, and the size–class distribution is more reasonable [29], having relatively stable stand regeneration and large trees. This study confirms the dominance of evergreen species at subtropical low altitudes, playing a more important role in biodiversity conservation and ecosystem services.
4.2. Spatial Distribution of the Species
The community function and community structure change through spatial distribution (colonization and death, etc.) [16,18]. It is believed that topographic factors significantly influence the distribution of species, and the local habitat differentiation caused by topography provides good growth space for tree species with different ecological strategies, which may be one of the mechanisms for regional species coexistence [15,18]. This study finds that the populations of Schima superba mainly clustered in the middle and upper slopes at higher elevations. The reason might be that the uphill position is far away from the urban roads and is less disturbed by the early artificial logging and damage, and thus, it has a high canopy density. Moreover, due to the shade tolerance of Schima superba at the seedling stage, the environment on the upper slopes favors seedling renewal. In addition, the uphill slope in the Wuchao mountain has high-quality soil fertilizer conditions, loose soil, and good water storage, which is conducive to the growth of Schima superba. At low elevations, anthropogenic activity is more frequent and relatively severe due to greater proximity to villages and roads. For example, in the early days, people would cut down trees as building materials or pay for charcoal, which destroyed the structure of the forest community. The forest windows formed after logging had a good light environment to promote the renewal and growth of Camellia fraternal, Alniphyllum fortunei, and Sassafras tzumu, as well as a rapid diffusion ability [16]. While these succession pioneer species serve as top tree species, seasonal leaves provide moderate shade and light to the lower evergreen shrubs, offering more niche space for species renewal and growth [17,18]. Therefore, small evergreen shrubs (Camellia fraterna; Symplocos anomala; Eurya rubiginosa. etc.) are more concentrated and distributed in the low-altitude flat-slope area with more light-demanding tree species. This study also finds significant differences in the spatial distribution of Cyclobalanopsis glauca and Cunninghamia lanceolata, suggesting that the two evergreen species may be competitive due to the same leaf habits [16,18], which are more resistant to cold temperatures and low light. In the case of low interference intensity, each tree species has equal competition opportunities for above-ground spatial resources; mainly lying in the competition of underground spatial resources, especially for soil nutrients, phosphorus and potassium may be important limiting elements in the region; these are important factors affecting the community composition and recovery direction in the region [29]. Therefore, in terms of forest vegetation restoration and precise forest quality improvement in the future, the forest recovery speed can be improved by increasing phosphorus fertilizer and potassium fertilizer [17].
4.3. Succession Process of Urban Forests
The secondary succession process of forest communities generally develops from deciduous to deciduous–evergreen broad-leaved mixed forest according to the sparse forest irrigation, and different interference intensities and types can change the succession speed and direction of the community. There are a large number of individuals of the species of Symplocaceae (as Symplocos anomala and Symplocos stellaris) in the Wuchao mountain sample, demonstrating the corresponding adaptability to different levels of disturbance, colonization, and rapid growth of tree species, and tall and straight above the crown, become the community interference. Some studies suggest that the medium artificial disturbance to community construction and forest community ecological restoration belongs to gain interference [31,32]. During forest succession, species diversity is mainly influenced by competition exclusion, interference processes, and environmental heterogeneity [18,22,28,30]. Moreover, the community structure tends to recover at significantly higher rates than the species richness [31]. With the progress of succession, the evergreen tree species of Cyclobalanopsis glauca, Castanopsis sclerophylla, and Schima superba gradually become the dominant tree species, and the urban forest of Zhejiang Wuchao Mountain shows the typical species composition and community structure of secondary evergreen broad-leaved forest in the middle and late stage of succession. Light-demanding evergreen tree species such as Schima superbas grow at a fast rate and play a significant role as the long-lived pioneer tree species in the recovery of evergreen broad-leaf forests. With the development of the community, the allocation of resources among populations becomes more and more uneven, with increasing species diversity of subtropical evergreen broad-leaf forest communities, more obvious dominant species effect, complex community structure, and improved stability [9]. Long-term dynamic monitoring of forest ecosystems that naturally return to communities after disturbance will help to study the changing law of resource competition in the succession process and provide a resource-sharing platform for exploring the succession process of forest communities, the maintenance mechanism, and the improvement of the ecosystem service function.
5. Conclusions
The suburban urban forest in Hangzhou belongs to the typical secondary evergreen broad-leaved forest with rich species composition and complex community structure, affected by human disturbance and other factors. Schima superba and Cyclobalanopsis glauca are the dominant species in the community, but there are relatively more deciduous species. This article aims at introducing the composition and structure of forest community during the restoration process of urban–suburban forests disturbed by human activities and expects long-term monitoring to explain the changes in forest community during the succession process. Long-term monitoring helps to understand the succession process and rules of urban forests under human disturbance and provides a theoretical basis for the conservation and maintenance of urban forest diversity. The researchers of this article will continue to monitor this large forest plot over a long period to explore community-building and maintenance mechanisms.
Author Contributions
Conceptualization, C.W. and L.Y.; methodology, Y.D.; software, B.J.; validation, T.L., J.J. and W.W.; formal analysis, Z.W.; investigation, X.Z.; resources, L.Y.; data curation, S.Y.; writing—original draft preparation, L.Y.; writing—review and editing, Y.D.; visualization, W.Y.; supervision, J.Z.; project administration, B.J.; funding acquisition, L.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by Zhejiang Provincial Scientific research Institute special project (2022F1068-2); the Major Collaborative Project between Zhejiang Province and the Chinese Academy of Forestry (2019SY08) and the Major Collaborative Project between Zhejiang Province and the Chinese Academy of Forestry (2021SY08).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
We would like to thank Zhejiang Hangzhou Urban Forest Ecosystem Research Station. And the support of all staff of Zhejiang Hangzhou Urban Forest Ecosystem Research Station.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aryal, P.C.; Aryal, C.; Bhusal, K.; Chapagain, D.; Dhamala, M.K.; Maharjan, S.R.; Chhetri, P.K. Forest Structure and Anthropogenic Disturbances Regulate Plant Invasion in Urban Forests. Urban Ecosyst. 2021, 7, 1–11. [Google Scholar] [CrossRef]
- Ballantyne, M.; Pickering, C.M. Differences in the Impacts of Formal and Informal Recreational Trails on Urban Forest Loss and Tree Structure. J. Environ. Manag. 2015, 159, 94–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenberg, J.W.N.; Millward, A.A.; Duinker, P.N.; Nowak, D.J.; Robinson, P.J. Neighbourhood-Scale Urban Forest Ecosystem Classification. J. Environ. Manag. 2015, 163, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Tavárez, H.; Elbakidze, L. Urban Forests Valuation and Environmental Disposition: The Case of Puerto Rico. For. Policy Econ. 2021, 131, 102572. [Google Scholar] [CrossRef]
- Alvey, A.A. Promoting and Preserving Biodiversity in the Urban Forest. Urban For. Urban Green. 2006, 5, 195–201. [Google Scholar] [CrossRef]
- Conway, T.M.; vander Vecht, J. Growing a Diverse Urban Forest: Species Selection Decisions by Practitioners Planting and Supplying Trees. Landsc. Urban Plan. 2015, 138, 1–10. [Google Scholar] [CrossRef]
- Wyse, S.v.; Beggs, J.R.; Burns, B.R.; Stanley, M.C. Protecting Trees at an Individual Level Provides Insufficient Safeguard for Urban Forests. Landsc. Urban Plan. 2015, 141, 112–122. [Google Scholar] [CrossRef]
- Jin, C.; Jiang, B.; Ding, Y.; Yang, S.; Xu, Y.; Jiao, J.; Huang, J.; Yuan, W.; Wu, C. Functional Traits Change but Species Diversity Is Not Influenced by Edge Effects in an Urban Forest of Eastern China. Urban For. Urban Green. 2021, 64, 127245. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Härdtle, W.; von Oheimb, G.; Yang, X.; et al. Impacts of Species Richness on Productivity in a Large-Scale Subtropical Forest Experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Pirnat, J. Conservation and Management of Forest Patches and Corridors in Suburban Landscapes. Landsc. Urban Plan. 2000, 52, 135–143. [Google Scholar] [CrossRef]
- Cortés-Calderón, S.; Mora, F.; Arreola-Villa, F.; Balvanera, P. Ecosystem Services Supply and Interactions along Secondary Tropical Dry Forests Succession. For. Ecol. Manag. 2021, 482, 118858. [Google Scholar] [CrossRef]
- Liu, H.; Weng, Q. Seasonal Variations in the Relationship between Landscape Pattern and Land Surface Temperature in Indianapolis, USA. Environ. Monit. Assess. 2008, 144, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Cao, F.; Wang, G. Effects of Spatial Pattern of Forest Vegetation on Urban Cooling in a Compact Megacity. Forests 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Zuppinger-Dingley, D. Plant Community History Affects the Interspecific Combining Ability of Plants. In Proceedings of the 97th ESA Annual Convention 2012, Portland, OR, USA, 5–10 August 2012. [Google Scholar]
- Jin, Y.; Russo, S.E.; Yu, M. Effects of Light and Topography on Regeneration and Coexistence of Evergreen and Deciduous Tree Species in a Chinese Subtropical Forest. J. Ecol. 2018, 106, 1634–1645. [Google Scholar] [CrossRef]
- Yao, L.; Xu, Y.; Jiang, B.; Wu, C.; Yuan, W.; Zhu, J.; Li, T.; Wang, Z. Competition Restricts the Growth, Development, and Propagation of Carpinus Tientaiensis: A Rare and Endangered Species in China. Forests 2021, 12, 503. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Ding, Y.; Yao, L.; Ai, X.; Zang, R. Trait Gradient Analysis for Evergreen and Deciduous Species in a Subtropical Forest. Forests 2020, 11, 364. [Google Scholar] [CrossRef] [Green Version]
- Mazei, Y.A.; Bubnova, O.A. Species Composition and Structure of Testate Amoebae Community in a Sphagnum Bog at the Initial Stage of Its Formation. Biol. Bull. 2007, 34, 619–628. [Google Scholar] [CrossRef]
- Edwards, D.P.; Tobias, J.A.; Sheil, D.; Meijaard, E.; Laurance, W.F. Maintaining Ecosystem Function and Services in Logged Tropical Forests. Trends Ecol. Evol. 2014, 29, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Shang, K.; Zhang, Q.; Da, L.; Hara, K.; Yang, Y.; Fujihara, M.; Tomita, M.; Zhao, Y. Effects of Natural and Artificial Disturbance on Landscape and Forest Structure in Tiantong National Forest Park, East China. Landsc. Ecol. Eng. Eng. 2014, 10, 163–172. [Google Scholar] [CrossRef]
- Bruelheide, H.; Böhnke, M.; Both, S.; Fang, T.; Assmann, T.; Baruffol, M.; Bauhus, J.; Buscot, F.; Chen, X.; Ding, B.; et al. Community Assembly during Secondary Forest Succession in a Chinese Subtropical Forest. Ecol. Monogr. 2011, 81, 25–41. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph Wright, S.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-ForestGEO: A Worldwide Network Monitoring Forests in an Era of Global Change. Glob. Change Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. 2016. Available online: https://www.r-project.org (accessed on 21 February 2022).
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher Levels of Multiple Ecosystem Services Are Found in Forests with More Tree Species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Aiba, S.; Hill, D.A.; Agetsuma, N. Comparison between Old-Growth Stands and Secondary Stands Regenerating after Clear-Felling in Warm-Temperate Forests of Yakushima, Southern Japan. For. Ecol. Manag. 2001, 140, 163–175. [Google Scholar] [CrossRef]
- Xiulian, C.; Qinggang, W.; Qiang, G.; Xian, Y.; Zhiyao, T. Sprouting Characteristics of Communities during Succession in an Evergreen Broad-Leaved Forest on Gutian Mountain, East China. Biodivers. Sci. 2019, 27, 24–32. [Google Scholar] [CrossRef]
- Henle, K.; Lindenmayer D, B.; Margules C, R.; Saunders D, A. Wissel C Species Survival in Fragmented Landscapes: Where Are We Now? Biodivers. Conserv. 2004, 13, 1–8. [Google Scholar] [CrossRef]
- Liu, Q.; Sterck, F.J.; Medina-Vega, J.A.; Sha, L.; Cao, M.; Bongers, F.; Zhang, J.; Poorter, L. Soil Nutrients, Canopy Gaps and Topography Affect Liana Distribution in a Tropical Seasonal Rain Forest in Southwestern China. J. Veg. Sci. 2021, 32, e12951. [Google Scholar] [CrossRef]
- Jian, M.; Liu, Q.; Tang, P.; Liang, Y. Floristic Analysis of the Evergreen Broad-Leaved Forest Community from Jiulianshan National Nature Reserve in Jiangxi Province, China. Front. For. China 2009, 4, 416–423. [Google Scholar] [CrossRef]
- Rozendaal, D.M.A.; Bongers, F.; Aide, T.M.; Alvarez-Dávila, E.; Ascarrunz, N.; Balvanera, P.; Becknell, J.M.; Bentos, T.v.; Brancalion, P.H.S.; Cabral, G.A.L.; et al. Biodiversity Recovery of Neotropical Secondary Forests. Sci. Adv. 2019, 5, eaau3114. [Google Scholar] [CrossRef] [Green Version]
- Simonyan, O.A.; Konovalova, M.A. Experience in Realizing the Educational Potential of Specially Protected Natural Areas of Malovishersky District, Novgorod Region. IOP Conf. Ser. Earth Environ. Sci. 2021, 852, 012099. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).