Abstract
Tree residue removal from Eucalyptus plantations after timber harvest can reduce soil functioning by reducing the organic matter input. To assess the effects of residue management systems (RMS) on soil aggregation, carbon (C) and nitrogen (N) content, and biological activities, a field trial was conducted in a commercial Eucalyptus plantation (loamy sand soil) in Mato Grosso do Sul, Brazil. The study assessed three RMS: cut-to-length (CTL), tree-length (TL), and bare litter (BL), respectively. After 21 months, undisturbed soil samples were collected and physically isolated into three aggregate-size fractions: large macroaggregates (LM), medium macroaggregates (SM), and microaggregates (MI). Results showed that these soils are mostly composed of LM (54%), and that removing harvest residues from the growing site included total organic carbon (TOC) by 28%, microbial biomass-C by 20%, fluorescein diacetate hydrolysis by 17%, and β-glucosidase activity by 26%, when compared to CTL. TL outperformed CTL for the proportion of LM and LM-associated TOC. Across fractions, a higher microbial quotient was observed in SM and MI fractions, suggesting that the TOC has higher stability inside the LM. This study suggests that leaving harvest residues on the soil should be recommended for Eucalyptus plantations, especially in low-fertility sandy soils, as it helps in maintaining the soil structure and biological activities critical for soil health and ecosystem function.
1. Introduction
The global abundance of Eucalyptus plantations underscores their high species diversity and ecological adaptability, combined with their high production and rapid growth, even on sandy soils [1]. Such soils are typically infertile and cannot maintain adequate soil organic matter (SOM) levels [2], thereby requiring management practices that promote OM accumulation and biological activities as well as improve soil fertility [3,4,5].
For sustainable agriculture in sandy soils, physical and chemical properties can be preserved through reduced tillage or no-till systems, crop residue management, carbon inputs, and increased macro-aggregation and total porosity [2,6,7]. Understandably, harvesting methods can impose substantial differences in the amounts of OM deposited onto the soil surface.
The “cut-to-length” system dominates as the most common eucalyptus harvesting method in Brazil [8], returning the most tree residues to the soil as trees are delimbed in situ. By leaving all harvest residues on the ground, this boosts soil OM levels, biological activities, nutrient cycling, and soil structure. On the other hand, the industry has identified that removing some (tree-length) or all residues (bare litter) facilitates mechanical operations, reduces costs, and increases the operational efficiency of the business [8].
Tree-length harvest returns a moderate level of Eucalyptus residues to the soil, as they cut the trees and semi-process on-site to remove the pointer and bark, leaving on the field leaves, branches, and litter. The most intensive system, “bare litter”, removes all trees from the site to be delimbed outside of the cultivation area at processing facilities. By removing all harvest residues from the site, only litter fall remains on the forest floor through the current cycle.
When the non-commercial components of the eucalyptus logs are maintained in the field, nutrient cycling through litter decomposition and forest harvest residues give the soils a great potential to meet the tree’s nutritional demands, especially those of calcium and magnesium [4,9,10]. After harvesting, eucalyptus residues offer a source of OM and nutrients for the following eucalyptus growth cycles, apart from protecting the soil, increasing the water retention capacity, and preventing compaction, erosion, and direct sunlight exposure [11].
Organic matter improves soil aggregation, mainly in sandy soils, but soil aggregate stabilization is only possible through coupling organic material with mineral particles, which can be introduced by roots, fungal hyphae, or other microbial combinations [12,13]. A higher soil aggregate stability improves structural stability, and facilitates the movement of water, solutes, microorganisms, gases, and plant root growth, thereby benefiting the soil functions [14,15].
Biochemical attributes and microbial activities can serve as indicators of soil functioning. The microbial quotient (qMic) indicates the percentage of total organic carbon in the soil, while the metabolic quotient (qCO2) shows the specific respiration rates of microbial biomass [16]. In this way, stressed soils typically have higher qCO2 values [17], while areas with low organic compound stocks have corresponding low values of the microbial quotient [18].
Aside from the importance of maintaining harvest residues to ensure the entry of OM, it is also essential to maintain the soil structure, the carbon storage, and soil aggregates under different habitats (such as aerobic and anaerobic microsites) and support the activities of a diverse microbial community [19,20].
The aggregate size has a significant influence on microbiological activities, for example, in sandy soils under native pasture, basal respiration is higher in 250–4000 µm than in <250 µm aggregates [21]. Other studies have also stated increased basal respiration and microbial biomass carbon at greater soil aggregate sizes [22,23].
In this context, we hypothesize that residue management systems (RMSs) that maintain residual biomass including leaves, branches, and barks, on the soil surface after harvest, such as the cut-to-length (CTL) and tree-length (TL), contribute to improving the soil organic matter levels, biological activities, nutrient cycling, and soil structure, along with removing all harvest residues, such as in the bare litter method, reducing inputs of the organic material and consequently decreasing organic matter (OM) and nutrient contents. Thus, the RMSs CTL and TL, for increasing the OM and nutrient supply, are assumed to reduce the exposure and loss of C and N, which remain within the aggregates and stimulate microbial and enzymatic activities.
Given how fragile the structure of sandy soils is, and how relevant soil aggregates are for carbon storage and biological activities, the assessment of biological activities in soil aggregate samples is a relevant approach to demonstrate how small changes inside these aggregates due to management practices can significantly impact soil carbon and soil biology. The eucalyptus industry is increasingly opting for processing trees outside the growing site, resulting in no tree residues left on the soil. This practice reduces the amount of C input and nutrients that are returned to the soil. The objective of this research was to assess the impact of removing harvest residues from Eucalyptus harvest sites on soil health indicators assessed in soil aggregate samples, as compared to retaining tree residues on-site after harvest. Through this assessment of biological activities in soil aggregates, we aimed to provide new insights into the biological activity and soil C storage of sandy soils under tropical climate, and identify the best management practices to ensure the sustainability of the eucalyptus industry and the health of the surrounding ecosystems. Our field trial in a commercial Eucalyptus farm compared a removal gradient of tree residues, and assessed indicators that play critical roles in soil health such as: (i) soil aggregation, (ii) carbon and nitrogen storage, and (iii) biological activities through microbial and enzymatic assays.
2. Materials and Methods
2.1. Location and Management History
In 2017, we established a field trial in the district of Água Clara—Mato Grosso do Sul State (latitude 20°38′57.22″ S, longitude 52°35′50.74″ W, 370 m asl), within a commercial Eucalyptus urograndis hybrid (E. urophylla × E. grandis) plantation (Figure 1). According to the Köppen classification, the regional climate was tropical with a dry winter and a rainy summer (Aw), with an annual mean rainfall of around 1370 mm at an average of 24.4 °C [24].
Figure 1.
Location of the perimeter experimental area.
The soil of the experimental area was a sandy Arenosol [25], with a loamy sand textural class (sand contents of 843–878 g kg−1), originally covered by Cerrado sensu stricto (savanna-like woodland) [26]. At the time of evaluation, the trees were 1.7 years old.
2.2. Treatments and Experimental Design
The first cycle of 6-year-old Eucalyptus in the experimental area was cut in June 2017. The trees were harvested in the cut-to-length system, and at the end of September 2017 (120 days after harvest), the experimental plots were prepared, simulating the RMSs described below: cut-to-length (CTL)–maintenance of all residues (leaves, branches, bark, and litter) at the felling site, C:N ratio = 31; tree-length (TL)—maintenance of all residues except for the bark (leaves, branches, and litter), C:N ratio = 22; and bare litter (BL)—removal of all residues, i.e., the area was cleaned by sweeping, and only received plant biomass from the litterfall of the current cycle.
The experiment was conducted in a completely randomized design, with four replicates in the 0.00–0.10 m layer. Each management systems plot (40.8 × 27.6 m, i.e., 1126 m2) had 12 tree rows, and were spaced 3.4 m between rows and 2.3 m between plants. Within each plot, we considered the eight central rows (500.8 m2) for evaluations, thereby avoiding the border effects and interference from the neighboring areas.
Evaluations occurred 21 months after planting, in August 2019. The soil was sampled and analyzed to determine the effects of different eucalypt RMSs, their influence on the contents of nitrogen and organic carbon, and the microbiological and enzymatic activities of the soil.
2.3. Sampling and Aggregates from Dry Sieve Preparation
Undisturbed soil samples were collected in August 2019, in each plot. Four intact soil cores (0.10 m height × 0.10 m length × 0.10 m width) were taken at 3 m intervals from the center of each plot, and 0.00–0.10 m from the surface layer.
To minimize aggregate disruption, each core was manually removed and sieved through a 4.00 mm sieve crumbling along the natural fracture lines. According to [27,28], the fractionation procedure was performed with four composite samples in each treatment, consisting of three subsamples from the 0.00–0.10 m layer, and each composite sample was air-dried and then stored at room temperature.
All subsamples were shaken through sieves for 2 min, resulting in large macroaggregates that were retained from the 2.00 mm sieve, medium macroaggregates that were retained from the 0.250 mm sieve, and microaggregates that passed through the 0.250 mm sieve. The soil aggregates were then spread out in a thin layer and set to be air-dried. In summary, the subsamples were separated into large macroaggregates (LM: 4.00–2.00 mm), medium macroaggregates (SM: 2.00–0.25 mm), and microaggregates (MI: 0.250–0.00 mm), respectively.
2.4. Sand Correction of Aggregates
The sand correction procedure consisted of a combined 2 g of aggregate sample (LM, SM, and MI) with 2 mL of sodium hydroxide and 20 mL of deionized water, followed by shaking for 16 h on a horizontal shaker. After that, samples were passed through the 250 and 53 μm sieves for the macroaggregate sample, and only the 53 μm sieve for the micro-aggregate sample. All obtained fractions were oven dried at 60 °C and thus, the sand associated aggregate mass was subtracted from the initial aggregate amount, as indicated by Equation (1), resulting in sand free soil aggregate:
where aggregate fraction mass represents the mass of the fraction obtained in physical fractionation, and sand associated aggregate mass represents the sand weight in the aggregate sample.
Sand free soil aggregate (g) = aggregate fraction mass − sand associated aggregate mass
2.5. Soil Aggregates Respiration Measurement and Microbial Biomass Carbon (MBC)
The air-dried aggregates (20 g dry weight) of each size class were placed in a thin and loose layer at the bottom of 100 mL jars, with three replicates. The moisture content was adjusted to 40% of the water holding capacity, and the samples were incubated in the dark at 25 °C for 24 h. Small vials with 5 mL of 1 M NaOH were placed in incubation jars to trap CO2 after adding distilled water to the soil. In addition, three incubation jars containing only NaOH were used as blanks to correct for the CO2 trapped inside the jars. The basal respiration of soil aggregates (BSR—μg CO2-C g−1 dry soil day−1) was estimated by titrating each jar with NaOH and 2 mL 1 M BaCl2 (1:1) with 0.1 M HCl and phenolphthalein indicator (1% w/v in ethanol).
The soil microbial biomass carbon (MBC—μg C g−1 dry soil) of each aggregate size was determined using the chloroform-fumigation-extraction method described by [29]. After fumigation, the microbial carbon was extracted with 50 mL of 0.5 M K2SO4 from all the samples. The amount of MBC was determined by assessing the difference between the carbon extracted from the fumigation and non-fumigation controls using a coefficient (Kc) of 0.33 [29].
The metabolic quotient (qCO2) was obtained by the ratio between the values of released CO2-C and MBC (μg1 CO2-C soil day−1 of μg−1 MBC), according to [30]. The microbial quotient (qMic) was obtained by the ratio between the MBC and the soil total organic carbon (MBC/TOC) × 100 and was expressed as a percentage [31].
2.6. Enzyme Activity
Beta-Glucosidase (β-Glu) activity was determined according to [32], with the exception that toluene was omitted from our assays. Analyzes were conducted in triplicate with a non-substrate control. The method conducted was based on a colorimetric determination of the p-nitrophenol released by this soil enzyme when 1 g of soil sample is incubated with buffered substrate solutions. A total of 1 mL of 0.05 M p-nitrophenyl-a-D-Glucosidase (PNG) buffered solution was used as the substrate. The p-nitrophenol released was extracted by filtration and was measured using a microplate reader (BioTek Instruments, Winooski, VT, USA) set at a wavelength of 410 nm.
The fluorescein diacetate hydrolytic activity (FDA) was determined using the method described by [33]. The FDA hydrolysis assay can provide an estimate of the overall microbial activity in the soil sample by measuring it in a wide variety of enzymes, including esterases, proteases, and lipases [34]. We used 1.0 g of soil aggregates, air-dried, with 20 mL of 60 mM sodium phosphate buffer (pH 7.6), and mixed for 15 min at 100 rpm. Then, we added 100 µL of 4.9 mM FDA substrate solution (20 mg 3′6′-diacetyl-fluorescein in 10 mL acetone), and mixed for 1 h and 45 min at 100 rpm placed at 37 °C. After incubation, acetone (20 mL) was added and mixed into the suspension, which was centrifuged, and the supernatant was filtered through a Whatman No. 4 paper filter. Standard curves were generated using serial dilutions of a standard solution containing 0.5 g fluorescein sodium salt (Sigma F6377; C2H10O5Na2) in 500 mL distilled water. Samples and standards were determined with absorbance readings using a microplate reader (BioTek Instruments, Winooski, VT, USA) fixed at a wavelength of 490 nm.
2.7. Total Carbon and Nitrogen
For each soil aggregate fraction sample in each RMS, approximately 2 g of dry soil was finely ground and subsampled for total carbon (TC) and total nitrogen (TN) determined by combustion on an elemental analyzer (Costech Analytical Technologies, Inc., Valencia, CA, USA).
2.8. Active Carbon
Active C assessed through the amount of oxidized soil organic matter (SOM) has also been used to estimate the size of the labile C pool. Analysis was adapted from the method of [35], and cited by [36]. Briefly, 20 mL of 0.02 mol L–1 KMnO4 was added to 50 mL screw-top centrifuge tubes containing 2.5 g air-dried soil. The tubes were shaken for 2 min at 240 oscillations min−1 and allowed to settle for 10 min. Following this, 0.5 mL of the supernatant was transferred into a second 50 mL centrifuge tube and mixed with 49.5 mL of deionized water. The sample absorbance was read with a spectrophotometer at 550 nm and active C (mg kg−1 soil) was calculated as Equation (2):
where 0.02 mol L−1 is the initial concentration of the KMnO4 solution, a is the intercept of the standard curve, b is the slope of the standard curve, Abs is the absorbance of the sample, 9000 mg is the amount of C oxidized by 1 mol of MnO4, with Mn7+ getting reduced to Mn4+, and 0.02 L is the volume of KMnO4 solution that reacted with the soil. The samples consisted of soil aggregates and are expressed in kg.
2.9. Statistical Analyzes
The results were analyzed with the analysis of variance (ANOVA), followed by the Tukey test at p < 0.05, for means comparison. The relationship between the aggregate diameter classes (AgDi) and the harvest residue management systems (RMSs) was computed using the mean and standard error (SE) in R software [37].
3. Results
The statistical analysis revealed that most variables (MBC; qCO2; qMic; TN; TOC; FDA; β-Glu; and active carbon) showed significant F-values for the impact of the residue management systems (RMSs), except for BSR. Moreover, significant F-values were observed for all variables concerning the effect of the aggregate diameter (AgDi) and the interaction between the RMS and AgDi (RMS × AgDi). The soil consisted of over 78% ± 8% in the largest macroaggregate class (LM), 3 ± 0.6% of SM, and 19 ± 8% of MI, respectively (Figure 2). The wide variation of each aggregate diameter significantly influenced the response of the evaluated properties (p < 0.001) (Table 1). Mean weight diameter estimated based on the proportion of each aggregate size particle [38] was 1795 μm in TL, 1600 μm in CTL, and 1531 μm in BL, respectively.
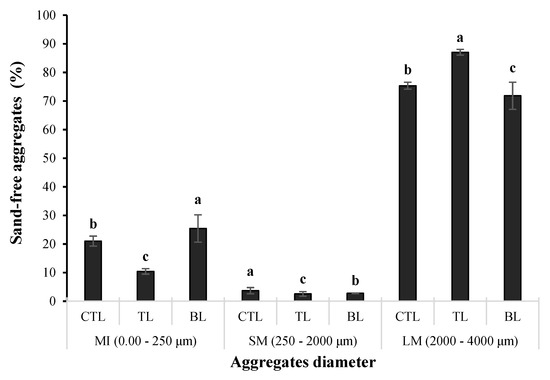
Figure 2.
The proportion of each soil aggregate class within the whole soil, classified by the aggregate diameter for three Eucalyptus urograndis harvest residue management systems. Error bars represent the standard errors (n = 4). Bars followed by different lowercase letters within each aggregate diameter were found to be significantly different by the Tukey test, at p ≤ 0.05. Abbreviations: LM, large macroaggregates (4.00–2.00 mm); SM, medium macroaggregates (2.00–0.25 mm); MI, microaggregates (0.250–0.00 mm), CTL, cut-to-length (maintenance of all harvest residues); TL, tree-length (removal of the bark and retention of the other residues); and BL, bare litter (removal of all harvest residues and receiving only the plant biomass from the current cycle’s litterfall).

Table 1.
Partitioning of the interaction between the Eucalyptus harvest residues management systems (RMSs) and the sandy free aggregate diameter (AgDi) for soil properties (whole soil).
To better understand the use of energy resources by the microbial biomass, we compared the basal soil respiration (BSR), which uses CO2 as a proxy for respiration, to the microbial biomass carbon (MBC), which denotes the size of the microbial community present, and defined the qCO2 as BSR/MBC. A significant interaction was found between the RMS and AgDi for the aggregate classes MI and SM, respectively. Under TL, the qCO2 in MI exceeded that than under BL and CTL three and four times, respectively, and was the highest qCO2 observed, with no variations in LM between the RMSs. Higher qCO2 values indicate a lower efficiency of the microorganisms in the use of organic material for energy [39], which was observed in the MI class under TL (Table 1), while the highest efficiency of microbial performance was observed in the SM class under BL and TL.
Digging deeper into the controlling factors of qCO2, it was apparent that basal soil respiration (BSR) (Table 1) did not vary significantly across the evaluated RMSs (Table 1). However, AgDi and RMS × AgDi interactions revealed that while MI released on average 4.27 ± 0.92 μg CO2 per gram of whole soil daily, LM respired four times as much, and SM only one-quarter of this amount (Table 1). For the MI class, BSR was the highest under BL, and LM respired most under TL, where 24% and 61% more CO2-C was released than in CTL and BL, respectively. SM BSR did not differ across the RMSs.
The MBC varied for each RMS, AgDi, and RMS × AgDi interaction (Table 1). While LM retained the most carbon density and SM the least under each RMS, MI MBC decreased from a median range in CTL and BL to indistinguishable levels from SM under TL with a five times lower mean. Under the two RMSs where vegetation remains on the surface (CTL and TL) to LM, the MBC values were approximately 20% higher than with the complete residue removal under BL.
To see where excess organic matter remains available to the microbial community, the MBC/TOC ratio defines the qMic. The MI qMic of the CTL as well as the BL, was found to significantly exceed that under TL by 27% on average. For the SM, BL had a significantly higher qMic than the RMS with residues, dwarfing CTL by 59% and 19% over TL, respectively. The lowest qMic was observed in LM (Table 1).
This directs insight towards nutrient levels, revealing that the contents of TOC and TN varied significantly different across the RMSs and AgDi, especially in the LM class. Twenty-one months after setting up the experiment, the harvest residues left on the surface significantly changed the TOC and TN contents only in the aggregates of the LM class, varying as TL > CTL > BL (Table 1). The TOC pattern differed only slightly from that of TN, with LM dominating MI by 90% and 88% respectively, and by 98% for both elements in SM, although TOC MI and SM were statistically indistinguishable (Table 1).
The total soil enzymatic activity (FDA), based on the measurement of fluorescein diacetate hydrolysis was not directly influenced by the different RMSs (Table 1); however, the RMS × AgDi interaction revealed a significant deviation of 15% less FDA under BL in the LM class, and generally CTL and TL presented the highest activities (31.43 and 33.98 mg fluorescein g−1 soil h−1, respectively), again grouping where the harvest residues remained on the soil.
The activity of the β-Glu enzyme responded to the RMSs in that the highest activity occurred in LM under CTL, where all eucalyptus harvest residues, including the bark, persisted on the surface (Table 1). The total removal of the harvest residues in BL halted β-Glu activity by 13% and 21% compared to TL and CTL, respectively, in this AgDi. The limited processing in the TL RMS lowered β-Glu activity by 10% compared to CTL.
The FDA and β-Glu activities under the different RMSs were lowest in the SM, except under TL where MI and SM did not differ for either enzyme (Table 1). Nevertheless, comparing the proportions of MI and LM in the soil, accounting for approximately 20% and 75% of the sand-free aggregates, respectively, clearly shows the relevance of MI.
Soil active carbon differed significantly for RMSs, AgDi, and their interactions (RMS × AgDi) across all aggregate diameters. In each RMS, the active carbon contents were highest in the LM and lowest in the SM, with MI at an intermediate content. Importantly, active C in the LM was 20 times higher than in the SM, which reinforces the need to maintain the soil structure, ensuring that carbon remains readily available. In MI, BL had a higher active carbon content than CTL (17%) and TL (60%) (Table 1). In SM, the active carbon contents followed CTL > BL > TL, with decreases of 24 and 30%, respectively. LM was highest under the TL RMS and decreased by 13% under CTL and 18% under BL, respectively. These results emphasize the differences in soil carbon storage. Since active carbon is readily available carbon, a higher content in LM may be associated with a greater amount of labile C (Table 1).
4. Discussion
4.1. Availability and C:N Ratio of Residues Affect C Cycle-Related Indicators
The management of harvest residues greatly influences carbon cycling in the soil. Large macroaggregates (LM) constitute 70–85% of sand-free soil, followed by microaggregates (MI) and small macroaggregates (SM), which make up 10–25% and 2–5% of the soil, respectively. The bare litter (BL) system, which removes all the harvest residues from the soil surface, exhibited the lowest proportion of LM, resulting in a decrease in the carbon content compared to the CTL and TL systems, which maintained some harvest residue. Aggregate-associated carbon is protected from decomposition, and in loamy and clayey soils, aggregates represent an average of 60% of total soil, and can store up to 54 kg per hectare of carbon [40]. In contrast, aggregates only represent 13% of the total soil in sandy soils, storing substantially less carbon at an average of 18 kg per hectare [41]. In this study, the share of C inside the LM reached 6.8 tons C ha−1 under BL (top 5 cm), representing a reduction of 22.3% and 37.5% compared to CTL and TL, respectively.
For sandy soils, this indicates that even small decreases in the proportion of aggregates or the concentration of carbon inside the aggregates can result in proportionally large losses of soil carbon. After harvest, tree residual biomass serves as the primary carbon source for soil microbes. Additionally, higher microbial and enzymatic activities were expected in LM for CTL and TL than BL due to the known improvements in soil structure resulting from the increased organic matter input on the soil. Indeed, biological activities in LM, such as microbial biomass carbon (MCB), active carbon, and enzyme activities (FDA and β-Glu), were found to have also slowed down under BL, as demonstrated by β-Glu activity in the decreasing order of CTL > TL > BL (36.62, 28.79, and 24.98 mg pNP g−1 soil h−1, respectively). Under similar climatic conditions, removing forest residues resulted in a 50% reduction in soil microbial respiration and a 40% reduction in MBC contents, respectively [3]. Our results also align with the findings of [21], who reported that aggregate size significantly affects microbiological activities in native pasture on sandy soil (loamy sand texture), where BSR was higher in LM and SM than in MI. Other studies have also reported increases in BSR and MBC with increasing aggregate size [22,23]. Based on these responses, the CTL and TL systems maintain soil health and functioning better than BL, as the residue input stimulates microbial-related processes, thereby promoting nutrient cycling and primary productivity, in addition to soil conservation provided by the surface coverage [42].
While removing all residues decreased total organic carbon (TOC) contents and slowed down biological activities, the presence of bark residue may have caused similar effects. TL (all residues minus bark) outperformed CTL (all residues including bark) for the proportion of LM (by 13.45%) and the LM’s concentration of TOC and total nitrogen (TN) by 19.6% and 16.7%, respectively. Similarly, most biological activities in the LM were enhanced in TL compared to CTL, namely basal soil respiration (BSR) (by 21%), MBC (by 5%), FDA (by 7.5%), and active C (by 13.5%). The slightly faster decomposition of the soil organic material (C:N ratio = 22) contributed to the increase in the activity of the microorganisms, which participate more actively in the soil aggregate process through fungus hyphae and bacterial filaments, as well as by the presence of binding substances [43,44]. However, in the CTL, a slightly slower decomposition (C:N ratio = 33) was observed due to the presence of the bark, meaning this effect was not so intense. Aggregate formation is normally associated with the stabilization of organic carbon inside aggregates, where it is protected from decomposition, and in this situation it was possible to observe increments in organic carbon and aggregate size [45]. The expected faster decomposition of residues under TL may have resulted in an enhanced release of binding agents, contributing to the larger proportion of LM under TL compared to CTL.
The stability of aggregate-associated C in sandy soils of tropical climates is unknown. Estimating the long-term stability of aggregate-associate C would require considering turnover rates of aggregates [46], and the environmental drivers of soil organic matter dynamics. Identifying sustainable soil management practices is therefore critical to addressing long-term C storage [20].
4.2. Stability and C Accumulation in Soil Aggregates
Across all resource management systems (RMS), it was found that the proportion of SM and MI was on average 9.6 and 1.8% lower than in LM, respectively. Moreover, TOC concentration in SM and MI was, on average, 50% lower than in LM. Considering the proportion of each aggregate size class and their respective TOC concentration, the amount of TOC associated with LM was 57 and 10 times greater than that associated with MI and SM, respectively. The microbial quotients (qMic) can serve as indicators to describe the relationship between the amount of microbial biomass carbon (MBC) and the soil TOC [39,47]. Our results indicate that MI and SM had qMic’s around 5.3 times larger than LM. In the LM fraction, there was no significant difference in the qMic observed between the treatments. High qMic values indicate that a large proportion of the TOC is actively cycled by the microbes, which can lead to increased CO2 emissions and decreased soil carbon sequestration, showing the vulnerability of these fractions to C losses [48].
This highlights the vulnerability of these fractions to carbon losses [41] and supports the idea that large aggregate size fractions can help protect and stabilize soil carbon. The present findings support the conclusions of [49], who conducted a mechanistic study demonstrating a direct correlation between the C input and total organic carbon (TOC) levels. Furthermore, they showed that C inputs from high input systems are more likely to be sequestered in larger aggregates, including both large and small macroaggregates.
When comparing different RMSs, TL had the lowest proportion of MI and the largest proportion of LM. This indicates that the MI may have been used in the formation of LM in this system [6]. On the other hand, in BL, where no residues were left on the soil, there may have been a breakdown of LM, as this system had the highest proportion of MI. These results suggest that the presence of these residues can help stabilize LM by promoting the formation of new aggregates, while the absence of residues may lead to the breakdown of existing LM [20]. Therefore, resource management practices that promote the formation and stabilization of LM, such as TL, should be encouraged to improve the soil structure and enhance soil health.
5. Conclusions
Identifying sustainable soil management practices is critical to addressing long-term issues of environmental degradation and global climate change. This study provides valuable insights into the stability of aggregates and carbon cycling in sandy soils under Eucalyptus plantations in tropical climates. Our research reveals that a significant proportion (about 54%) of these soils are made up of large macroaggregates (LM, 2000–4000 µm), which rely on harvest residues for their formation and stabilization. When trees were processed off-site and all harvest residues were removed, as in the bare litter treatment, we observed a significant decline in LM-associated total organic carbon (TOC), microbial biomass carbon, FDA, β-glucosidase activity, and active C after 21 months, compared to treatments where harvest residues were left on site. Removing these residues also led to an increase in the amount of microaggregates (MI, 0–25 µm) in the soil, indicating that some of the MI was not used to build LM, or that the absence of residue input contributed to the breakdown of LM. Our analysis of aggregate soil fractions revealed that LM was the most effective in protecting and stabilizing soil C, as evidenced by the lowest microbial quotient (qMic), which was about one order of magnitude lower than MI and small macroaggregates (SM, 25–2000 µm). These findings demonstrate that the preserving of these residues on site (i.e., CTL) offers a greatly improved option for sandy soils in the West-Center of Brazil. Our results also indicate that the CTL has a great potential to improve soil biological conditions and the efficacy of nutrient cycling from residues, as verified by the microbial enzyme soil. Therefore, maintaining the harvest residues on site is essential to promote the formation of LM and the stabilization of soil C in sandy soils under Eucalyptus plantations, and the adoption of CTL can significantly contribute to sustainable soil management practices.
Author Contributions
Conceptualization, T.S.B., K.L.M., A.M.R.C. and E.I.P.P.; Methodology, K.N.S.; Formal analysis, K.N.S.; Writing—original draft, K.N.S.; Writing—review & editing, T.S.B., K.L.M., A.M.R.C. and E.I.P.P.; Supervision, K.L.M. and E.I.P.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the São Paulo Research Foundation (FAPESP, grant number 2017/14049-2) and the Coordination of Superior Level Staff Improvement (CAPES) for providing doctorate scholarship and financial support to K.N.S. and T.S.B., respectively.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the Eldorado Brazil, for allowing evaluations and analysis in their experimental sites, and for the logistical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Gonçalves, J.L.; Alvares, C.A.; Higa, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; de Barros Ferraz, S.F.; de Paula Lima, W.; Brancalion, P.H.S.; Hubner, A. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Laclau, J.P.; Ranger, J.; de Moraes Gonçalves, J.L.; Maquère, V.; Krusche, A.V.; M’bou, A.T.; Nouvellon, Y.; Saint-André, L.; Bouillet, J.P.; de Cassia Piccolo, M. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; de Moraes Gonçalves, J.L.; Brandani, C.B.; de Vicente Ferraz, A.; Franci, A.F.; Marques, E.R.G.; Arthur Junior, J.C.; Hubner, A. Forest residue removal decreases soil quality and affects wood productivity even with high rates of fertilizer application. For. Ecol. Manag. 2018, 430, 188–195. [Google Scholar] [CrossRef]
- Ferreira, G.W.D.; Soares, E.M.B.; Oliveira, F.C.C.; Silva, I.R.; Dungait, J.A.J.; Souza, I.F.; Vergutz, L. Nutrient release from decomposing Eucalyptus harvest residues following simulated management practices in multiple sites in Brazil. For. Ecol. Manag. 2016, 370, 1–11. [Google Scholar] [CrossRef]
- Vinhal-Freitas, I.C.; Corrêa, G.F.; Wendling, B.; Bobuľská, L.; Ferreira, A.S. Soil textural class plays a major role in evaluating the effects of land use on soil quality indicators. Ecol. Indic. 2017, 74, 182–190. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Du, H.; Zeng, F.; Peng, W.; Wang, K.; Zhang, H.; Liu, L.; Song, T. Carbon Storage in a Eucalyptus Plantation Chronosequence in Southern China. Forests 2015, 6, 1763–1778. [Google Scholar] [CrossRef]
- Malinovski, J.R.; Camargo, C.M.S.; Malinovski, R.A.; Malinovski, R.A.; Castro, G.P. Sistemas. In Colheita Florestal, 3rd ed.; Machado, C.C., Ed.; UFV: Viçosa, Brasil, 2014; Volume 3, pp. 178–205. [Google Scholar]
- De Souza, I.F.; de Barros, N.F.; da Silva, I.R.; Renier, R.F.; de Ávila Silva, L.; de Novais, R.F. Decomposition of eucalypt harvest residues as affected by management practices, climate and soil properties across southeastern Brazil. For. Ecol. Manag. 2016, 374, 186–194. [Google Scholar] [CrossRef]
- Barros, I.B.; Cavalcante, V.S.; Moulin, A.S.; da Silva, I.R.; de Barros, N.F.; Vergütz, L.; Valadares, S.V. Integrating Forest residue and mineral fertilization: Effects on nutrient acquisition, nutrient use efficiency and growth of eucalypt plants. For. Ecol. Manag. 2021, 496, 119461. [Google Scholar] [CrossRef]
- Momolli, D.R.; Schumacher, M.V. O manejo dos resíduos florestais afeta a produção de biomassa e a sustentabilidade de povoamentos de Eucalyptus sp. Rev. Bras. Gestão Ambient. Sustentabilidade 2019, 6, 251–261. [Google Scholar] [CrossRef]
- Tisdall, J.; Oades, J. Organic-matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, X.; Zhu, A.; Yang, W.; Zhang, J.; Ding, S.; Mu, L.; Shao, L. Linking macroaggregation to soil microbial community and organic carbon accumulation under different tillage and residue managements. Soil Tillage Res. 2018, 178, 99–107. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Nicolodi, M.; Gianello, C. Understanding Soil as an Open System and Fertility as an Emergent Property of the Soil System. Sustain. Agric. Res. 2014, 4, 94. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Erickson, T.E.; Dixon, K.W.; Merritt, D.J. Soil quality indicators to assess functionality of restored soils in degraded semiarid ecosystems. Rest. Ecol. 2016, 24, S43–S52. [Google Scholar] [CrossRef]
- Insam, H.; Haselwandter, K. Metabolic quotient of the soil microflora in relation to plant succession. Oecologia 1989, 79, 174–178. [Google Scholar] [CrossRef]
- Araujo, A.S.F.; Cesarz, S.; Leite, L.F.C.; Borges, C.D.; Tsai, S.M.; Eisenhauer, N. Soil microbial properties and temporal stability in degraded and restored lands of Northeast Brazil. Soil Biol. Biochem. 2013, 66, 175–181. [Google Scholar] [CrossRef]
- Gupta, V.V.S.R.; Germida, J.J. Soil aggregation: Influence on microbial biomass and implications for biological processes. Soil Biol. Biochem. 2015, 80, 3–9. [Google Scholar] [CrossRef]
- Vicente, L.C.; Gama-Rodrigues, E.F.; Gama-Rodrigues, A.C.; Marciano, C.R. Organic carbon within soil aggregates under forestry systems and pasture in a southeast region of Brazil. Catena 2019, 182, 104139. [Google Scholar] [CrossRef]
- Yang, C.; Liu, N.; Zhang, Y. Soil aggregates regulate the impact of soil bacterial and fungal communities on soil respiration. Geoderma 2019, 337, 444–452. [Google Scholar] [CrossRef]
- Li, N.; Yao, S.H.; Qiao, Y.F.; Zou, W.X.; You, M.Y.; Han, X.Z.; Zhang, B. Separation of soil microbial community structure by aggregate size to a large extent under agricultural practices during early pedogenesis of a Mollisol. Appl. Soil Ecol. 2015, 88, 9–20. [Google Scholar] [CrossRef]
- Zhang, H.J.; Ding, W.X.; Yu, H.Y.; He, X.H. Linking organic carbon accumulation to microbial community dynamics in a sandy loam soil: Result of 20 years compost and inorganic fertilizers repeated application experiment. Biol. Fertil. Soils 2015, 51, 137–150. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; de Moraes Gonçalves, J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports, 106; FAO (Food and Agriculture Organization of the United Nations): Rome, Italy, 2015. [Google Scholar]
- Instituto Brasileiro de Geografia e Estatística (IBGE). Mapa de Biomas do Brasil; Primeira Aproximação. Escala 1:5 000.000. Projeção Policônica. Meridiano de Referência: 54 W. Gr; Paralelo de Referência: 0, 2014. Available online: https://geoftp.ibge.gov.br/informacoes_ambientais/estudos_ambientais/biomas/mapas/biomas_5000mil.pdf (accessed on 9 March 2023).
- Elliott, E.T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Tian, J.; Pausch, J.; Yu, G.R.; Blagodatskaya, E.; Gao, Y.; Kuzyakov, Y. Aggregate size and their disruption affect C-14-labeled glucose mineralization and priming effect. Appl. Soil Ecol. 2015, 90, 1–10. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kec value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Domsch, K.H. The metabolic quotient of CO2 (qCO2) as a specific activity parameter to assess the effects of environmental condition, such as pH, on the microbial of forest soil. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil Enzymes. In Methods of Soil Analysis: Part 2. Microbiological and Biochemical Properties, 1st ed.; Weaver, R.W., Angle, J.S., Bottomley, P.S., Eds.; Soil Science Society of America Book Series; Soil Science Society of America: Madison, WI, USA, 1994; Volume 3, pp. 775–833. [Google Scholar]
- Adam, G.; Duncan, H. Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 2001, 33, 943–951. [Google Scholar] [CrossRef]
- Benckiser, G.; Schnell, S. Biodiversity in Agricultural Production Systems; CRC Press—Taylor and Fancis Group: Boca Raton, FL, USA, 2006; p. 429. [Google Scholar]
- Weil, R.; Islam, K.R.; Stine, M.A.; Gruver, J.B.; Samson-Liebig, S.E. Estimating active carbon for soil quality assessment: A simplified method for laboratory and field use. AJAA 2003, 18, 3–17. [Google Scholar]
- Moebius-Clune, B.N.; Moebius-Clune, D.J.; Gugino, B.K.; Idowu, O.J.; Schindelbeck, R.R.; Ristow, A.J.; van Es, H.M.; Thies, J.E.; Shayler, H.A.; McBride, M.B.; et al. Comprehensive Assessment of Soil Health. In The Cornell Framework, 3rd ed.; Cornell University: Geneva, Switzerland, 2017; pp. 53–54. ISBN 0-967-6507-6-3. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 March 2021).
- Van Bavel, C.H.M. Mean weight-diameter of soil aggregates as a statistical index of aggregation. Soil Sci. Soc. Am. J. 1950, 14, 20–23. [Google Scholar] [CrossRef]
- Feketeová, Z.; Hrabovský, A.; Šimkovic, I. Microbial Features Indicating the Recovery of Soil Ecosystem Strongly Affected by Mining and Ore Processing. Int. J. Environ. Res. Public. Health 2021, 18, 3240. [Google Scholar] [CrossRef] [PubMed]
- Okolo, C.C.; Gebresamuel, G.; Zenebe, A.; Haile, M.; Eze, P.N. Accumulation of organic carbon in various soil aggregate sizes under different land use systems in a semi-arid environment. Agric. Ecosyst. Environ. 2020, 297, 106924. [Google Scholar] [CrossRef]
- Sena, K.N.; Maltoni, K.L.; Troleis, M.J.B.; Faria, G.A. Forest harvest management systems and residual phytomass affecting physical properties of a sandy soil. Rev. Bras. Cienc. Solo 2021, 45, e0200190. [Google Scholar] [CrossRef]
- Du, X.; Jian, J.; Du, C.; Stewart, R.D. Conservation management decreases surface runoff and soil erosion. Int. Soil Water Conserv. Res. 2022, 10, 188–196. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Miller, R.M.; Jastrow, J.D. Mycorrhizal Fungi Influence Soil Structure. In Arbuscular Mycorrhizas: Physiology and Function; Kapulnik, Y., Douds, D.D., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 3–18. [Google Scholar]
- Mohanty, M.; Sinha, N.K.; Sammi Reddy, K.; Chaudhary, R.S.; Subba Rao, A.; Dalal, R.C.; Menzies, N.W. How Important is the Quality of Organic Amendments in Relation to Mineral N Availability in Soils? Agric. Res. 2013, 2, 99–110. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, X.; Yang, W.; Zhu, A.; Ding, S. Short-term decomposition, turnover and retention of residue-derived carbon are influenced by the fertility level in a sandy loam soil. Geoderma 2019, 349, 68–78. [Google Scholar] [CrossRef]
- Boni, T.S.; Pereira, E.I.P.; Santos, A.A.; Cassiolato, A.M.R.; Maltoni, K.L. Biomass residues improve soil chemical and biological properties reestablishing native species in an exposed subsoil in Brazilian Cerrado. PLoS ONE 2022, 17, e0270215. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Kong, A.Y.Y.; Six, J.; Bryant, D.C.; Denison, R.F.; van Kessel, C. The relationship between carbon input, aggregation, and soil organic carbon stabilization in sustainable cropping systems. Soil Sci. Soc. Am. J. 2005, 69, 1078–1085. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).