Abstract
Frequent M. aeruginosa outbreaks pose a major risk to public health and have a detrimental effect on aquatic ecosystems. Researchers are looking into ways to stop and control M. aeruginosa blooms, a problem that affects both the aquatic environment and human health significantly. It is important to develop proper monitoring methods to identify M. aeruginosa blooms. However, the existing control and monitoring techniques have some drawbacks that limit the field’s applicability. Therefore, we must improve current methods for effectively monitoring and controlling M. aeruginosa blooms. Mitigation strategies should be customized for particular bodies of water utilizing techniques that are fast, economical, and field-applicable. This review critically identifies and evaluates green technologies, especially those focused on the presence of M. aeruginosa in freshwater, and compares and discusses problems with these green technologies. Furthermore, they were characterized and ranked according to their cost, effectiveness, and field applicability. A few suggestions for improvements were provided, along with ideas for future research projects that would take anticipated environmental changes into account.
1. Introduction
Microcystis aeruginosa has been identified as one of the major bloom-forming cyanobacteria expanding globally as a result of intensifying eutrophication [1]. In May 2007, excessive nutrient concentrations and extremely warm temperatures led to a major hazardous Microcystis bloom in Lake Taihu, China, raising concerns across the globe [2]. Winnipeg lake has been declared “the threatened lake of the year” by the Global Nature Fund due to increased phosphorus concentrations [3]. The frequency and severity of severe summer–fall cyanobacterial harmful algal blooms (cHABs) have increased, according to satellite imagery and measurements of the lake’s biomass [4]. Microcystin concentrations can reach high levels during cyanobacterial blooms in Lake Erie’s western basin [5]. The city of Toledo, Ohio, received a “do not drink” warning in August 2014 after microcystin levels in the water exceeded the recommended limit of 1 g/L set by the World Health Organization [6]. In several regions of the Laurentian Great Lakes, the poisonous cyanobacterium Microcystis aeruginosa has developed into a regular summertime occurrence, raising public concerns [7]. Numerous reports of cyanobacterial blooms in the Guadiana River along its downstream course through Portuguese territory have been produced. These blooms are typically dominated by the potentially toxic Microcystis spp. [8]. Problematic cyanobacterial blooms have also affected the Murray River in New South Wales (NSW). Their concentrations exceeded 4 mm3 L−1. The National Health and Medical Research Council (2008) established this as the red alert threshold for recreational water consumption in Australia, at which point New South Wales begins to take bloom management measures [9]. From the 1980s to 2022, Microcystis aeruginosa dramatically increased in frequency and abundance throughout the world, causing high levels of water contamination and affecting human health [10,11].
Reducing pollution from both point and nonpoint sources is vital to controlling eutrophication via regulation of nitrogen (N) and phosphorus (P) inputs [12]. Nitrogen (N) and phosphorus (P) are the principal nutrients of concern because their supply frequently affects aquatic life [13]. Effective treatments and preventive measures, along with their detailed mechanisms (Figure 1), must be developed to control M. aeruginosa blooms, which have constituted a significant threat to the security of the aquatic environment [14].
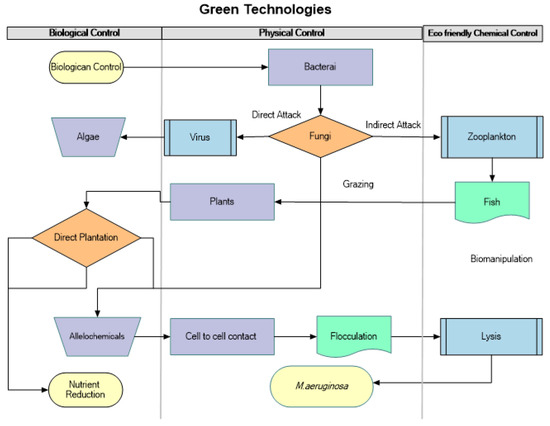
Figure 1.
Green technologies and associated mechanism to control M. aeruginosa.
To reduce M. aeruginosa abundance and eliminate toxins from freshwater, the purpose of this narrative review is to focus on current green physical, chemical, and biological technologies. However, it is true that the deployment of most chemical and physical techniques is constrained due to negative ecological impacts, expensive costs, or low field operability. Since few techniques for addressing the harmful M. aeruginosa blooms can be applied on a wide scale in the field, it is necessary to consider a variety of strategies, including microorganisms, aquatic animals, plant allelopathy, and clay applications. We systematically compared and analyzed the development status, advantages and disadvantages, applicable conditions, and future development trends of green technologies for monitoring and controlling M. aeruginosa blooms.
2. Green Technologies to Control Microcystis aeruginosa
The three main options available to date for treating and controlling the growth of M. aeruginosa are:
- Physical methods
- Chemical methods
- Biological methods.
2.1. Phyical Control
Freshwater treatment frequently involves physical methods and water quality standards are constantly being improved in many countries [15]. It is considered as an emergency preventive measure rather than a control method [16]. Physical preventive measures include: harvesting of M. aeruginosa [17], air flotation [18], magnetic flocculation [19], hydrodynamic cavitation [20], light shading [21], dredging sediments [22], ultrasound technology [23], flocculants [24], etc.
Toxic algal blooms can be removed by air extraction by using tiny bubbles to attach to the algae. Through the action of the minute oxygen bubbles, fish are protected from hypoxia and harmful algal blooms. There may be restrictions on the use of large mechanical bubblers in areas where toxic algal blooms are a problem [25].
Ultrasonic treatment has been suggested as a major control measure that can destroy algal cells on a local scale [26]. It is useful to control M. aeruginosa blooms due to the chemical and physical effects of sonication [27].
Disadvantages of Physical Control
Large blooms represent a challenge for the majority of physical strategies because they are generally expensive and slow [28]. Physical approaches are mainly utilized as emergency measures for algal blooms rather than as a control strategy, as they are not always practical [29]. The cost and impact of physical measures for reducing dangerous algal blooms are poorly understood due to a lack of field experience [30]. Ultrasound technology is highly unlikely to have any control impact on harmful algal blooms in natural systems, except for extremely high-intensity ultrasound used within an extremely small body of water. Such intense ultrasound has been shown to destroy zooplankton grazers and may have an impact on fish behavior and population [31,32,33].
2.2. Chemical Control
Recently, there has been much discussion concerning the use of clay additions to suppress hazardous algal species, a strategy that has been utilized successfully in several aquaculture operations worldwide [34]. Clays, the primary component of soil, have many advantages over other materials, including the fact that they are cheap, easy to use in the field, and non-polluting [35]. As a result, it is considered the most promising mitigation strategy against harmful algal blooms [36].
2.2.1. Natural and Modified Clays
A very viable and ecologically sustainable method is to use natural, nontoxic, and inexpensive clays to remove harmful algal blooms [37,38]. The most efficient flocculants have been observed to be clays such as montmorillonite, kaolinite, and phosphatic, with the lowest loading of 0.25 g/L and a removal efficiency of 90% [24,39,40,41,42]. The author of [42] stated that 5720 publications on harmful algal blooms (HABs) were examined throughout the course of the previous 30 years. While research publications regarding the use of natural clays to inhibit M. aeruginosa growth have been very limited until now. In two papers [16], natural clays were used to manage M. aeruginosa in Lake Taihu, China. There are presently few studies on M. aeruginosa’s reaction to naturally occurring, typical clay particles like kaolinite and montmorillonite.
Disadvantages of Natural Clays
The clay technique is still hampered by the following issues: Natural clays have low flocculating efficiency, which is the most serious drawback and often leads to the requirement of an exorbitant amount of clays to achieve an effective efficiency in the field [43]. For example, it was noted that the dosage of clay in aquaculture sites in Japan ranged from 110 to 400 t/km2 [44], and 384 t/km2 of loess was utilized in Korea to reduce Cochlodinium polykrikoides blooming [45]. Algae can be eliminated naturally and non-toxically by clay precipitation and flocculation; however, this process can also bring on new water blooms [46]. The negative charge of natural clays prevents them from being an ideal flocculant. Clays have not yet been given a standard definition. Since antiquity, these materials have been used and studied from a variety of angles, giving rise to various conflicting terminology [16]. Therefore, when using the clay method in the field, the barrier is caused by low removal efficiency, a high dose, and substantial deposition loads on the sediments [47,48]. Unclear mechanisms and a lack of systematic kinetic studies of clay–cell flocculation in fresh and seawaters are other reported issues of natural clays [24].
2.2.2. Modified Clays
Another control method is the application of modified clays to treat M. aeruginosa blooms. Currently, common flocculants (such as sediments [49], modified clay [50], iron salts [51], aluminum salts [50], etc.) are frequently employed to control M. aeruginosa blooms in freshwaters. Organic polymer chitosan is also combined with soil or other ballast component to treat M. aeruginosa blooms [52].
In several works, a number of modified clays have been assayed with M. aeruginosa. Two phosphate fixatives i.e., Phoslock®, Europe GmbH (Zug, Switzerland)and Aqual-PTM, (Tokoroa, New Zealand) have been revealed to have marginal impact on M. aeruginosa and reduce its growth rates by reducing phosphate [50]. They were used in conjunction with eight other compounds to combat M. aeruginosa. Montmorillonite modified lime-ceramic sand-lake sediments [49], hexadecyl trimethyl ammonium bromide (CTAB) modified clays [53], modified attapulgite [54], modified vermiculite [55], amphoteric starch-based bicomponent modified soil [56], cationic starch modified soils etc. are some recent modified clays used to inhibit M. aeruginosa. Table 1 shows the removal efficiency of modified clays and their applicability at lab and field scale.

Table 1.
Removal efficiency of modified clays and their applicability at lab and field scale.
Disadvantages of Modified Clays
It was noted that stirring operations might be one of the modified clay’s biggest drawbacks in real-world applications [61]. Moreover, they also affect benthic flora and fauna. According to a study by [52], the effect of sediment loading on phytoplankton communities was studied and it was observed that dinoflagellates switched to heterotrophy in numerous degrees, and some dinoflagellates shaped impermanent cysts. In terms of practical applicability, flocs settling on surfaces is another of the biggest drawbacks [55]. According to [43], the majority of research on the eco-environmental effects of clay or modified clay (MC) has focused mostly on describing phenomena and findings, whereas mechanistic analysis of modified clay (MC) effects is rather weak. As a result, mechanistic research that necessitates both more in-depth examinations of mechanisms and theoretical knowledge must be reinforced.
2.2.3. Eco-Friendly Chemicals
Some copper-related compounds, chlorine, and oxidizing agents like H2O2 have historically been widely used to control M. aeruginosa blooms. They are considered to be relatively safe materials [62]. A novel environmentally safe and selective algaecide called 2-((1,3,4-thiadiazol-2-yl)thio)-N-(4-chlorophenyl) acetamide (Q2) was created to suppress M. aeruginosa blooms [10]. It was different from the impact on ecosystem functioning of the traditionally used harmful algaecide diuron. Their findings demonstrated that Q2 might be beneficial to the aquatic environment and offered a novel approach to the management of harmful cyanobacterial blooms (HCBs) in the future. Additionally, chitosan’s anionic characteristics have been shown to make it an effective flocculant for removing cyano-HABs from water resources [63]. Ferric or aluminum salts are also extensively studied for controlling cyanobaterial blooms [64]. Some surfactants and engineered nanoparticles like titanium dioxide, silver nanoparticles, zinc oxide, and yttrium(III) oxide are also used to improve algae removal efficiency [65]. The results of recent studies on compounds that are eco friendly, their applicability at lab and field scales, and the factors examined are compiled in Table 2.
Table 2.
Recent studies conducted on the application of eco-friendly chemicals to manage M. aeruginosa blooms.
Disadvantages of Eco-Friendly Chemicals
Copper addition to lakes and reservoirs raises concerns about heavy metal accumulation and toxicity [85]. H2O2 is widely used in water treatment and in the aquaculture industry [91]. However, different types of water bodies react differently and different amounts of H2O2 are required to control cyanoHABs, ranging from 2 to 20 mg L−1 [83,84,85,86]. Varying background stressors in freshwater ecosystems may interact with H2O2, altering its efficacy in controlling cyanoHABs. Co-occurring stressors can have complex impacts on organisms and communities as stressors in combination can either amplify (synergistic) or attenuate (antagonistic) effects [92,93,94]. The variation of the dose of H2O2 required might therefore be linked to the growing number of background stressors faced by aquatic ecosystems. These background stressors include a temperature increase, an elevated level of CO2, anthropogenic inputs such as pharmaceuticals, personal care products, pesticides, and, relevant to this study, tiny plastic fragments [58]. Chitosan is a non-toxic and biodegradable material, but the acidic condition of a chitosan solution sprayed over a water body for the control of M. aeruginosa can negatively influence water quality. Tiny chitosan particles (i.e., chitosan nanoparticles) can also cause physiological stress in aquatic biota [63].
2.3. Biological Control
One alternative approach to the control of algal blooms involves the use of biological control (biocontrol) agents [95]. Biological control includes the use of microorganisms [96], plants [97] and biomanipulation approach [98] to control M. aeruginosa blooms. Compounds such as biochar [99] are also in use to control M. aeruginosa.
2.3.1. Microorganisms Control
Microorganisms such as viruses, bacteria, actinomycetes, fungi, amoebae, and cyanophages have been shown to kill cyanobacteria.
Bacteria
Among these, antagonistic bacteria have the potential to become useful agents for algal control, as they are simple to culture and manipulate [100]. Bacteria, being one of the most common and varied species in the aquatic environment, form complex ecological interactions with cyanobacteria, including predation, competition, mutualism, commensalism, and amensalism [101]. Those bacteria that display obviously adverse effects on cyanobacterial growth are recognized as cyanobactericidal bacteria [102].
Against pathogenic M. aeruginosa, recently discovered algicidal bacteria A. bestiarum HYD0802-MK36 and P. syringae KACC10292T have been found to be effective [103]. The growth of M. aeruginosa can be hampered by a number of bacteria from the genera Aeromonas [104] and Pseudomonas [105]. These bacteria affect the growth of M. aeruginosa by two modes (Figure 2): direct attack and indirect attack [106].
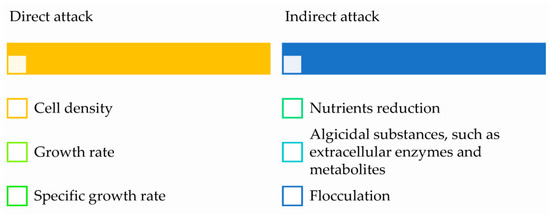
Figure 2.
Attacking modes of cyanobactericidal bacteria against M. aeruginosa.
Direct attack mode requires physical contact between predatory bacteria and cyanobacteria [107]. The Bdellovibrio-like bacteria lysed M. aeruginosa by breaking down cell structures after penetrating the host cell [108]. Indirect attack mode occurs when bacteria suppress or kill cyanobacteria without physically contacting them. Indirect assault methods include:
- Releasing cyanobactericidal substances, such as extracellular enzymes [105]
- Release of metabolites (e.g., indole, 3-oxo-α-ionone) [109]
- Deteriorating algal survival environment, e.g., by competing nutrients [110]
- Flocculating algae cells [111,112]
Cyanobactericidal bacteria affecting M. aeruginosa are listed in Table 3, according to the published literature.
Table 3.
Recent studies conducted on application of bacteria to manage M. aeruginosa blooms, Removal efficiency: RE, References: Ref.
- (1)
- Disadvantages of Bacteria
Several cyanobactericidal bacteria or their released chemicals have proven useful in reducing cyanobacterial blooms in the environment. There are many uncertainties when it comes to using cyanobactericidal bacteria or compounds to effectively control or eliminate cyanobacterial blooms in natural waters. Cyanobacteria in natural water tend to have stronger resistance than laboratory culture due to the colonial form of algal cells [155,156] and that the single- or two-celled Microcystis used in cyanobactericidal research is less resilient than the colon [157]. The financial expenses of using cyanobactericidal bacteria or allelopathic chemicals for bloom-control on a wide scale should be a concern. Simply put, the size of their contribution must be sufficient to overcome the relevant cutoff. If the channels for agent generation (i.e., bacterial culture) are not cost-effective, the application will be constrained. Before using cyanobactericidal bacterial agents, it is important to weigh the potential consequences for the environment [152].
The use of the cyanobactericidal bacterium Lysobacter enzymogenes subsp. enzymogenes AL-1 to eradicate M. aeruginosa in a microcosm was determined to be of high ecological concern [158]. A considerable decrease in ciliates, flagellates, and fungi was seen as a result of the use of cyanobactericidal bacteria. For example, using L-lysine to reduce Microcystis blooms led to blooms of Euglena sp. and Phormidium sp. in ponds, [159], suggesting that the removal of certain cyanobacterial blooms with these agents can generate other unforeseen algal bloom problems. Therefore, in order to achieve an algicidal effect on target cyanobacteria, the use of cyanobactericidal bacteria and substances in the control of cyanobacterial blooms in natural waters must overcome biological and abiotic uncertainties. The widespread use of cyanobactericidal microorganisms or chemicals in aquatic ecosystems requires first conducting biosafety studies [135,136].
Fungi
Degradation by fungal strains and the elimination of cyanobacterial cells are mainly unexplored [160]. There are just 15 known fungus species that may inhibit and lyse cyanobacterial cells. There are several structural similarities between the two fungus groups to which these species belong: Ascomycetes (nine of the species) and Basidiomycetes (six of the species). In addition, it has been claimed that fungi play a significant role in water treatment, with strong data showing that microcystins (MC) breakdown by fungal stains is faster [155] than that occurring by bacterial strains [156,157]. Some fungi (such as Aurobasidium pullulans and Trichoderma citrinoviride) have been shown to restrict the development of cyanobacteria while leaving the growth of more beneficial algae alone [155,161]. As an interesting side note, certain fungal strains (Trichaptum abietinum, Trichoderma citrinoviride) showed a dual-functional feature, efficiently lysing cyanobacteria and decomposing MCs produced by the decaying cells [162]. In Table 4, we have a brief overview of several recently discovered algicidal fungi that are effective against Microcystis aeruginosa.
Table 4.
Summary of some recent algicidal fungi active against Microcystis aeruginosa.
- (1)
- Disadvantages of Fungi
Although fungi have been infrequently reported as potential biological controllers, there are parasitic associations between freshwater microalgae and Chytridiomycetes fungi [169,170] and with biflagellate fungi belonging to Oomycetes. However, this latter association occurs to a lesser extent [171]. This fungal infection in planktonic diatoms has been associated with mortality of host organisms, suppression or retardation of phytoplankton blooms, and changes in the size, distribution, and composition of planktonic populations and communities [172,173,174,175]. Moreover, there is scarce information concerning the relationship between freshwater toxic microalgae and pathogenic fungi [16].
Virus
Viral treatment may be one of the important factors that can control HABs [176]. The virus typically uses species-specific interaction [177], the bursting of cells, and the viral lytic cycle. Viral degradation has the advantage of a species-specific attack [178]. Cyanophage (Ma-LMM01) specifically infects a toxic strain of the bloom-forming cyanobacterium Microcystis aeruginosa [179,180]. Cyanophage infection may have a significant impact on the succession of cyanobacteria in the pond. Cyanophage from Myoviridae family isolated from Chinese freshwater; GenBank, accession number KF356199.1., named MaMV-DC, was thought to have a half-life of between 24 and 48 h and 80 infectious units per cell (Microcystic aeruginosa FACHB-524). Cyanophages are found to be effective biocontrol agents of M. aeruginosa. [181,182,183].
A novel, wide-ranging freshwater cyanophage called MinS1has the ability to infect multiple different cyanobacterial orders and could be used as a biological control measure against cyanobacterial blooms [184]. In host–range experiments, a novel freshwater cyanophage called Mae-Yong1326-1 was effective in lysing M. aeruginosa FACHB-1326 [181].
- (1)
- Disadvantages of Virus
Many biological phenomena related to viruses are poorly understood because of host specificity and seasonal issues. For example, several reports suggested that algal viruses often existed in stable numbers, even when their hosts were absent [185]. Reports claimed that the summer and spring seasons are showing the highest decay rates of cultivated viruses after four seasons of analysis [186]. The seasonal study found that the low decay rates of the algal virus during the winter allowed for the survival of about 126 days under the ice cover in a frozen freshwater pond [182]. Another thing is that these agents show high specificity and efficiency, but they have the limitation of high cost and require upscaled level experiment confirmation [183].
Phytoplankton and Zooplankton
Recent efforts to control toxic algal blooms have concentrated on isolating anti-algal active compounds from micro- and macro-algae [187]. Marine macro-algae (seaweed) extraction has yielded many compounds with the potential to inhibit many other micro-algae, including red tide dinoflagellates [188]. Micro- and macroalgae are reported as effective biocontrol agents, but these studies are mainly focused on the removal of red tides [65]. Research studies regarding the use of algae against M. aeruginosa removal are limited.
Zooplankton are able to limit the dominance of cyanobacteria in aquatic ecosystems up to a certain density [189]. The majority of the data on the function of MC comes from research with generalist grazers (Daphnia), which only occasionally coexist with cyanobacterial blooms [190,191]. M. aeruginosa physiological and metabolic changes are affected by Daphnia magna exudates [192]. However, another investigation using copepods suggested that MC may serve as a warning to avoid Microcystis [193].
- (1)
- Disadvantages of Phytoplankton and Zooplankton
Mechanisms of cyanobacterial metabolites and their impacts in the presence of grazers are scarce [194]. While some studies demonstrate that the production of MC is induced by zooplankton [195], other studies contend that the zooplankton may even impede the formation of MC [196]. Although it has been proposed that zooplankton can affect the dynamics of cyanobacterial metabolites, their chemical structures are unknown since detection and isolation of these organisms are difficult [197]. Limitations on the predator’s potential usage outside the lab result from logistical challenges in applying the predator and growing the culture to produce enough zooplankton predators [65].
2.3.2. Fish
Due to some fish’s ability to consume and digest the poison directly, fish species have always employed this technique for bloom clearance. To regulate HABs for the lake ecology, bio-manipulation is a viable method [198]. There have been numerous attempts to reduce cyanobacterial blooms in China and other regions by utilizing filter-feeding fish such as bighead carp and silver carp, which have occasionally proved successful [57,199,200].
Field trials were conducted to eliminate Microcystis blooms by stocking tilapia in Lake Yuehu and other eutrophic lakes in Ningbo, China, between 2000 and 2003, as well as feeding studies to evaluate tilapia’s consumption and digestion of M. aeruginosa in a lab setting [201]. Microcystis could be consumed and digested in significant amounts by tilapia. At a water temperature of 25 °C, the digestion efficiency ranged from 58.6% to 78.1%. Salazar Torres [202] study provides evidence of reducing cyanobaterial biomass almost 60% in the presence of Oreochromis niloticus in eutrophic reservoirs. Hybrids of silver carp Hypophthalmichthys molitrix and bighead carp H. nobilis have been reported to alter phytoplankton species composition [203].
Disadvantages of Fish
Animal growth may be slowed down or impeded because of the potential for poor digestion of Microcystis species and potential low/imbalanced nutritional values. The digestive proteases trypsin and/or chymotrypsin can be inhibited by the protease inhibitors produced by Microcystis spp., such as aeruginosins, cyanopeptolins, micropeptins, microviridins, and microcins [204]. The reallocation of energy to the detoxification of MCs and other cyanotoxin also resulted in reduced animal growth. This strategy for managing algal blooms is not highly suggested due to the difficulties (Figure 3) linked with health hazards for animals from the digestion of Microcystis aeruginosa and the reported enormous mortality of fish species [32].
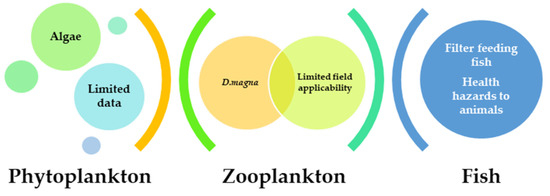
Figure 3.
Major issues limiting the application of phytoplankton, zooplankton, and fish species to control M. aeruginosa.
2.3.3. Plants
Allelopathic application is a promising strategy to control HABs. As a method inspired by natural phenomena, the effectiveness of allelochemicals in inhibiting microalgae cells has been discovered and confirmed for many years [205]. Both planting macrophytes [142] and adding extracted allelochemicals [97] were effective for introducing inhibition effects on microalgae cells. Four main categories of allelochemicals, including polyphenolics [206], N-containing compounds [207], fatty acids/eaters [208], terpenoids [209], and their derivatives, were proved to be efficient in M. aeruginosa-inhibiting capacities, respectively. Figure 4 shows an overview of allelochemicals and their effects on algae.
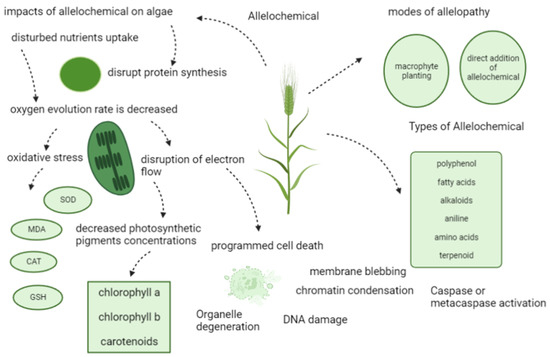
Figure 4.
An illustration of allelochemicals and their mode of action.
The sensitivities of microalgal species upon allelochemicals were significantly different and M. aeruginosa was widely confirmed as the most sensitive microalgal species to allelochemicals [210].
Allelochemicals induced damages on multiple levels of microalgal cells, including interfering the photosynthesis, generating oxidative stress, triggering programmed cell death (PCD), and disturbing other physiological and biochemical processes (Figure 5) [211].
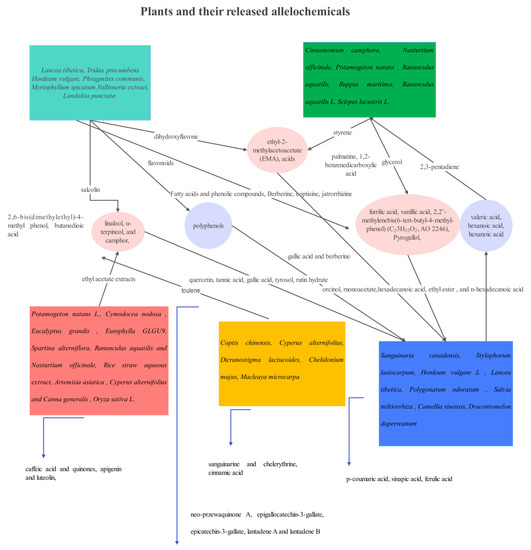
Figure 5.
Plants and their released allelochemicals effective against M. aeruginosa.
Disadvantages of Plants
Though many plant derived allelochemicals have been screened, few are feasible candidates for application in field environments. Few reasons discourage the application of plant-derived allelochemicals in field environments (Figure 6).
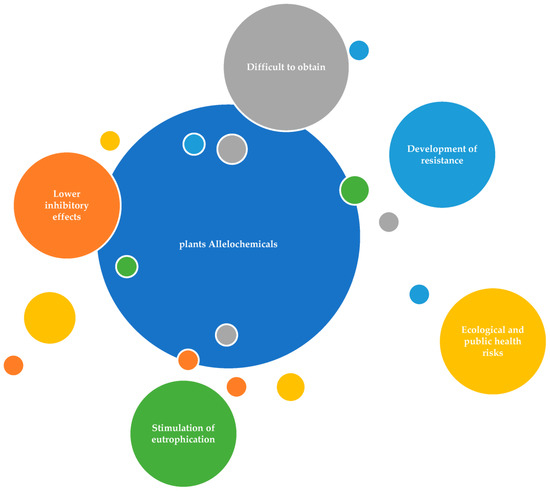
Figure 6.
Problems associated with the application of allelochemicals.
Some allelochemicals show only weak inhibitory effects on cyanobacteria. For example, bionone is an antialgal compound that inhibits Microcystis, but the EC50 is 22 mg/L [212]. Cyanobacteria can adapt to the inhibitory effect of some biologically derived substances (BDS) and become resistant to them. Nonanoic acid was reported as an allelochemical showing a strong inhibitory effect on M. aeruginosa, with a median effective concentration (EC50) as low as 0.5 mg/L [208]; however, a following study indicated that, under the stress of nonanoic acid, cells of M. aeruginosa soon adapt to this environment [213]. Some allelochemicals are difficult to obtain. Even though some natural antialgal chemicals strongly inhibit cyanobacteria, the supply of these biologically derived chemicals is limited, and the structures of those antialgal chemicals are very complex, so their chemical synthesis is difficult or prohibitively expensive. For example, Tellimagrandin II originating from M. spicatum shows a strong inhibitory effect on Anabaena [171], but the content of Tellimagrandin II in M. spicatum is very low, and the structure of this chemical is too complex for facile synthesis by chemical engineers. In this way, the extract of plants, such as barley straw, may be more applicable in algae control since it is cheaper and easier to obtain. Beside the potential damage of allelochemicals to nontarget aquatic organisms, the health risks of allelochemicals to humans are also not known. Allelochemicals such as lysine, rice hull, and wheat bran leachate include N and/or P, which may increase bioavailable N and/or P in waters where they are applied, thereby exacerbating eutrophication [214].
3. Summary of Limitations of Green Technologies
This review thoroughly explained current and previous green technologies, their efficiency and problems associated with the success rate of all of these applications which have been applied over the years to mitigate M. aeruginosa. We noticed the following crucial aspects (Figure 7).
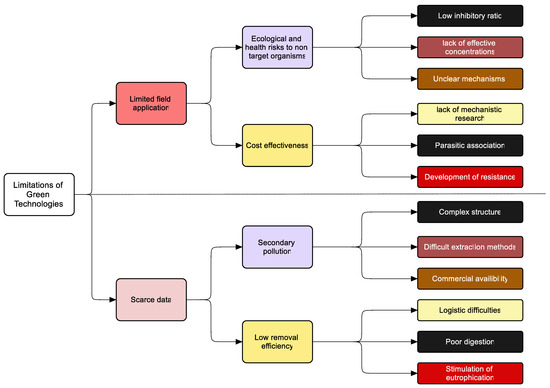
Figure 7.
Summary of limitations of green technologies.
- Physical methods are preferred to chemical methods, but they are expensive and are not easy to adapt in field conditions.
- Chemical methods are efficient in M. aeruginosa removal, but they are a source of secondary pollution.
- For the mitigation of M. aeruginosa, many biological control agents existed that includes bacteria, fungi, phages, zooplankton, plants, fish, etc. Many reports of laboratory success have been reported, but when it comes to field management, the success rate appears quite low.
4. Conclusions and Future Solutions
Regular M. aeruginosa outbreaks are harmful to aquatic ecosystems and constitute a serious risk to the public’s health. Research on M. aeruginosa removal is mainly focused on physical, chemical, or biological techniques. Each of these techniques has the potential to be effective in removing algae, there are certain drawbacks, and several techniques differ significantly from one another. It is important to do in-depth study on the integration of various approaches when assessing the social, economic, and environmental benefits as well as other comprehensive variables. Physical methods, i.e., harvesting of M. aeruginosa, air flotation, magnetic flocculation, hydrodynamic cavitation, light shading, dredging sediments, ultrasound technology, and flocculants, have promising applications. Physical method application has two main problems: they are expensive and difficult to implement at larger scale. Compared to physical control, chemical control—which includes natural, modified clays, and eco-friendly chemicals—is heavily debated.
However, there are conflicting results regarding the application of clay and the induction of physiological stress in aquatic biota by eco-friendly chemicals, which restricts the use of this approach. As an alternative to physical and chemical control, biological control focuses on the utilization of biological agents, such as bacteria, fungi, viruses, plants, etc., their released products (plants + allelochemicals), and biomanipulation (fish) techniques. Among biological agents, bacterial species have been frequently reported as potential biological control agents. Parasitic association of fungi, poorly understood mechanisms of viruses, zooplankton, phytoplankton, and poor digestion of M. aeruginosa still raise many questions about biological control techniques. In conclusion, we think that some effective, affordable, and environmentally friendly new algae removal methods and their combination processes are the future development direction.
Based on this narrative review, the following are some of the proposed solutions to get rid of M. aeruginosa blooms and combat currently existing problems with green technologies.
- There is a need to further explore the use of natural clays because of their abundance, cost effectiveness, and easy application. The only problem which has been mentioned in literature is their lower removal efficiency, which has been dealt by using modified clay but still data regarding the application of natural clays on M. aeruginosa is scarce.
- Combined application of ecofriendly chemicals and biological agents should be studied to evaluate their efficiency in M. aeruginosa blooms removal.
- Effect of physical, ecofriendly chemicals and biological agents on nutrient concentrations is also required to understand control mechanism deeply
- Further research is required regarding the effects of all these green technologies, i.e., physical, ecofriendly chemical, and biological methods on non-target organisms.
Keeping in mind all above proposed solutions, more research is needed to fully implement any of these methods in the field for the achievement of a sustainable environment.
Author Contributions
Z.K. and S.H.: Conceptualization, writing—original draft preparation, A.B., H.T.J., G.K., A.A., and E.B.: investigation and review and editing, M.W., R.G., T.Z., and L.L.: validation, data Curation. All authors have read and agreed to the published version of the manuscript.
Funding
The study was financially supported by National Natural Science Foundation of China (52170165, 11672139, 51861135314).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agathokleous, E.; Peñuelas, J.; Azevedo, R.A.; Rillig, M.C.; Sun, H.; Calabrese, E.J. Low Levels of Contaminants Stimulate Harmful Algal Organisms and Enrich Their Toxins. Environ. Sci. Technol. 2022, 56, 11991–12002. [Google Scholar] [CrossRef]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A Drinking Water Crisis in Lake Taihu, China: Linkage to Climatic Variability and Lake Management. Environ. Manag. 2009, 45, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Global Nature Fund. Threatened Lake of the Year 2013: Lake Winnipeg in Canada. 2013. Available online: https://www.globalnature.org/35753/Living-Lakes/Threatened-Lake-2016/Threatened-Lake-2013/resindex.aspx (accessed on 1 April 2023).
- Wassenaar, L.I.; Rao, Y.R. Lake Winnipeg: The forgotten great lake. J. Great Lakes Res. 2012, 38, 1–5. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Mishra, S.; Kane, D.D.; Bade, D.L.; Stanislawczyk, K.; Slodysko, K.N.; Jones, K.W.; Parker, E.M.; Fox, E.L. Cyanobacterial blooms in the central basin of Lake Erie: Potentials for cyanotoxins and environmental drivers. J. Great Lakes Res. 2019, 45, 277–289. [Google Scholar] [CrossRef]
- Francy, D.S.; Brady, A.M.; Stelzer, E.A.; Cicale, J.; Hackney, C.; Dalby, H.D.; Struffolino, P.; Dwyer, D.F. Predicting microcystin concentration action-level exceedances resulting from cyanobacterial blooms in selected lake sites in Ohio. Environ. Monit. Assess. 2020, 192, 513. [Google Scholar] [CrossRef]
- Wynne, T.T.; Stumpf, R.P.; Tomlinson, M.C.; Warner, R.A.; Tester, P.A.; Dyble, J.; Fahnenstiel, G.L. Relating spectral shape to cyanobacterial blooms in the Laurentian Great Lakes. Int. J. Remote Sens. 2008, 29, 3665–3672. [Google Scholar] [CrossRef]
- Moreno, I.; Cameán, A.; Tavares, M.J.; Pereira, P.; Franca, S. Toxicity of Cyanobacteria Isolated from the Guadiana River. Aquat. Ecosyst. Health Manag. 2003, 6, 409–413. [Google Scholar] [CrossRef]
- Bowling, L.C.; Merrick, C.; Swann, J.; Green, D.; Smith, G.; Neilan, B.A. Effects of hydrology and river management on the distribution, abundance and persistence of cyanobacterial blooms in the Murray River, Australia. Harmful Algae 2013, 30, 27–36. [Google Scholar] [CrossRef]
- Ke, M.; Feng, L.; Huang, S.; Lu, T.; Yu, Z.; Yang, Y.; Hu, H.; Peijnenburg, W.J.; Feng, L.; Qian, H. Development of a Potentially New Algaecide for Controlling Harmful Cyanobacteria Blooms Which is Ecologically Safe and Selective. J. Agric. Food Chem. 2022, 70, 10134–10143. [Google Scholar] [CrossRef]
- Preston, T.; Stewart, W.D.P.; Reynolds, C.S. Bloom-forming cyanobacterium Microcystis aeruginosa overwinters on sediment surface. Nature 1980, 288, 365–367. [Google Scholar] [CrossRef]
- Peng, J.; Jin, C.; Wu, Y.; Hou, Z.; Gao, S.; Chu, Z.; Zheng, B. Modeling Non-Point Source Nutrient Loads with Different Cropping Systems in an Agricultural Lake Watershed in Southwestern China: From Field to Watershed Scale. Mathematics 2022, 10, 4047. [Google Scholar] [CrossRef]
- Xia, Y.; Zhang, M.; Tsang, D.C.W.; Geng, N.; Lu, D.; Zhu, L.; Igalavithana, A.D.; Dissanayake, P.D.; Rinklebe, J.; Yang, X. Recent advances in control technologies for non point source pollution with nitrogen and phosphorous from agricultural runoff: Current practices and future prospects. Appl. Biol. Chem. 2020, 63, 8. [Google Scholar] [CrossRef]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived Intensification in Harmful Algal Blooms Is a Wave of Cumulative Threat to the Aquatic Ecosystems. Biology 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ding, S.; Li, L.; Liao, Q.; Chu, W.; Li, H. Ultrasound-enhanced coagulation for Microcystis aeruginosa removal and disinfection by-product control during subsequent chlorination. Water Res. 2021, 201, 117334. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Rodríguez, J.J.; Astuya-Villalón, A.; Llanos-Rivera, A.; Avello-Fontalba, V.; Ulloa-Jofré, V. A critical review on control methods for harmful algal blooms. Rev. Aquac. 2018, 11, 661–684. [Google Scholar] [CrossRef]
- Lin, Z.; Xu, Y.; Zhen, Z.; Fu, Y.; Liu, Y.; Li, W.; Luo, C.; Ding, A.; Zhang, D. Application and reactivation of magnetic nanoparticles in Microcystis aeruginosa harvesting. Bioresour. Technol. 2015, 190, 82–88. [Google Scholar] [CrossRef]
- Teixeira, M.R.; Rosa, M.J. Comparing dissolved air flotation and conventional sedimentation to remove cyanobacterial cells of Microcystis aeruginosa: Part I: The key operating conditions. Sep. Purif. Technol. 2006, 52, 84–94. [Google Scholar] [CrossRef]
- Noor, M.H.M.; Wong, S.; Ngadi, N.; Inuwa, I.M.; Opotu, L.A. Assessing the effectiveness of magnetic nanoparticles coagulation/flocculation in water treatment: A systematic literature review. Int. J. Environ. Sci. Technol. 2021, 19, 6935–6956. [Google Scholar] [CrossRef]
- Li, P.; Song, Y.; Yu, S. Removal of Microcystis aeruginosa using hydrodynamic cavitation: Performance and mechanisms. Water Res. 2014, 62, 241–248. [Google Scholar] [CrossRef]
- Chen, X.; He, S.; Huang, Y.; Kong, H.; Lin, Y.; Li, C.; Zeng, G. Laboratory investigation of reducing two algae from eutrophic water treated with light-shading plus aeration. Chemosphere 2009, 76, 1303–1307. [Google Scholar] [CrossRef]
- Lürling, M.; Faassen, E.J. Controlling toxic cyanobacteria: Effects of dredging and phosphorus-binding clay on cyanobacteria and microcystins. Water Res. 2012, 46, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Park, C.-B.; Baik, S.; Kim, S.; Choi, J.-W.; Lee, S.-H.; Kim, Y.J. The use of ultrasonic frequencies to control the bloom formation, regrowth, and eco-toxicity in Microcystis aeruginosa. Int. J. Environ. Sci. Technol. 2017, 14, 923–932. [Google Scholar] [CrossRef]
- Pan, G.; Zhang, M.-M.; Chen, H.; Zou, H.; Yan, H. Removal of cyanobacterial blooms in Taihu Lake using local soils. I. Equilibrium and kinetic screening on the flocculation of Microcystis aeruginosa using commercially available clays and minerals. Environ. Pollut. 2006, 141, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Zhan, M.-M.; Liu, P.-R.; Liu, X.-Y.; Hong, Y.; Xie, X. Inactivation and Removal Technologies for Algal-Bloom Control: Advances and Challenges. Curr. Pollut. Rep. 2021, 7, 392–406. [Google Scholar] [CrossRef]
- Wu, X.; Joyce, E.M.; Mason, T.J. Evaluation of the mechanisms of the effect of ultrasound on Microcystis aeruginosa at different ultrasonic frequencies. Water Res. 2012, 46, 2851–2858. [Google Scholar] [CrossRef]
- Lee, T.J.; Nakano, K.; Matsumara, M. Ultrasonic Irradiation for Blue-Green Algae Bloom Control. Environ. Technol. 2001, 22, 383–390. [Google Scholar] [CrossRef]
- Anderson, D.M. Approaches to monitoring, control and management of harmful algal blooms (HABs). Ocean. Coast. Manag. 2009, 52, 342–347. [Google Scholar] [CrossRef]
- Anderson, D.M. Prevention, control and mitigation of harmful algal blooms: Multiple approaches to HAB management. Harmful Algae Manag. Mitig. 2004, 123–130. Available online: https://www.researchgate.net/publication/255649174_Prevention_control_and_mitigation_of_harmful_algal_blooms_multiple_approaches_to_HAB_management (accessed on 1 April 2023).
- Park, J.; Church, J.; Son, Y.; Kim, K.-T.; Lee, W.H. Recent advances in ultrasonic treatment: Challenges and field applications for controlling harmful algal blooms (HABs). Ultrason. Sonochem. 2017, 38, 326–334. [Google Scholar] [CrossRef]
- Chambers, L.A.; Harvey, E.N. Some histological effects of ultrasonic waves on cells and tissues of the fish lebistes reticulatus and on the larva of rana sylvatica. J. Morphol. 1931, 52, 155–164. [Google Scholar] [CrossRef]
- Chen, G.; Ding, X.; Zhou, W. Study on ultrasonic treatment for degradation of Microcystins (MCs). Ultrason. Sonochem. 2020, 63, 104900. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, L.; Qi, Y.; Ma, C. Imaging mass spectrometry of interspecies metabolic exchange revealed the allelopathic interaction between Microcystis aeruginosa and its antagonist. Chemosphere 2020, 259, 127430. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Feng, Y.; Li, H.; Yang, Y.; Wu, R. Research progress of phosphorus adsorption by attapulgite and its prospect as a filler of constructed wetlands to enhance phosphorus removal from mariculture wastewater. J. Environ. Chem. Eng. 2022, 10, 108748. [Google Scholar] [CrossRef]
- Talawar, M.; Sivabalan, R.; Mukundan, T.; Muthurajan, H.; Sikder, A.; Gandhe, B.; Rao, A.S. Environmentally compatible next generation green energetic materials (GEMs). J. Hazard. Mater. 2009, 161, 589–607. [Google Scholar] [CrossRef]
- Ibrahim, N.H.; Iqbal, A.; Mohammad-Noor, N.; Razali, R.; Yanto, D.H.Y.; Wilson, L.D.; Mahadi, A.H. A Review on The Biological, Physical and Chemical Mitigation of Harmful Algal Bloom. Squalen Bull. Mar. Fish. Postharvest Biotechnol. 2022, 17, 95–110. [Google Scholar] [CrossRef]
- Anderson, D.M. Turning back the harmful red tide—Commentary. Nature 1997, 388, 513–514. [Google Scholar] [CrossRef]
- Sengco, M.R.; Li, A.; Tugend, K.; Kulis, D.; Anderson, D.M. Removal of red- and brown-tide cells using clay flocculation. I. Laboratory culture experiments with Gymnodinium breve and Aureococcus anophagefferens. Mar. Ecol. Prog. Ser. 2001, 210, 41–53. [Google Scholar] [CrossRef]
- Avnimelech, Y.; Troeger, B.W.; Reed, L.W. Mutual Flocculation of Algae and Clay: Evidence and Implications. Science 1982, 216, 63–65. [Google Scholar] [CrossRef]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994; Volume 10. [Google Scholar]
- Pan, G. A Method for Simultaneously Clearing up Harmful Algal Blooms and Harnessing Organic Pollutants to Promote the Primary Productivity in the Sea. UK Patent GB2337749, 11 March 1975. [Google Scholar]
- Sha, J.; Xiong, H.; Li, C.; Lu, Z.; Zhang, J.; Zhong, H.; Zhang, W.; Yan, B. Harmful algal blooms and their eco-environmental indication. Chemosphere 2021, 274, 129912. [Google Scholar] [CrossRef]
- Song, X.; Zhang, Y.; Yu, Z. An eco-environmental assessment of harmful algal bloom mitigation using modified clay. Harmful Algae 2021, 107, 102067. [Google Scholar] [CrossRef]
- Shirota, A. Red tide problem and countermeasures. II. Int. J. Aqua. Fish. Technol. 1989, 1, 195–223. [Google Scholar]
- Anderson, D.M.; Andersen, P.; Bricelj, V.M.; Cullen, J.J.; Rensel, J.J. Monitoring and Management Strategies for Harmful Algal Blooms in Coastal Waters; UNESCO: Paris, France, 2001. [Google Scholar]
- Zeng, G.; Zhang, R.; Liang, D.; Wang, F.; Han, Y.; Luo, Y.; Gao, P.; Wang, Q.; Wang, Q.; Yu, C.; et al. Comparison of the Advantages and Disadvantages of Algae Removal Technology and Its Development Status. Water 2023, 15, 1104. [Google Scholar] [CrossRef]
- Getchis, T.; Shumway, S. Harmful Algae: An Executive Summary. Connecticut Sea Grant College Program; CTSG-17-08; Background and Purpose: Groton, CT, USA, 2017; 16p. [Google Scholar]
- Yu, Z.; Sengco, M.R.; Anderson, D.M. Flocculation and removal of the brown tide organism, Aureococcus anophagefferens (Chrysophyceae), using clays. J. Appl. Phycol. 2004, 16, 101–110. [Google Scholar] [CrossRef]
- Xia, T.; Wang, S.-Q.; Yan, H.; Gao, Z.-Z.; Qiu, Y.; Yuan, F.; Huang, G.-Y.; Zhou, J. Effect of hydrodynamic condition on algae control based on montmorillonite modified lime-ceramic sand-lake sediments. Water Qual. Res. J. 2022, 57, 200–214. [Google Scholar] [CrossRef]
- Kang, L.; Mucci, M.; Lürling, M. Compounds to mitigate cyanobacterial blooms affect growth and toxicity of Microcystis aeruginosa. Harmful Algae 2022, 118, 102311. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Li, P.; Xu, B.; Wang, L.; Ma, G.; Chen, S.; Tan, F.; Shao, Y.; Zhang, L.; Yang, Z.; et al. A novel technology using iron in a coupled process of moderate preoxidation–hybrid coagulation to remove cyanobacteria in drinking water treatment plants. J. Clean. Prod. 2022, 342, 130947. [Google Scholar] [CrossRef]
- Noyma, N.P.; de Magalhães, L.; Furtado, L.L.; Mucci, M.; van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M.; Lürling, M. Controlling cyanobacterial blooms through effective flocculation and sedimentation with combined use of flocculants and phosphorus adsorbing natural soil and modified clay. Water Res. 2015, 97, 26–38. [Google Scholar] [CrossRef]
- Liu, G.; Fan, C.; Zhong, J.; Zhang, L.; Ding, S.; Yan, S.; Han, S. Using hexadecyl trimethyl ammonium bromide (CTAB) modified clays to clean the Microcystis aeruginosa blooms in Lake Taihu, China. Harmful Algae 2010, 9, 413–418. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, H.; Liu, X.; Cai, D.; Feng, H.; Miao, C.; Wang, X.; Wu, Z.; Yu, Z. Flocculation of harmful algal blooms by modified attapulgite and its safety evaluation. Water Res. 2011, 45, 2855–2862. [Google Scholar] [CrossRef]
- Miao, C.; Tang, Y.; Zhang, H.; Wu, Z.; Wang, X. Harmful algae blooms removal from fresh water with modified vermiculite. Environ. Technol. 2014, 35, 340–346. [Google Scholar] [CrossRef]
- Jin, Y.; Pei, H.; Hu, W.; Zhu, Y.; Xu, H.; Ma, C.; Sun, J.; Li, H. A promising application of chitosan quaternary ammonium salt to removal of Microcystis aeruginosa cells from drinking water. Sci. Total Environ. 2017, 583, 496–504. [Google Scholar] [CrossRef]
- Xie, P.; Liu, J. Practical Success of Biomanipulation using Filter-Feeding Fish to Control Cyanobacteria Blooms: A Synthesis of Decades of Research and Application in a Subtropical Hypereutrophic Lake. Sci. World J. 2001, 1, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; O’brien, A.M.; Lins, T.F.; Shahmohamadloo, R.S.; Almirall, X.O.; Rochman, C.M.; Sinton, D. Effects of Hydrogen Peroxide on Cyanobacterium Microcystis aeruginosa in the Presence of Nanoplastics. ACS ES&T Water 2021, 1, 1596–1607. [Google Scholar] [CrossRef]
- Gao, L.; Pan, X.; Zhang, D.; Mu, S.; Lee, D.-J.; Halik, U. Extracellular polymeric substances buffer against the biocidal effect of H2O2 on the bloom-forming cyanobacterium Microcystis aeruginosa. Water Res. 2015, 69, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Song, Q.; Long, J.; Song, G.; Mi, W.; Bi, Y. Optimization method for Microcystis bloom mitigation by hydrogen peroxide and its stimulative effects on growth of chlorophytes. Chemosphere 2019, 228, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, J.M. Phytoplankton and episodic suspended sediment loading: Phosphate partitioning and mechanisms for survival. Limnol. Oceanogr. 1992, 37, 974–988. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Hydrogen peroxide, an ecofriendly remediation method for controlling Microcystis aeruginosa toxic blooms. J. Appl. Phycol. 2020, 32, 3133–3142. [Google Scholar] [CrossRef]
- Park, Y.H.; Kim, S.; Kim, H.S.; Park, C.; Choi, Y.-E. Adsorption Strategy for Removal of Harmful Cyanobacterial Species Microcystis aeruginosa Using Chitosan Fiber. Sustainability 2020, 12, 4587. [Google Scholar] [CrossRef]
- Zhu, Y.; Cheng, S.; Wang, P.; Chen, H.; Zhang, X.; Liu, L.; Li, X.; Ding, Y. A possible environmental-friendly removal of Microcystis aeruginosa by using pyroligneous acid. Ecotoxicol. Environ. Saf. 2020, 205, 111159. [Google Scholar] [CrossRef] [PubMed]
- Balaji-Prasath, B.; Wang, Y.; Su, Y.P.; Hamilton, D.P.; Lin, H.; Zheng, L.; Zhang, Y. Methods to control harmful algal blooms: A review. Environ. Chem. Lett. 2022, 20, 3133–3152. [Google Scholar] [CrossRef]
- Xian, X.; Li, X.; Ye, C.; Wan, K.; Feng, M.; Luo, C.; Yu, X. Higher sensitivity to Cu2+ exposure of Microcystis aeruginosa in late lag phase is beneficial to its control. Water Res. 2022, 214, 118207. [Google Scholar] [CrossRef]
- Facey, J.A.; Violi, J.P.; King, J.J.; Sarowar, C.; Apte, S.C.; Mitrovic, S.M. The Influence of Micronutrient Trace Metals on Microcystis aeruginosa Growth and Toxin Production. Toxins 2022, 14, 812. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Huang, Y.; Ma, J.; Zhang, S.; Liu, J.; Li, T.; Song, L. Toxicity of the disinfectant benzalkonium chloride (C14) towards cyanobacterium Microcystis results from its impact on the photosynthetic apparatus and cell metabolism. J. Environ. Sci. 2024, 135, 198–209. [Google Scholar] [CrossRef]
- Tsai, K.-P. Effects of two copper compounds on Microcystis aeruginosa cell density, membrane integrity, and microcystin release. Ecotoxicol. Environ. Saf. 2015, 120, 428–435. [Google Scholar] [CrossRef]
- Wu, H.; Wei, G.; Tan, X.; Li, L.; Li, M. Species-dependent variation in sensitivity of Microcystis species to copper sulfate: Implication in algal toxicity of copper and controls of blooms. Sci. Rep. 2017, 7, 40393. [Google Scholar] [CrossRef]
- Iwinski, K.J.; Calomeni, A.J.; Geer, T.D.; Rodgers, J.H., Jr. Cellular and aqueous microcystin-LR following laboratory exposures of Microcystis aeruginosa to copper algaecides. Chemosphere 2016, 147, 74–81. [Google Scholar] [CrossRef]
- García-Villada, L.; Rico, M.; Altamirano, M.; Sánchez-Martín, L.; López-Rodas, V.; Costas, E. Occurrence of copper resistant mutants in the toxic cyanobacteria Microcystis aeruginosa: Characterisation and future implications in the use of copper sulphate as algaecide. Water Res. 2004, 38, 2207–2213. [Google Scholar] [CrossRef] [PubMed]
- Aragão, M.C.; dos Reis, K.C.; Rocha, M.A.M.; Guedes, D.d.O.; dos Santos, E.C.; Capelo-Neto, J. Removal of Dolichospermum circinale, Microcystis aeruginosa, and their metabolites using hydrogen peroxide and visible light. Aquat. Toxicol. 2021, 232, 105735. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Chen, H.; Wen, Y. Effect of hydrogen peroxide on Microcystic aeruginosa: Role of cytochromes P450. Sci. Total Environ. 2018, 626, 211–218. [Google Scholar] [CrossRef]
- Li, D.; Kang, X.; Chu, L.; Wang, Y.; Song, X.; Zhao, X.; Cao, X. Algicidal mechanism of Raoultella ornithinolytica against Microcystis aeruginosa: Antioxidant response, photosynthetic system damage and microcystin degradation. Environ. Pollut. 2021, 287, 117644. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X.; Jiang, X.; Liu, L.; Wang, H. Algicidal activity of Aspergillus niger induced by calcium ion as signal molecule on Microcystis aeruginosa. Algal Res. 2021, 60, 102536. [Google Scholar] [CrossRef]
- Kong, Y.; Ji, L.; Wang, Y.; Li, J.; Lu, H.; Mo, S.; Wang, X.; Zhu, L.; Xu, X.; Zheng, X. Combined Effect of NZVI and H2O2 on the Cyanobacterium Microcystis aeruginosa: Performance and Mechanism. Nanomaterials 2022, 12, 3017. [Google Scholar] [CrossRef] [PubMed]
- Anam, G.B.; Guda, D.R.; Ahn, Y.-H. Impact of melatonin on the hydrogen peroxide treatment efficacy in Microcystis aeruginosa: Cell growth, oxidative stress response, and gene transcription. Chemosphere 2022, 307, 136036. [Google Scholar] [CrossRef] [PubMed]
- Kansole, M.M.R.; Lin, T.-F. Impacts of Hydrogen Peroxide and Copper Sulfate on the Control of Microcystis aeruginosa and MC-LR and the Inhibition of MC-LR Degrading Bacterium Bacillus sp. Water 2017, 9, 255. [Google Scholar] [CrossRef]
- Ding, Y.; Gan, N.; Li, J.; Sedmak, B.; Song, L. Hydrogen peroxide induces apoptotic-like cell death in Microcystis aeruginosa (Chroococcales, Cyanobacteria) in a dose-dependent manner. Phycologia 2012, 51, 567–575. [Google Scholar] [CrossRef]
- Huo, X.; Chang, D.-W.; Tseng, J.-H.; Burch, M.D.; Lin, T.-F. Exposure of Microcystis aeruginosa to Hydrogen Peroxide under Light: Kinetic Modeling of Cell Rupture and Simultaneous Microcystin Degradation. Environ. Sci. Technol. 2015, 49, 5502–5510. [Google Scholar] [CrossRef]
- Lürling, M.; Meng, D.; Faassen, E.J. Effects of Hydrogen Peroxide and Ultrasound on Biomass Reduction and Toxin Release in the Cyanobacterium, Microcystis aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef] [PubMed]
- Nam, G.; Jeon, M.-S.; Choi, Y.-E.; Jung, J. Toxicity Reduction of Microcystis Aeruginosa Using Microbubble Ozonation. Ozone Sci. Eng. 2022, 45, 182–190. [Google Scholar] [CrossRef]
- Coral, L.A.; Zamyadi, A.; Barbeau, B.; Bassetti, F.J.; Lapolli, F.R.; Prévost, M. Oxidation of Microcystis aeruginosa and Anabaena flos-aquae by ozone: Impacts on cell integrity and chlorination by-product formation. Water Res. 2013, 47, 2983–2994. [Google Scholar] [CrossRef]
- Gu, J.; Huang, Z.; Fan, H.; Jin, Z.; Yan, Z.; Zhang, J. Mineralogy, geochemistry, and genesis of lateritic bauxite deposits in the Wuchuan–Zheng’an–Daozhen area, Northern Guizhou Province, China. J. Geochem. Explor. 2013, 130, 44–59. [Google Scholar] [CrossRef]
- Nam, G.; Mohamed, M.M.; Jung, J. Novel treatment of Microcystis aeruginosa using chitosan-modified nanobubbles. Environ. Pollut. 2021, 292, 118458. [Google Scholar] [CrossRef]
- Pei, H.-Y.; Ma, C.-X.; Hu, W.-R.; Sun, F. The behaviors of Microcystis aeruginosa cells and extracellular microcystins during chitosan flocculation and flocs storage processes. Bioresour. Technol. 2014, 151, 314–322. [Google Scholar] [CrossRef]
- Cao, J.; Zhao, H.; Dong, Z.; Yang, Z. Allelopathic effect of Chitosan Fiber on the growth of Microcystis aeruginosa. E3S Web Conf. 2020, 206, 02012. [Google Scholar] [CrossRef]
- Zhao, X.; Zeng, S.; Feng, H.; Wang, Y.; Li, S.; Zhou, X.; Wang, M.; Rei, L. Antifouling performance of in situ synthesized chitosan-zinc oxide hydrogel film against alga M. aeruginosa. Int. J. Biol. Macromol. 2022, 200, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Hu, W.; Pei, H.; Xu, H.; Pei, R. Enhancing integrated removal of Microcystis aeruginosa and adsorption of microcystins using chitosan-aluminum chloride combined coagulants: Effect of chemical dosing orders and coagulation mechanisms. Colloids Surf. A Physicochem. Eng. Asp. 2016, 490, 258–267. [Google Scholar] [CrossRef]
- Villar-Navarro, E.; Levchuk, I.; Rueda-Márquez, J.J.; Manzano, M. Combination of solar disinfection (SODIS) with H2O2 for enhanced disinfection of marine aquaculture effluents. Sol. Energy 2019, 177, 144–154. [Google Scholar] [CrossRef]
- Folt, C.L.; Chen, C.Y.; Moore, M.V.; Burnaford, J. Synergism and antagonism among multiple stressors. Limnol. Oceanogr. 1999, 44, 864–877. [Google Scholar] [CrossRef]
- Jackson, M.C.; Loewen, C.J.; Vinebrooke, R.D.; Chimimba, C.T. Net effects of multiple stressors in freshwater ecosystems: A meta-analysis. Glob. Chang. Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef]
- Schinegger, R.; Palt, M.; Segurado, P.; Schmutz, S. Untangling the effects of multiple human stressors and their impacts on fish assemblages in European running waters. Sci. Total Environ. 2016, 573, 1079–1088. [Google Scholar] [CrossRef]
- Gumbo, R.J.; Ross, G.; Cloete, E.T. Biological control of Microcystis dominated harmful algal blooms. Afr. J. Biotechnol. 2008, 7, 4765–4773. [Google Scholar]
- Kim, B.-H.; Sang, M.; Hwang, S.-J.; Han, M.-S. In situ bacterial mitigation of the toxic cyanobacterium Microcystis aeruginosa: Implications for biological bloom control. Limnol. Oceanogr. Methods 2008, 6, 513–522. [Google Scholar] [CrossRef]
- Zhao, W.; Zheng, Z.; Zhang, J.; Roger, S.-F.; Luo, X. Allelopathically inhibitory effects of eucalyptus extracts on the growth of Microcystis aeruginosa. Chemosphere 2019, 225, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, P.; Wu, X.-P. Grazing on toxic and non-toxic Microcystis aeruginosa PCC7820 by Unio douglasiae and Corbicula fluminea. Limnology 2009, 10, 1–5. [Google Scholar] [CrossRef]
- Han, Y.; Zheng, J.; Jiang, C.; Zhang, F.; Wei, L.; Zhu, L. Hydrochloric acid-modified algal biochar for the removal of Microcystis aeruginosa: Coagulation performance and mechanism. J. Environ. Chem. Eng. 2022, 10, 108903. [Google Scholar] [CrossRef]
- Kim, B.-H.; Kang, Y.-H.; Choi, H.-J.; Ka, S.-K.; Han, M.-S. Environment. Antialgal interactions of biological control agents on cyanobacterium and diatom blooms in vitro. Korean J. Ecol. Environ. 2005, 38, 494–502. [Google Scholar]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae–bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Gerphagnon, M.; Macarthur, D.J.; Latour, D.; Gachon, C.M.M.; Van Ogtrop, F.; Gleason, F.H.; Sime-Ngando, T. Microbial players involved in the decline of filamentous and colonial cyanobacterial blooms with a focus on fungal parasitism. Environ. Microbiol. 2015, 17, 2573–2587. [Google Scholar] [CrossRef]
- Park, B.S.; Park, C.-S.; Shin, Y.; Yoon, S.; Han, M.-S.; Kang, Y.-H. Different Algicidal Modes of the Two Bacteria Aeromonas bestiarum HYD0802-MK36 and Pseudomonas syringae KACC10292T against Harmful Cyanobacteria Microcystis aeruginosa. Toxins 2022, 14, 128. [Google Scholar] [CrossRef]
- Das Nishu, S.; Kang, Y.; Han, I.; Jung, T.Y.; Lee, T.K. Nutritional status regulates algicidal activity of Aeromonas sp. L23 against cyanobacteria and green algae. PLoS ONE 2019, 14, e0213370. [Google Scholar] [CrossRef]
- Zhou, S.; Yin, H.; Tang, S.; Peng, H.; Yin, D.; Yang, Y.; Liu, Z.; Dang, Z. Physiological responses of Microcystis aeruginosa against the algicidal bacterium Pseudomonas aeruginosa. Ecotoxicol. Environ. Saf. 2016, 127, 214–221. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Xu, Y.; Li, P.; Zhang, K.; Jiang, X.; Zheng, T.; Wang, H. Stress of algicidal substances from a bacterium Exiguobacterium sp. h10 on Microcystis aeruginosa. Lett. Appl. Microbiol. 2017, 64, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Wang, Y.; Tang, S.; Liang, J.; Lin, W.; Luo, L. Isolation and Identification of Algicidal Compound from Streptomyces and Algicidal Mechanism to Microcystis aeruginosa. PLoS ONE 2013, 8, e76444. [Google Scholar] [CrossRef]
- Caiola, M.G.; Pellegrini, S. Lysis of microcystis aeruginosa (kütz.) By bdellovibrio-like bacteria. J. Phycol. 1984, 20, 471–475. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Yang, L.; Chen, H.; Zhang, S.; Zhao, H.; Zhang, N. Allelopathic control of cyanobacterial blooms by periphyton biofilms. Environ. Microbiol. 2011, 13, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Jianhong, Q.; Shaobin, L. The growth of Bacillus sp. and Microcystis aeruginosa and their competition for resources. J. Zhanjiang Ocean. Univ. 2002, 22, 13–18. [Google Scholar]
- Zhang, C.; Massey, I.Y.; Liu, Y.; Huang, F.; Gao, R.; Ding, M.; Xiang, L.; He, C.; Wei, J.; Li, Y.; et al. Identification and characterization of a novel indigenous algicidal bacterium Chryseobacterium species against Microcystis aeruginosa. J. Toxicol. Environ. Health Part A 2019, 82, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ye, Q.; Zhang, F.; Shao, X.; Fan, Y.; Zhu, X.; Li, Y.; Yao, L.; Tian, Y.; Zheng, T.; et al. Flocculating properties and potential of Halobacillus sp. strain H9 for the mitigation of Microcystis aeruginosa blooms. Chemosphere 2019, 218, 138–146. [Google Scholar] [CrossRef]
- Gumbo, J.; Cloete, T. The mechanism of Microcystis aeruginosa death upon exposure to Bacillus mycoides. Phys. Chem. Earth Parts A/B/C 2011, 36, 881–886. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Hu, Z.; Chen, D.; Li, F.; Huang, X.; Li, C. Transcriptome Analysis Reveals the Algicidal Mechanism of Brevibacillus laterosporus against Microcystis aeruginosa through Multiple Metabolic Pathways. Toxins 2022, 14, 492. [Google Scholar] [CrossRef]
- Jing, W.; Sui, G.; Liu, S. Characteristics of a Microcystin-LR Biodegrading Bacterial Isolate: Ochrobactrum sp. FDT5. Bull. Environ. Contam. Toxicol. 2014, 92, 119–122. [Google Scholar] [CrossRef]
- Mu, R.; He, Y.; Liu, S.; Wang, X.; Fan, Z. The Algicidal Characteristics of One Algae-Lysing FDT5 Bacterium on Microcystis aeruginosa. Geomicrobiol. J. 2009, 26, 516–521. [Google Scholar] [CrossRef]
- Sun, P.; Lin, H.; Wang, G.; Zhang, X.; Zhang, Q.; Zhao, Y. Wheat Bran Enhances the Cytotoxicity of Immobilized Alcaligenes aquatilis F8 against Microcystis aeruginosa. PLoS ONE 2015, 10, e0136429. [Google Scholar] [CrossRef]
- Xuan, H.; Dai, X.; Li, J.; Zhang, X.; Yang, C.; Luo, F. A Bacillus sp. strain with antagonistic activity against Fusarium graminearum kills Microcystis aeruginosa selectively. Sci. Total Environ. 2017, 583, 214–221. [Google Scholar] [CrossRef]
- Benegas, G.R.S.; Bernal, S.P.F.; de Oliveira, V.M.; Passarini, M.R.Z. Antimicrobial activity against Microcystis aeruginosa and degradation of microcystin-LR by bacteria isolated from Antarctica. Environ. Sci. Pollut. Res. 2021, 28, 52381–52391. [Google Scholar] [CrossRef]
- Liu, Z.Z.; Zhu, J.P.; Li, M.; Xue, Q.Q.; Zeng, Y.; Wang, Z.P. Effects of freshwater bacterial siderophore on Microcystis and Anabaena. Biol. Control. 2014, 78, 42–48. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Z.; Huang, Q.; Xiao, X.; Wang, X.; Zhang, F.; Wang, X.; Liu, Y.; Hu, C. Isolation, identification and characterization of phytoplankton-lytic bacterium CH-22 against Microcystis aeruginosa. Limnologica 2011, 41, 70–77. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, J.; Yang, C.; Ding, M.; Hamilton, P.B.; Zhang, X.; Yang, C.; Zhnag, L.; Dai, X. A Streptomyces globisporus strain kills Microcystis aeruginosa via cell-to-cell contact. Sci. Total Environ. 2021, 769, 144489. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Purohit, H.J.; Qureshi, A. Genomic insight for algicidal activity in Rhizobium strain AQ_MP. Arch. Microbiol. 2021, 203, 5193–5203. [Google Scholar] [CrossRef]
- Manage, P.M.; Kawabata, Z.; Nakano, S.-I. Algicidal effect of the bacterium Alcaligenes denitrificans on Microcystis spp. Aquat. Microb. Ecol. 2000, 22, 111–117. [Google Scholar] [CrossRef]
- Yang, L.; Maeda, H.; Yoshikawa, T.; Zhou, G.-Q. Algicidal effect of bacterial isolates of Pedobacter sp. against cyanobacterium Microcystis aeruginosa. Water Sci. Eng. 2012, 5, 375–382. [Google Scholar]
- Zhang, B.-H.; Chen, W.; Li, H.-Q.; Zhou, E.-M.; Hu, W.-Y.; Duan, Y.-Q.; Mohamad, O.A.; Gao, R.; Li, W.-J. An antialgal compound produced by Streptomyces jiujiangensis JXJ 0074T. Appl. Microbiol. Biotechnol. 2015, 99, 7673–7683. [Google Scholar] [CrossRef] [PubMed]
- Su, J.F.; Shao, S.C.; Ma, F.; Lu, J.S.; Zhang, K. Bacteriological control by Raoultella sp. R11 on growth and toxins production of Microcystis aeruginosa. Chem. Eng. J. 2016, 293, 139–150. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Zhang, C.; Pan, Z.; Han, S. Synergistic removal of Microcystis aeruginosa by a novel H2O2 pre-oxidation enhanced pressurization method: Performance and mechanism. J. Clean. Prod. 2022, 379, 134745. [Google Scholar] [CrossRef]
- Kang, Y.-H.; Park, C.-S.; Han, M.-S. Pseudomonas aeruginosa UCBPP-PA14 a useful bacterium capable of lysing Microcystis aeruginosa cells and degrading microcystins. J. Appl. Phycol. 2012, 24, 1517–1525. [Google Scholar] [CrossRef]
- Yi, Y.-L.; Yu, X.-B.; Zhang, C.; Wang, G.-X. Growth inhibition and microcystin degradation effects of Acinetobacter guillouiae A2 on Microcystis aeruginosa. Res. Microbiol. 2015, 166, 93–101. [Google Scholar] [CrossRef]
- Santos, A.A.; Soldatou, S.; de Magalhães, V.F.; Azevedo, S.M.F.O.; Camacho-Muñoz, D.; Lawton, L.A.; Edwards, C. Degradation of Multiple Peptides by Microcystin-Degrader Paucibacter toxinivorans (2C20). Toxins 2021, 13, 265. [Google Scholar] [CrossRef]
- Crettaz-Minaglia, M.; Fallico, M.; Aranda, O.; Juarez, I.; Pezzoni, M.; Costa, C.; Andrinolo, D.; Giannuzzi, L. Effect of temperature on microcystin-LR removal and lysis activity on Microcystis aeruginosa (cyanobacteria) by an indigenous bacterium belonging to the genus Achromobacter. Environ. Sci. Pollut. Res. 2020, 27, 44427–44439. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Tan, J.; Lin, S.; Li, D.; Yang, H. Isolation, identification and characterization of an algicidal bacterium from Lake Taihu and preliminary studies on its algicidal compounds. J. Environ. Sci. 2012, 24, 1823–1831. [Google Scholar] [CrossRef]
- Shao, J.; He, Y.; Chen, A.; Peng, L.; Luo, S.; Wu, G.; Zou, H.; Li, R. Interactive effects of algicidal efficiency of Bacillus sp. B50 and bacterial community on susceptibility of Microcystis aeruginosa with different growth rates. Int. Biodeterior. Biodegrad. 2015, 97, 1–6. [Google Scholar] [CrossRef]
- Yang, C.; Hou, X.; Wu, D.; Chang, W.; Zhang, X.; Dai, X.; Du, H.; Zhang, X.; Igarashi, Y.; Luo, F. The characteristics and algicidal mechanisms of cyanobactericidal bacteria, a review. World J. Microbiol. Biotechnol. 2020, 36, 188. [Google Scholar] [CrossRef]
- Yang, J.; Qiao, K.; Lv, J.; Liu, Q.; Nan, F.; Xie, S.; Feng, J. Isolation and Identification of Two Algae-Lysing Bacteria against Microcystis aeruginosa. Water 2020, 12, 2485. [Google Scholar] [CrossRef]
- Sun, P.; Esquivel-Elizondo, S.; Zhao, Y.; Wu, Y. Glucose triggers the cytotoxicity of Citrobacter sp. R1 against Microcystis aeruginosa. Sci. Total Environ. 2017, 603–604, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lyu, P.; Li, H.; Zheng, X.; Zhang, H.; Wang, C.; Qin, Y.; Xia, B.; Wang, D.; Xu, S.; Zhuang, X. Oxidative stress of Microcystis aeruginosa induced by algicidal bacterium Stenotrophomonas sp. KT48. Appl. Microbiol. Biotechnol. 2022, 106, 4329–4340. [Google Scholar] [CrossRef]
- Zhang, S.; He, X.; Cao, L.; Tong, Y.; Zhao, B.; An, W. A Novel Wide-Range Freshwater Cyanophage MinS1 Infecting the Harmful Cyanobacterium Microcystis aeruginosa. Viruses 2022, 14, 433. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, X. High-Cell-Density Cultivation and Algicidal Activity Assays of a Novel Algicidal Bacterium to Control Algal Bloom Caused by Water Eutrophication. Water Air Soil Pollut. 2014, 225, 2120. [Google Scholar] [CrossRef]
- Xue, G.; Wang, X.; Xu, C.; Song, B.; Chen, H. Removal of harmful algae by Shigella sp. H3 and Alcaligenes sp. H5: Algicidal pathways and characteristics. Environ. Technol. 2022, 43, 4341–4353. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Han, Z.; Ye, J.; Liu, Z. The influence of aquatic macrophytes on Microcystis aeruginosa growth. Ecol. Eng. 2012, 42, 130–133. [Google Scholar] [CrossRef]
- Guo, X.; Liu, X.; Pan, J.; Yang, H. Synergistic algicidal effect and mechanism of two diketopiperazines produced by Chryseobacterium sp. strain GLY-1106 on the harmful bloom-forming Microcystis aeruginosa. Sci. Rep. 2015, 5, 14720. [Google Scholar] [CrossRef]
- Furusawa, G.; Iwamoto, K. Removal of Microcystis aeruginosa cells using the dead cells of a marine filamentous bacterium, Aureispira sp. CCB-QB1. PeerJ 2022, 10, e12867. [Google Scholar] [CrossRef]
- Phankhajon, K.; Somdee, A.; Somdee, T. Algicidal activity of an actinomycete strain, Streptomyces rameus, against Microcystis aeruginosa. Water Sci. Technol. 2016, 74, 1398–1408. [Google Scholar] [CrossRef]
- Somdee, T.; Sumalai, N.; Somdee, A. A novel actinomycete Streptomyces aurantiogriseus with algicidal activity against the toxic cyanobacterium Microcystis aeruginosa. J. Appl. Phycol. 2013, 25, 1587–1594. [Google Scholar] [CrossRef]
- Yu, Y.; Zeng, Y.; Li, J.; Yang, C.; Zhang, X.; Luo, F.; Dai, X. An algicidal Streptomyces amritsarensis strain against Microcystis aeruginosa strongly inhibits microcystin synthesis simultaneously. Sci. Total Environ. 2019, 650, 34–43. [Google Scholar] [CrossRef]
- Zhang, B.-H.; Chen, W.; Li, H.-Q.; Yang, J.-Y.; Zha, D.-M.; Duan, Y.-Q.; Hozzein, N.W.; Xiao, M.; Gao, R.; Li, W.-J. L-valine, an antialgal amino acid from Streptomyces jiujiangensis JXJ 0074T. Appl. Microbiol. Biotechnol. 2016, 100, 4627–4636. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-H.; Peng, P.; Liu, Y.-M.; Jia, R.-B.; Li, L. Algicidal activity of a dibenzofuran-degrader Rhodococcus sp. J. Microbiol. Biotechnol. 2013, 23, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Kovalerchick, D.; Lieman-Hurwitz, J.; Murik, O.; De Philippis, R.; Carmeli, S.; Sukenik, A.; Kaplan, A. Increased algicidal activity of Aeromonas veronii in response to Microcystis aeruginosa: Interspecies crosstalk and secondary metabolites synergism. Environ. Microbiol. 2019, 21, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Mu, R.-M.; Fan, Z.-Q.; Pei, H.-Y.; Yuan, X.-L.; Liu, S.-X.; Wang, X.-R. Isolation and algae-lysing characteristics of the algicidal bacterium B5. J. Environ. Sci. 2007, 19, 1336–1340. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Chi, Y.; Wu, D.; Dai, X.; Zhang, X.; Igarashi, Y.; Luo, F. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake. J. Basic Microbiol. 2019, 59, 1112–1124. [Google Scholar] [CrossRef]
- Sun, P.; Hui, C.; Wang, S.; Khan, R.A.; Zhang, Q.; Zhao, Y.-H. Enhancement of algicidal properties of immobilized Bacillus methylotrophicus ZJU by coating with magnetic Fe3O4 nanoparticles and wheat bran. J. Hazard. Mater. 2016, 301, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, M.; Hong, M.; Park, W. Killing effect of deinoxanthins on cyanobloom-forming Microcystis aeruginosa: Eco-friendly production and specific activity of deinoxanthins. Environ. Res. 2021, 200, 111455. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Alamri, S.; Hashem, M.; Mostafa, Y. Growth inhibition of Microcystis aeruginosa and adsorption of microcystin toxin by the yeast Aureobasidium pullulans, with no effect on microalgae. Environ. Sci. Pollut. Res. 2020, 27, 38038–38046. [Google Scholar] [CrossRef]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial activity and bacterial community structure during degradation of microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef]
- Kinley, C.M.; Iwinski-Wood, K.J.; Geer, T.D.; Hendrikse, M.; McQueen, A.D.; Calomeni, A.J.; Liang, J.; Friesen, V.; Simair, M.C.; Rodgers, J.H. Microcystin-LR Degradation Following Copper-Based Algaecide Exposures. Water Air Soil Pollut. 2018, 229, 62. [Google Scholar] [CrossRef]
- Shibata, K.; Amemiya, T.; Itoh, K. Activities of Algicidal Bacteria and Their Influences on Microbial Communities. 2008. Available online: https://www.researchgate.net/publication/37570727_Activities_of_Algicidal_Bacteria_and_Their_Influences_on_Microbial_Communities (accessed on 1 April 2023).
- Takamura, Y.; Yamada, T.; Kimoto, A.; Kanehama, N.; Tanaka, T.; Nakadaira, S.; Yagi, O. Growth Inhibition of Microcystis Cyanobacteria by L-Lysine and Disappearance of Natural Microcystis Blooms with Spraying. Microbes Environ. 2004, 19, 31–39. [Google Scholar] [CrossRef]
- Zeng, G.; Gao, P.; Wang, J.; Zhang, J.; Zhang, M.; Sun, D. Algicidal Molecular Mechanism and Toxicological Degradation of Microcystis aeruginosa by White-Rot Fungi. Toxins 2020, 12, 406. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Hashem, M.; Alamri, S.A. Growth inhibition of the cyanobacterium Microcystis aeruginosa and degradation of its microcystin toxins by the fungus Trichoderma citrinoviride. Toxicon 2014, 86, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Du, J.; Song, F.; Zhao, G.; Tian, X. A Fungus Capable of Degrading Microcystin-LR in the Algal Culture of Microcystis aeruginosa PCC7806. Appl. Biochem. Biotechnol. 2012, 166, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Ma, H.; Ren, S.; Gao, X.; He, X.; Zhu, S.; Deng, R.; Zhang, S. Insights into the mechanism of cyanobacteria removal by the algicidal fungi Bjerkandera adusta and Trametes versicolor. Microbiologyopen 2020, 9, e1042. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Feng, X.; Jia, Y.; Wang, C.; He, X.; Zhou, Q.; Tian, X. Isolation and evaluation of terrestrial fungi with algicidal ability from Zijin Mountain, Nanjing, China. J. Microbiol. 2011, 49, 562–567. [Google Scholar] [CrossRef]
- Sun, R.; Sun, P.; Zhang, J.; Esquivel-Elizondo, S.; Wu, Y. Microorganisms-based methods for harmful algal blooms control: A review. Bioresour. Technol. 2018, 248, 12–20. [Google Scholar] [CrossRef]
- Gao, X.; Wang, C.; Dai, W.; Ren, S.; Tao, F.; He, X.; Han, G.; Wang, W. Proteomic analysis reveals large amounts of decomposition enzymes and major metabolic pathways involved in algicidal process of Trametes versicolor F21a. Sci. Rep. 2017, 7, 3907. [Google Scholar] [CrossRef]
- Jia, Y.; Han, G.; Wang, C.; Guo, P.; Jiang, W.; Li, X.; Tian, X. The efficacy and mechanisms of fungal suppression of freshwater harmful algal bloom species. J. Hazard. Mater. 2010, 183, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhou, Q.; Lilje, O.; Xu, W.; Zhu, Y.; van Ogtrop, F.F. Inhibition mechanism of Penicillium chrysogenum on Microcystis aeruginosa in aquaculture water. J. Clean. Prod. 2021, 299, 126829. [Google Scholar] [CrossRef]
- Gerphagnon, M.; Latour, D.; Colombet, J.; Sime-Ngando, T. Fungal Parasitism: Life Cycle, Dynamics and Impact on Cyanobacterial Blooms. PLoS ONE 2013, 8, e60894. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W.; De Bruin, A.; Kagami, M.; Rijkeboer, M.; Brehm, M.; Van Donk, E. Host parasite interactions between freshwater phytoplankton and chytrid fungi (chytridiomycota). J. Phycol. 2004, 40, 437–453. [Google Scholar] [CrossRef]
- Mountfort, D.O.; Atkinson, M.; Ponikla, K.; Burke, B.; Todd, K. Lysis of Gymnodinium Species by the Fungus Verticillium lecanii. Bot. Mar. 1996, 39, 159–166. [Google Scholar] [CrossRef]
- Canter, H.M.; Lund, J. Studies on plankton parasites: II. The parasitism of diatoms with special reference to lakes in the English Lake District. Trans. Br. Mycol. Soc. 1953, 36, 13–37. [Google Scholar] [CrossRef]
- Van Donk, E. The role of fungal parasites in phytoplankton succession. Plankton Ecol. Succession Plankton Communities 1989, 171–194. [Google Scholar] [CrossRef]
- Kudoh, S.; Tokahashi, M. Fungal control of population changes of the planktonic diatom asterionella formosa in a shallow eutrophic lake. J. Phycol. 1990, 26, 239–244. [Google Scholar] [CrossRef]
- Sime-Ngando, T. Phytoplankton chytridiomycosis: Fungal parasites of phytoplankton and their imprints on the food web dynamics. Front. Microbiol. 2012, 3, 361. [Google Scholar] [CrossRef]
- Tucker, S.; Pollard, P. Identification of Cyanophage Ma-LBP and Infection of the Cyanobacterium Microcystis aeruginosa from an Australian Subtropical Lake by the Virus. Appl. Environ. Microbiol. 2005, 71, 629–635. [Google Scholar] [CrossRef]
- Mankiewicz-Boczek, J.; Jaskulska, A.; Pawełczyk, J.; Gągała, I.; Serwecińska, L.; Dziadek, J. Cyanophages infection of Microcystis bloom in lowland dam reservoir of Sulejów, Poland. Microb. Ecol. 2016, 71, 315–325. [Google Scholar] [CrossRef]
- Pollard, P.C.; Young, L.M. Lake viruses lyse cyanobacteria, Cylindrospermopsis raciborskii, enhances filamentous-host dispersal in Australia. Acta Oecol. 2010, 36, 114–119. [Google Scholar] [CrossRef]
- Yoshida, T.; Takashima, Y.; Tomaru, Y.; Shirai, Y.; Takao, Y.; Hiroishi, S.; Nagasaki, K. Isolation and Characterization of a Cyanophage Infecting the Toxic Cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 2006, 72, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Manage, P.M.; Kawabata, Z.; Nakano, S. Dynamics of cyanophage-like particles and algicidal bacteria causing Microcystis aeruginosa mortality. Limnology 2001, 2, 73–78. [Google Scholar] [CrossRef]
- Wang, F.; Li, D.; Cai, R.; Pan, L.; Zhou, Q.; Liu, W.; Qian, M.; Tong, Y. A Novel Freshwater Cyanophage Mae-Yong1326-1 Infecting Bloom-Forming Cyanobacterium Microcystis aeruginosa. Viruses 2022, 14, 2051. [Google Scholar] [CrossRef]
- Long, A.M. Persistence of Algal Viruses and Cyanophages in Freshwater Environments; University of Toronto (Canada): Toronto, ON, Canada, 2017. [Google Scholar]
- Pal, M.; Yesankar, P.J.; Dwivedi, A.; Qureshi, A. Biotic control of harmful algal blooms (HABs): A brief review. J. Environ. Manag. 2020, 268, 110687. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, Y.; Xie, W.; He, W.; Xie, J.; Liu, W. Identifying Algicides of Enterobacter hormaechei F2 for Control of the Harmful Alga Microcystis aeruginosa. Int. J. Environ. Res. Public Health 2022, 19, 7556. [Google Scholar] [CrossRef]
- Suttle, C.A. The Ecological, Evolutionary and Geochemical Consequences of Viral Infection of Cyanobacteria and Eukaryotic Algae. Available online: https://www.researchgate.net/publication/258503632_The_Ecological_Evolutionary_and_Geochemical_Consequences_of_Viral_Infection_of_Cyanobacteria_and_Eukaryotic_Algae (accessed on 1 April 2023).
- Short, S.M. The ecology of viruses that infect eukaryotic algae. Environ. Microbiol. 2012, 14, 2253–2271. [Google Scholar] [CrossRef]
- Ben Gharbia, H.; Kéfi-Daly Yahia, O.; Cecchi, P.; Masseret, E.; Amzil, Z.; Herve, F.; Rovillon, G.; Nouri, H.; M’rabet, C.; Couet, D. New insights on the species-specific allelopathic interactions between macrophytes and marine HAB dinoflagellates. PLoS ONE 2017, 12, e0187963. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Zhang, Z. Potential applications of alginate oligosaccharides for biomedicine—A mini review. Carbohydr. Polym. 2021, 271, 118408. [Google Scholar] [CrossRef] [PubMed]
- Asselman, J.; Hochmuth, J.; De Schamphelaere, K. A comparison of the sensitivities of Daphnia magna and Daphnia pulex to six different cyanobacteria. Harmful Algae 2014, 39, 1–7. [Google Scholar] [CrossRef]
- Ger, K.A.; Hansson, L.-A.; Lürling, M. Understanding cyanobacteria-zooplankton interactions in a more eutrophic world. Freshw. Biol. 2014, 59, 1783–1798. [Google Scholar] [CrossRef]
- Wilson, A.E.; Sarnelle, O.; Tillmanns, A.R. Effects of cyanobacterial toxicity and morphology on the population growth of freshwater zooplankton: Meta-analyses of laboratory experiments. Limnol. Oceanogr. 2006, 51, 1915–1924. [Google Scholar] [CrossRef]
- Savic, G.B.; Edwards, C.; Briand, E.; Lawton, L.; Wiegand, C.; Bormans, M. Daphnia magna Exudates Impact Physiological and Metabolic Changes in Microcystis aeruginosa. Toxins 2019, 11, 421. [Google Scholar] [CrossRef]
- Ger, K.A.; Panosso, R.; Lürling, M. Consequences of acclimation to Microcystis on the selective feeding behavior of the calanoid copepod Eudiaptomus gracilis. Limnol. Oceanogr. 2011, 56, 2103–2114. [Google Scholar] [CrossRef]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.-A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The interaction between cyanobacteria and zooplankton in a more eutrophic world. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef]
- Jang, M.-H.; Jung, J.-M.; Takamura, N. Changes in microcystin production in cyanobacteria exposed to zooplankton at different population densities and infochemical concentrations. Limnol. Oceanogr. 2007, 52, 1454–1466. [Google Scholar] [CrossRef]
- Van Gremberghe, I.; Vanormelingen, P.; Vanelslander, B.; Van der Gucht, K.; D’hondt, S.; De Meester, L.; Vyverman, W. Genotype-dependent interactions among sympatric Microcystis strains mediated by Daphnia grazing. Oikos 2009, 118, 1647–1658. [Google Scholar] [CrossRef]
- Van Donk, E. Chemical information transfer in freshwater plankton. Ecol. Inform. 2007, 2, 112–120. [Google Scholar] [CrossRef]
- Shapiro, J. Biomanipulation: An ecosystem approach to lake restoration. In Proceedings of the Symposium on Water Quality Management through Biological Control, Gainesville, FL, USA, 23–30 January 1975; pp. 85–95. [Google Scholar]
- Starling, F. Control of eutrophication by silver carp in the tropical Pranoa Resevoir, Brazil: A mesocosm experiment. Hydrobiologia 1993, 257, 143–151. [Google Scholar] [CrossRef]
- Turker, H.; Eversole, A.G.; Brune, D.E. Filtration of green algae and cyanobacteria by Nile tilapia, Oreochromis niloticus, in the Partitioned Aquaculture System. Aquaculture 2003, 215, 93–101. [Google Scholar] [CrossRef]
- Lu, K.; Jin, C.; Dong, S.; Gu, B.; Bowen, S.H. Feeding and control of blue-green algal blooms by tilapia (Oreochromis Niloticus). Hydrobiologia 2006, 568, 111–120. [Google Scholar] [CrossRef]
- Torres, G.S.; Silva, L.H.S.; Rangel, L.M.; Attayde, J.L.; Huszar, V.L.M. Cyanobacteria are controlled by omnivorous filter-feeding fish (Nile tilapia) in a tropical eutrophic reservoir. Hydrobiologia 2016, 765, 115–129. [Google Scholar] [CrossRef]
- Görgényi, J.; Boros, G.; Vitál, Z.; Mozsár, A.; Várbíró, G.; Vasas, G.; Borics, G. The role of filter-feeding Asian carps in algal dispersion. Hydrobiologia 2016, 764, 115–126. [Google Scholar] [CrossRef]
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Tazart, Z.; Douma, M.; Tebaa, L.; Loudiki, M. Use of macrophytes allelopathy in the biocontrol of harmful Microcystis aeruginosa blooms. Water Supply 2019, 19, 245–253. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Growth inhibition of blue-green algae (Microcystis aeruginosa) by Myriophyllum spicatum-releasing four polyphenols. Water Res. 2000, 34, 3026–3032. [Google Scholar] [CrossRef]
- Tan, K.; Huang, Z.; Ji, R.; Qiu, Y.; Wang, Z.; Liu, J. A review of allelopathy on microalgae. Microbiology 2019, 165, 587–592. [Google Scholar] [CrossRef]
- Nakai, S.; Yamada, S.; Hosomi, M. Anti-cyanobacterial fatty acids released from Myriophyllum spicatum. Hydrobiologia 2005, 543, 71–78. [Google Scholar] [CrossRef]
- Kong, C.H.; Wang, P.; Zhang, C.X.; Zhang, M.X.; Hu, F. Herbicidal potential of allelochemicals from Lantana camara against Eichhornia crassipes and the alga Microcystis aeruginosa. Weed Res. 2006, 46, 290–295. [Google Scholar] [CrossRef]
- Wang, R.; Hua, M.; Yu, Y.; Zhang, M.; Xian, Q.-M.; Yin, D.-Q. Evaluating the effects of allelochemical ferulic acid on Microcystis aeruginosa by pulse-amplitude-modulated (PAM) fluorometry and flow cytometry. Chemosphere 2016, 147, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A review on control of harmful algal blooms by plant-derived allelochemicals. J. Hazard. Mater. 2021, 401, 123403. [Google Scholar] [CrossRef]
- Shao, J.; Yu, G.; Wang, Z.; Wu, Z.; Peng, X.; Li, R. Towards clarification of the inhibitory mechanism of wheat bran leachate on Microcystis aeruginosa NIES-843 (cyanobacteria): Physiological responses. Ecotoxicology 2010, 19, 1634–1641. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.-H.; Wu, X.-Q.; Li, R.-H. Physiological responses of Microcystis aeruginosa PCC7806 to nonanoic acid stress. Environ. Toxicol. 2009, 24, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Li, R.; Lepo, J.E.; Gu, J.-D. Potential for control of harmful cyanobacterial blooms using biologically derived substances: Problems and prospects. J. Environ. Manag. 2013, 125, 149–155. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).