Abstract
The aim of this study was to determine the effect of drought on the accumulation and tolerance of energy willow (Salix viminalis L. var. ‘Gigantea’) to copper (Cu) and nickel (Ni) in the context of phytoremediation potential of the plant and biomass production under adverse water conditions. Drought was simulated with polyethylene glycol (PEG-6000. 5%), and metals were added at a concentration of 1 mM. Plants were cultivated in greenhouse conditions for 21 days according to the experimental variants: control, Cu, Ni, PEG, PEG + Cu and PEG + Ni. The results indicate high toxicity of Cu (chlorosis, necrosis, decrease in biomass, plant dehydration, increase in the content of proline and phenolic compounds), and PEG + Cu co-treatment increased the toxicity of the metal. Ni applied at the same concentration did not cause toxicity symptoms. The willow exhibits the ability to accumulate Ni, and mutual application of PEG + Ni increased Ni uptake to new shoots. Cu caused elevated accumulation of proline and phenolics in leaves accompanied with a decreased carbon and nitrogen content in roots in favor of young shoots. Both metals and drought led to disruption in the content of mineral nutrients (Ca, Mg, Fe). Due to high tolerance to Ni and drought, S. viminalis var. ‘Gigantea’ bears high potential for biomass production on Ni-polluted sites with accompanying metal uptake increased under water deficit.
1. Introduction
The cultivation of energy crops for biomass is one of the solutions to the problem of the exhaustion of conventional energy sources. Low quality of coal and its insufficient supply, as well as the emission of pollutants associated with the combustion of fossil fuels are main environmental concerns [1]. For efficient production of biomass as a renewable energy source, it is necessary to select plants with high yield, even when cultivated under adverse soil and weather conditions [2].
One of the plants that can be cultivated on both agricultural and degraded lands is energy willow, a perennial plant, which is obtained as a result of crossing and selection of various species of willows. In the first year of cultivation, it grows up to 1.8 to 2.4 m tall, while in the third (most often during this period the crop is harvested) it reaches a height of 5.77 m on average [3]. For individual varieties, annual growth and resistance to environmental factors differ compared to the wild willow. Due to genetic variation and differences in soil requirements, the selection of the variety or taxon to the characteristics of the cultivation area is crucial [4]. Various genotypes of energy willow are cultivated for the purpose of production of briquettes, pellets or biomass for biogas plants. Energy willow biomass can also be used for co-combustion with coal [5] or to produce bioethanol [6,7].
Plants of the Salix genus are grown mainly as a source of biomass for energy purposes; however, they can be used to neutralize sewage sludge and slurry applied to fertilize plants [8]. Other benefits of willow cultivation range from soil protection against erosion, to the creation of buffer zones at water intakes (water protection), and to creating natural barriers against air pollution or noise. The willow plantation acts as a habitat for birds, insects and other animals, and willow wood is used for the production of furniture, boards, paper, charcoal and in wicker basketry [9,10]. Willows, as obligate phreatophytes, need a permanent water supply for efficient biomass production. Their tap roots go deep into the soil and reach the water table, while a capillary fringe forms above it. Thus, willows and poplars can acclimate to the environmental conditions of different geographical regions and succeed in both biomass production for energy and in phytoremediation and soil cleanup [11]. Recent studies revealed the potential of energy willow for phytoremediation of degraded lands in industrial areas such as landfills, mine tailings, flotation sludge deposits, industrial plants and communication routes where the soil is degraded [12]. During the remediation process, plants take up contaminants from the soil through roots and store them or metabolize them in their plant cells [13,14]. Energy willow accumulates heavy metals such as copper (Cu), iron (Fe), manganese (Mn) and zinc (Zn) (in roots), and cadmium (Cd) and Zn (in leaves) [15]. Besides metal accumulation, the cultivation of willow plants restores soil properties. In the recent study [14], after two years of growing energy willow in degraded soil, the content of humus and C/N ratio increased, and the sodium content decreased by 12.6%. However, willow genotypes vary in their tolerance to heavy metal presence in soil [16], and significant differences can be found in their physiological and biochemical response to metal stress, including the profile of low-molecular weight organic acids, phenolic compounds or non-protein thiols [11,17].
Drought is an additional abiotic stress threatening willow crop photosynthesis capacity, thus lowering biomass yield. Soil drought is caused by lack of water in the ground, atmospheric drought by low humidity and high temperature, and physiological drought occurs when the plant is unable to take up available water due to high salinity, poor aeration, low soil temperature or damaged root system. In the temperate climate zone, over recent decades, the amount of precipitation in winter has increased significantly, while a significant decrease in precipitation is observed in spring and summer [18]. Drought or water deficiency results in a decline of the water potential in leaves and the closure of stomata by the action of abscisic acid (ABA). During drought stress, the plant reduces the expression of genes responsible for photosynthesis, while inducing the expression of genes responsible for the synthesis of ABA, osmoprotectants, which are metabolites protecting against the formation of reactive oxygen species (including phenolic compounds), and signaling genes that increase sensitivity to abscisic acid [19]. Reducing the intensity of photosynthesis and generating reactive oxygen species affect the plant condition negatively, reducing yield and damaging tissues [20,21].
Soil pollution with copper (Cu) and nickel (Ni) are concerns in mining and processing sites of non-ferrous ores. In mine tailings, their concentration may reach over 370 and 260 mg kg−1 (for Cu and Ni, respectively) [22]. However, in agricultural soil also, the Cu level was found to reach 506 mg kg−1 as a result of the application of copper-based fungicides over years [23]. The Finnish legislation established standard values as an approximation of national systems in Europe and India at 200 and 150 mg kg−1 (for Cu and Ni, respectively) for industrial and transport areas [24]. Above these values a remediation process should be employed.
Thus, the aim of this study was to determine the effect of simulated drought conditions on the accumulation of Cu and Ni by energy willow and their influence on the plant growth and response parameters under conditions of their interaction. The metals were chosen for this study based on our previous investigations on energy willow (S. purpurea × viminalis) in the context of its application for phytoremediation of copper–nickel mining sites.
2. Materials and Methods
2.1. Plant Material and Experiment Set-Up
The investigated willow variety was Salix viminalis var. ‘Gigantea’, a Danish one growing up to 4 to 10 m tall, demonstrating strong branching with thick trunks. In field cultivation, the variety gives large and stable yields on various types of soil, including those with high salinity [25,26,27].
The experiment was carried out in greenhouse conditions in April/May of 2022. Commercially obtained two-year cuttings (25 cm long and 10–15 mm in diameter) were rooted in a half-strength Knop’s medium for 14 days, and then transferred to hydroponic pots equipped with a net insert and a water level indicator (3 plants per pot, 2 pots per variant). Cultivation was carried out in a modified Knop’s medium and plant rods were stabilized with a steamed quartz sand (3–4 mm). In individual experimental variants, Cu(NO3)2 × 3H2O or Ni(NO3)2 × 6H2O were applied at an equimolar concentration of 1 mM, and polyethylene glycol (PEG-6000) was added to the medium to simulate drought stress at a moderate level (5%, w/v) [28]. Metal concentration was chosen based on our previous research on willows and both metals caused significantly different effects in the hydroponic system at 1 mM treatment. Higher Cu concentrations (up to 3 mM) caused severe growth retardation to S. purpurea × viminalis and plant death in prolonged treatment [17]. PEG is a water-soluble polymer, an inert and non-toxic chemical for plants. It was found to mimic drought stress and its action is based on changing the water potential of the cultivation solutions by inducing potential osmotic pressure [29]. Plants cultivated in a Knop’s solution were used as a control. The experiment comprised 6 variants: control (c), copper addition (Cu), nickel addition (Ni), drought simulation (PEG) and metal and drought combined treatments (PEG + Cu, PEG + Ni). Plants were irrigated daily with tap water according to their needs to maintain a constant water level according to the indicator. After 21 days of cultivation, plant material was collected for analyses, and individual organs (shoots and roots) were weighed. The tolerance index (TI) of willow plants was calculated for particular treatments (t) as a ratio of combined shoots and roots fresh weight vs. control (c) according to the modified formula [30]:
For biochemical investigations, fully developed middle leaves were harvested and frozen in liquid nitrogen in situ and stored at −80 °C for analyses. For the elemental investigations, the remaining tissue was dried at 105 ± 5 °C in an electric oven (SLW 53 STD, Pol-Eko, Wodzisław Śląski, Poland), and then ground in a Cutting Boll Mill PM 200 (Retsch GmbH, Haan, Germany). To meet the minimum weight requirements, fresh tissue from two plants (one from each pot separately) was combined.
2.2. Relative Water Content of Leaves
The middle leaves were weighed immediately after the harvest to determine fresh weight (FW) using an analytical balance. Afterwards, leaves were placed in plastic string bags with petiole soaked in distilled water and left for 4 h in the dark to reach full turgidity. After measuring the turgid weight (TW), leaves were dried in an electric oven at 80 °C for 48 h to determine dry weight (DW). The relative water content (RWC) of leaves was calculated using the following formula [31]:
2.3. Metal Content and Uptake Ratios
Atomic absorption spectrophotometry (AAS) was employed to study the total content of Ni and Cu, bivalent mineral nutrients (Ca, Mg) and Fe in willow roots and shoots. Subsamples of dried material (0.1–0.5 g ±0.0001 g) were weighed from a homogeneous sample and mineralized with concentrated HNO3 (Sigma Aldrich, Steinheim, Germany) in a Mars Xpress microwave mineralizer (CEM International Corporations, Matthews, NC, USA) for 30 min at an electromagnetic frequency of 2450 MHz. Mineralized samples were quantitatively transferred to volumetric flasks, diluted with demineralized water up to 25 or 50 mL and filtered through a paper filter. For each element, an appropriate calibration curve was prepared and the necessary conversion of sample weight and solution volume were applied. The metal concentration in the solutions was determined with an AA280 FS/Z AA spectrometer (Agilent Technologies, Santa Clara, CA, USA) using flame atomization. The calibration curve for each element was prepared based on a series of freshly prepared standard solutions obtained from 1000 mg L−1 basic standards (Sigma Aldrich, Steinheim, Germany). The final calibration curves for each element were prepared automatically using the spectrophotometer. The concentration range of the calibration curves was as follows: Ni: 1.0–6.0 mg L−1, Cu: 1.0–4.0 mg L−1, Ca, Mg and Fe: 1.667–5.000 mg L−1. The results were expressed as average values from three measurements. The total content of the element was expressed in mg kg−1 DW.
For investigated metals, the translocation factor (TF) was calculated as a ratio of metal content in shoots to roots. Bioconcentration factor (BCF) was calculated as a ratio of metal content in roots to its initial concentration in a cultivation medium [32,33].
2.4. Elemental Analysis of Plant Material
The content of nitrogen (N), carbon (C), hydrogen (H) and sulfur (S) in willow roots and shoots was determined using the FLASH 2000 Series elemental analyzer (Thermo Fisher Scientific, Waltham, MA, USA). The samples were weighed (2–3 mg) using an electronic microbalance (Mettler XP6, Mettler-Toledo, LLC, Columbus, OH, USA) into tin capsules (Thermo Fisher Scientific, Waltham, MA, USA) and introduced through the MAS 200R autosampler into the combustion reactor. After sample combustion, the resulting gases were detected by a thermal conductivity detector (TCD). The calibration curve was prepared using BBOT (2,5-bis(5-tert-butyl-benzoxazol-2-yl)thiophene standard) (Thermo Fisher Scientific, Waltham, MA, USA) and Alfalfa certified reference material (Elemental Microanalysis Ltd., Okehampton, UK). A five-point calibration curve was prepared for each element. The correctness of the calibration curve preparation was verified by the analysis of a certified reference material—Birch leaf (Elemental Microanalysis Ltd., Okehampton, UK).
2.5. Proline and Total Phenolic Content in Leaves
Willow leaves were homogenized in a liquid nitrogen using a mortar and a pestle. A sample of homogeneous material was extracted with 80% methanol, shaken in an orbital shaker, and then centrifuged (10,000 rpm, 15 min) to obtain a clear supernatant that was used for colorimetric assays.
For the analysis of proline, the colorimetric method was applied [34] using a Carry Bio (Varian) spectrophotometer. Standard solutions with a concentration range of 0.04 to 1 mM were prepared by dissolving the proline standard in an ethanol:water mixture (40:60, v/v). A reaction mixture (1000 µL) consisting of ethanol (20%, v/v), ninhydrin (1%, w/v) and acetic acid (60%, v/v) was mixed with the methanolic extracts (500 µL). The samples were placed in a water bath at 95 °C for 20 min. Afterwards, the samples were centrifuged (10,000 rpm, 10 min), and the resulting supernatant was transferred to a measuring cuvette. During incubation with an ethanolic solution of ninhydrin, a reaction takes place with proline to create a yellow product determined by measuring the absorbance at λ = 520 nm. The results of proline content were expressed in µmoles g−1 FW.
The total phenolic content (TPC) was determined according to the Folin–Ciocalteu assay with some modifications [35]. The extracts (200 µL) were mixed with 1 mL of a Folin–Ciocalteu reagent, and after 3 min 3 mL of 10% Na2CO3 was added. After 30 min of incubation in the dark at room temperature, the absorbance at λ = 765 nm was measured with a Cary 300 Bio UV–vis scanning spectrophotometer (Varian, Mulgrave VIC, Australia). Gallic acid (GA) was used as a calibration standard and the results were expressed in mg of GA equivalents g−1 FW. The extracts were diluted according to the concentration range of the standard curve (0.01–0.5 mg mL−1).
2.6. Statistical Analysis
Statistical analyses were performed using R software (ver. 4.2; R Development Core Team). The differences in growth and biochemical parameters as well as element concentration between treatments were assessed using one-dimensional analysis of variance (ANOVA). Analyses were followed by the multiple comparisons Tukey’s HSD or Dunnett test (which does not meet the criteria required for Tukey’s) as a post hoc analysis for significant effects. Test assumptions were checked with Levene’s test assessed homogeneity of variance. For Principal Component Analysis (PCA), data normalization was performed to prevent one variable from dominating others, and variables were shifted to be zero-centered. The data were visualized using ggplot2 package [36].
3. Results
3.1. Plant Growth, Toxicity Symptoms and Relative Water Content
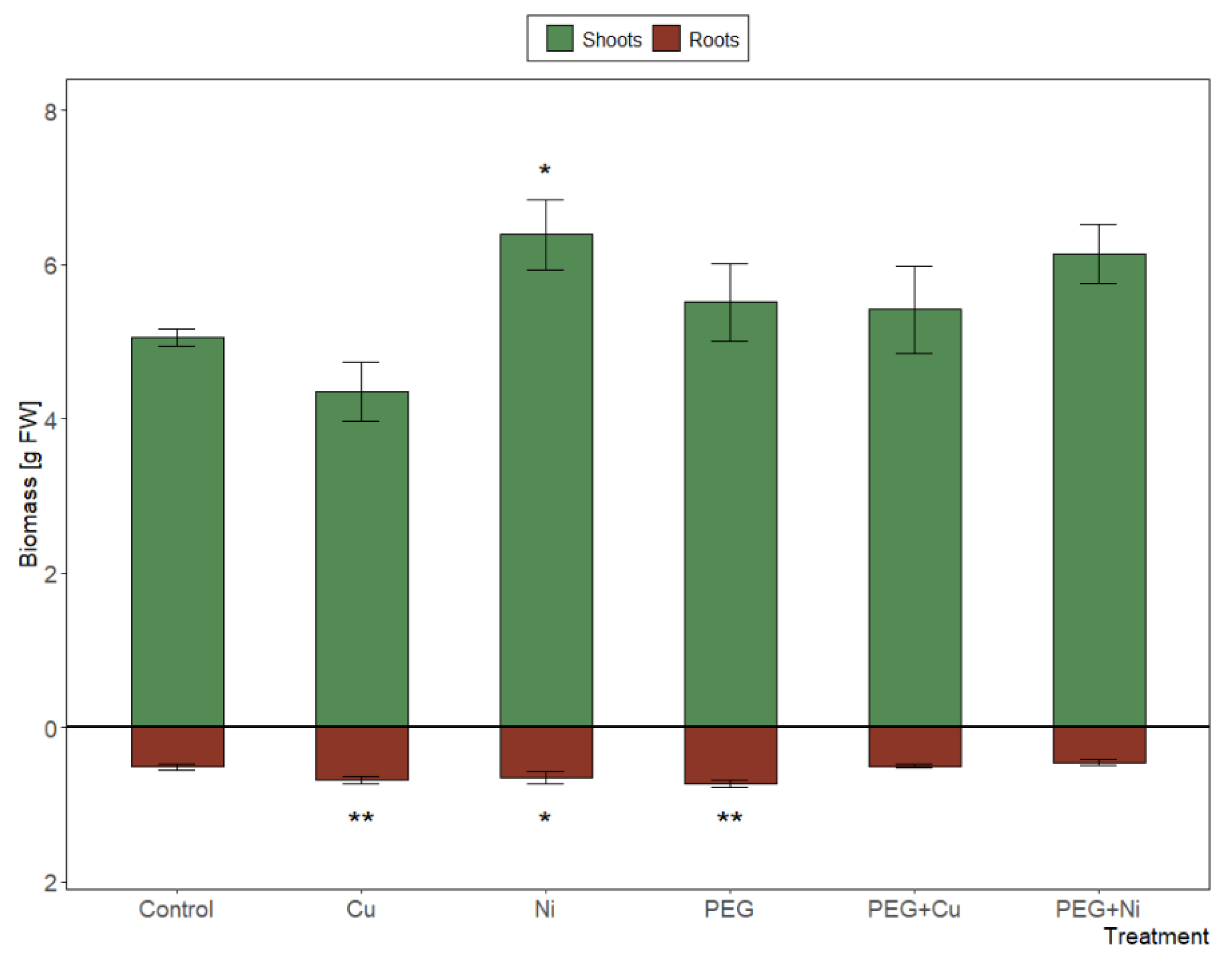
The significant (p < 0.001) effect of the applied treatments on the biomass of both roots and shoots was observed, along with a strong differentiation between Cu and Ni additions (5.03 and 7.04 g FW, respectively; p < 0.001) (Figure 1). Shoot biomass increased significantly in the presence of Ni (6.39 g FW; p < 0.05). Furthermore, the additions of Cu, Ni or PEG significantly induced root growth relative to the control, while the combined metal and PEG caused inhibition of root growth versus metal additions and drought simulation (p < 0.01). Consequently, the tolerance index calculated for roots and new shoots combined fresh weight showed the lowest value in the case of Cu addition (0.904), and the highest for Ni (1.264) vs. control plants. Combined metal and PEG additions did not cause significant changes relative to untreated control or treatment with PEG.
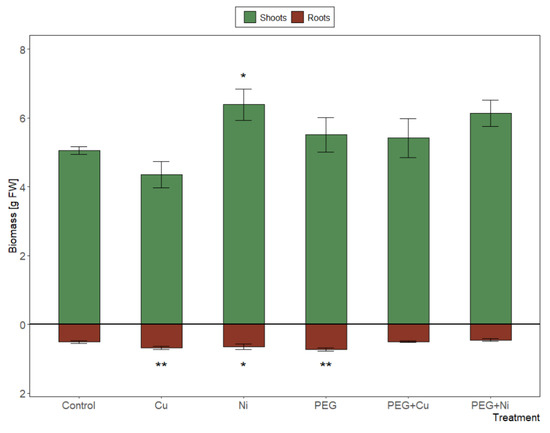
Figure 1.
Biomass of shoots and roots of S. viminalis var. ‘Gigantea’ in experimental variants (data presented as mean ± SD, n = 6; * p < 0.05, ** p < 0.01).
Morphometric changes and visible toxicity symptoms of leaves were also observed in response to applied treatments (Figure 2). The addition of Cu to the medium caused inhibition of the leaf growth and chlorosis. The addition of Ni caused a relatively weaker negative effect on the length and width of leaves, and did not cause significant symptoms on the leaf surface. Drought simulation causes chlorosis and necrotic lesions on the leaf surface without influencing shoot and leaf morphometry. Simultaneous drought and Cu treatments enhanced the negative effect of the metal and caused the reduction of leaf width and development of necrotic lesions (necrosis of the edges of the leaf blade).

Figure 2.
Representative leaves of S. viminalis var. ‘Gigantea’ in experimental variants.
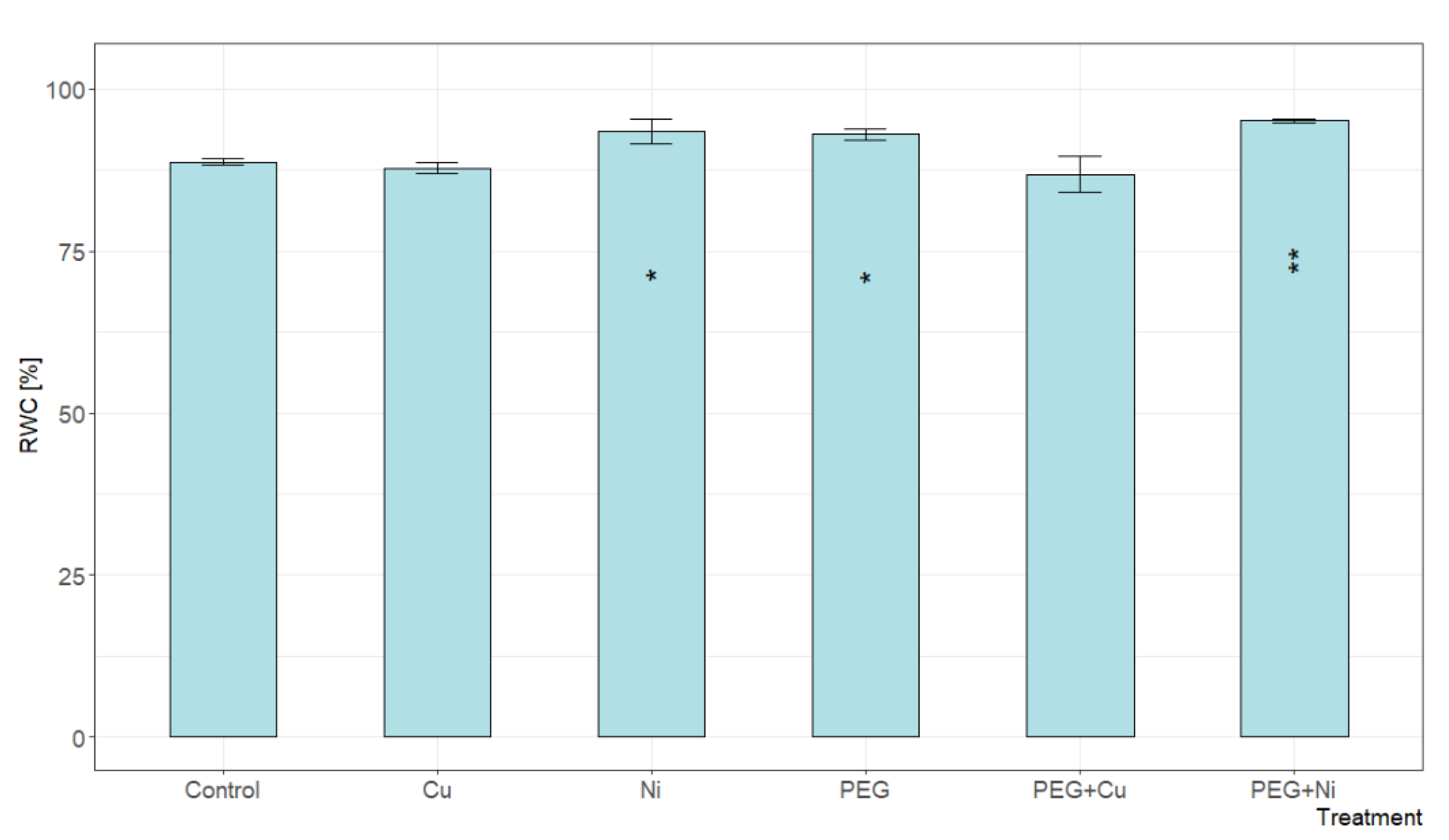
The relative water content (RWC) expressed as a percentage determines the degree of hydration of tissues, with 100% being full turgor. The addition of Cu and combined PEG + Cu treatments did not cause significant changes in the RWC level of willow leaves compared to the control (Figure 3). In contrast to Cu, the addition of Ni and PEG resulted in increased leaf tissue hydration. The highest RWC value was recorded for simultaneous PEG + Ni treatment, which was 7% higher than the control.
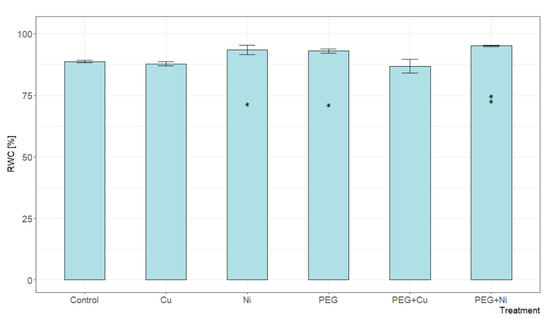
Figure 3.
The relative water content (RWC) of S. viminalis var. ‘Gigantea’ leaves in experimental variants (data presented as mean ± SD, n = 6; * p < 0.05, ** p < 0.01).
3.2. Metal Uptake and Mineral Nutrient Allocation
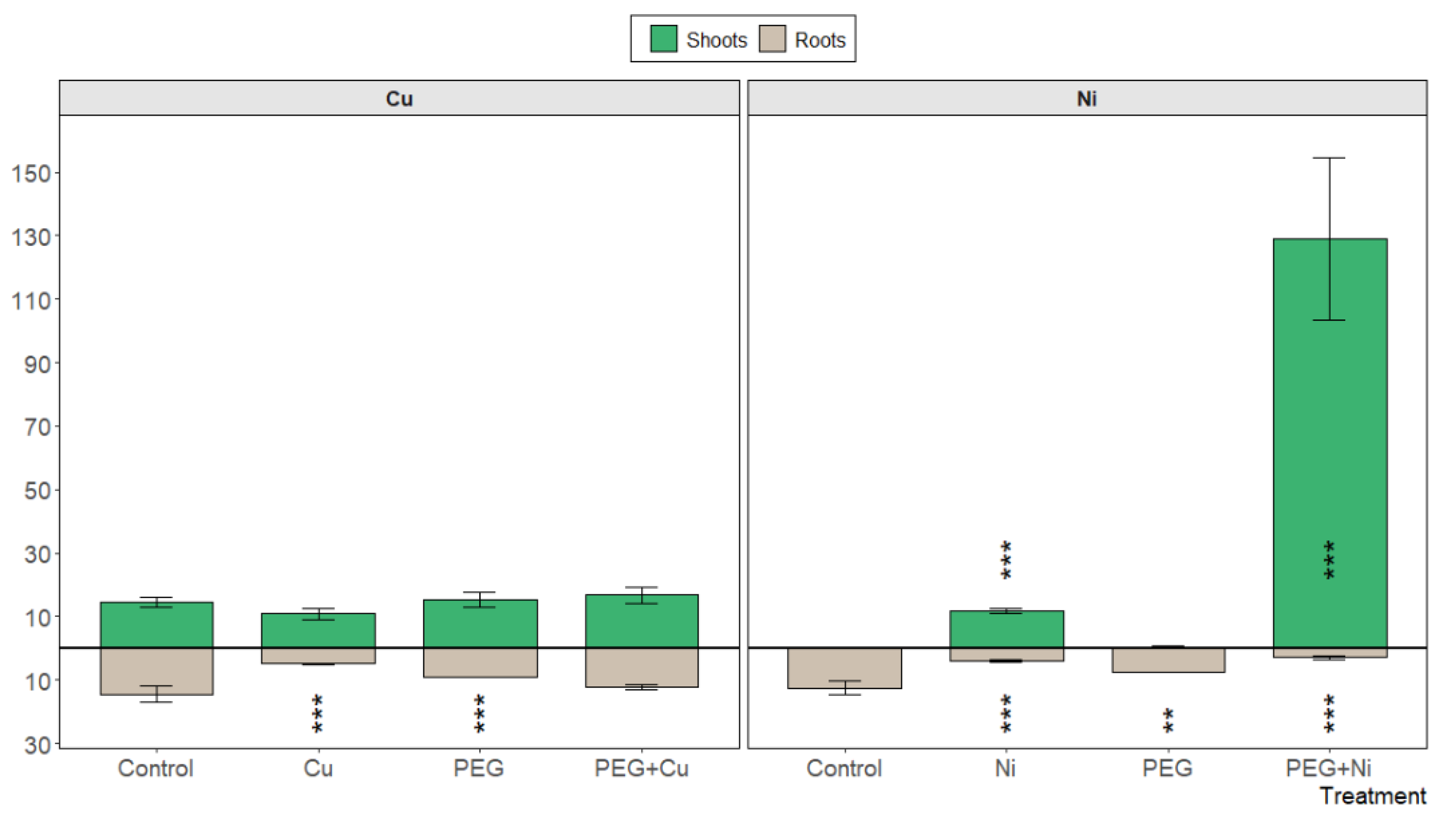
S. viminalis var. ‘Gigantea’ showed no accumulation capacity for Cu, applied also with PEG, in roots (Figure 4). Similarly, there was no increased transport of the metal to young shoots. In contrast, the addition of Ni to the cultivation medium changed the accumulation pattern in favor of willow shoots, while the overall metal load was only slightly increased compared to the control plants. However, nickel addition accompanied by simulated drought conditions stimulated nickel transport and accumulation in the newly developed photosynthetic tissue, leading to a conspicuous increase in Ni accumulation in young shoots (Figure 4).
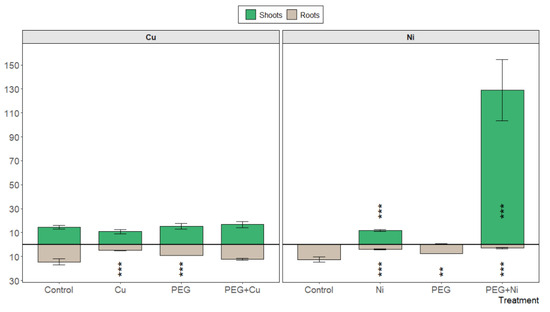
Figure 4.
The content of Cu and Ni [mg kg−1 DW] in shoots and roots of S. viminalis var. ‘Gigantea’ in experimental variants (data presented as mean ± SD, n = 3; ** p < 0.01, *** p < 0.001).
The bioconcentration factor did not reveal active accumulation of added metals in willow roots, which was confirmed by low absolute values of metal content. Similarly, low translocation factor values (<1) were calculated for Cu and Ni additions and for PEG + Cu combined treatment. The exception was found for Ni and PEG co-treatment with a TF value of 3.31, confirming the high translocation rate of Ni from roots to young shoots under drought simulation.
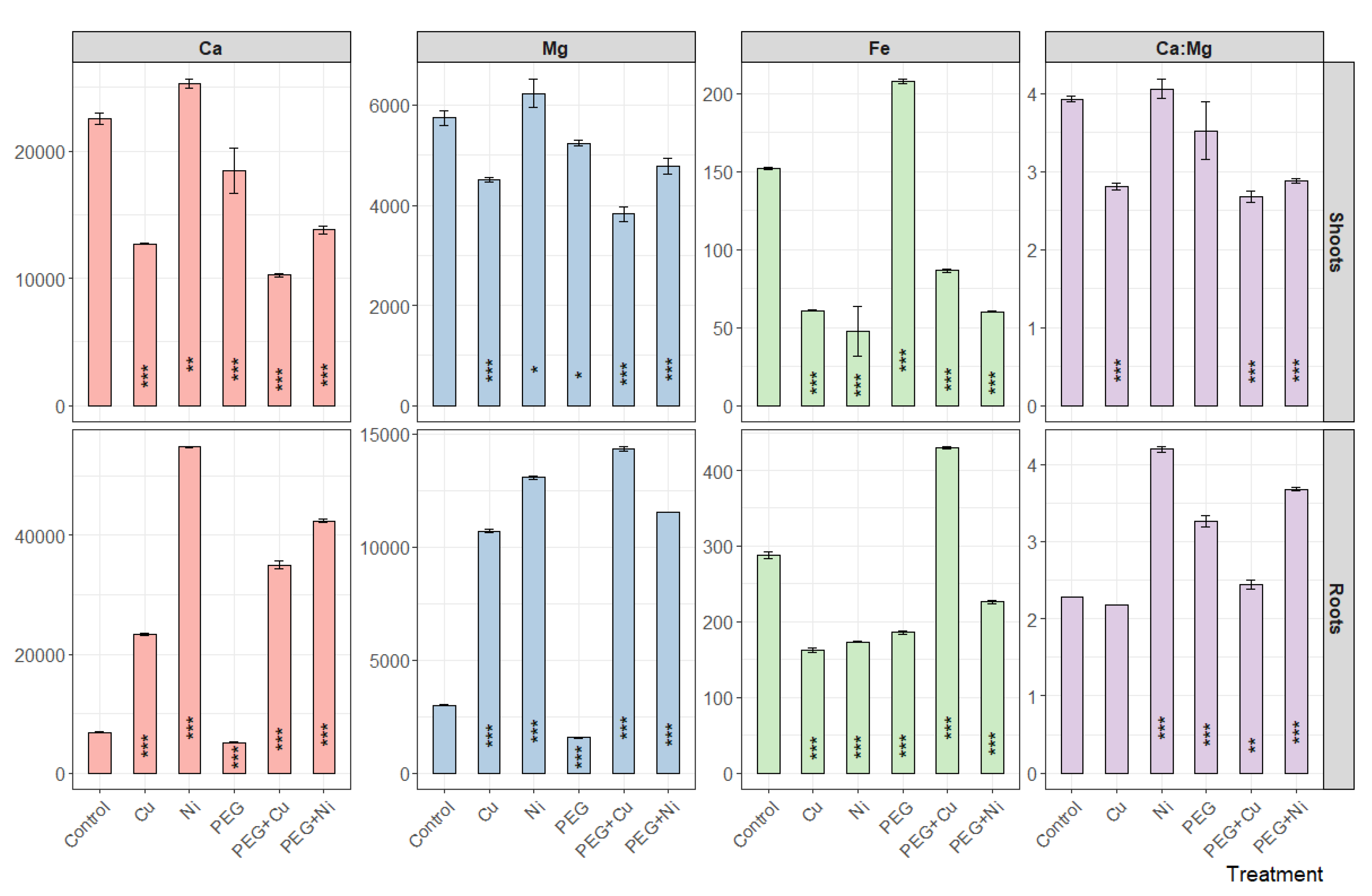
The applied treatments caused an overall decrease in Ca content in willow shoots, which was most pronounced in the case of Cu and PEG + Cu applications (Figure 5). The exception was the addition of Ni; however, metal application along with PEG strongly depleted the effect. In roots, an opposite response was observed, and metals (as isolated treatments as well as combined with PEG) caused a significant rise in Ca content, which was more pronounced in the case of Ni. A similar pattern was noted for Mg content and influenced the Ca:Mg ratio both in shoots and roots in a metal-specific manner (Figure 5). Consequently, in shoots a strong depletion was noted in the case of Cu addition and combined metals and PEG, while in roots a conspicuous increase was noted for Ni and PEG + Ni treatments. The addition of metals was accompanied with depletion of iron content in willow organs excluding PEG + Cu treatment in the case of roots (Figure 5). PEG elevated Fe content in shoots with a simultaneous reduction in roots to a similar level compared to metals.
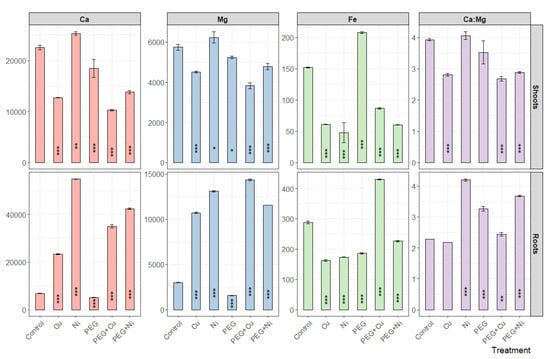
Figure 5.
The content of Ca, Mg, Fe [mg kg−1 DW] and Ca:Mg ratio in shoots and roots of S. viminalis var. ‘Gigantea’ in experimental variants (data presented as mean ± SD, n = 3; * p < 0.05, ** p < 0.01, *** p < 0.001).
3.3. Elemental Composition of Roots and Shoots
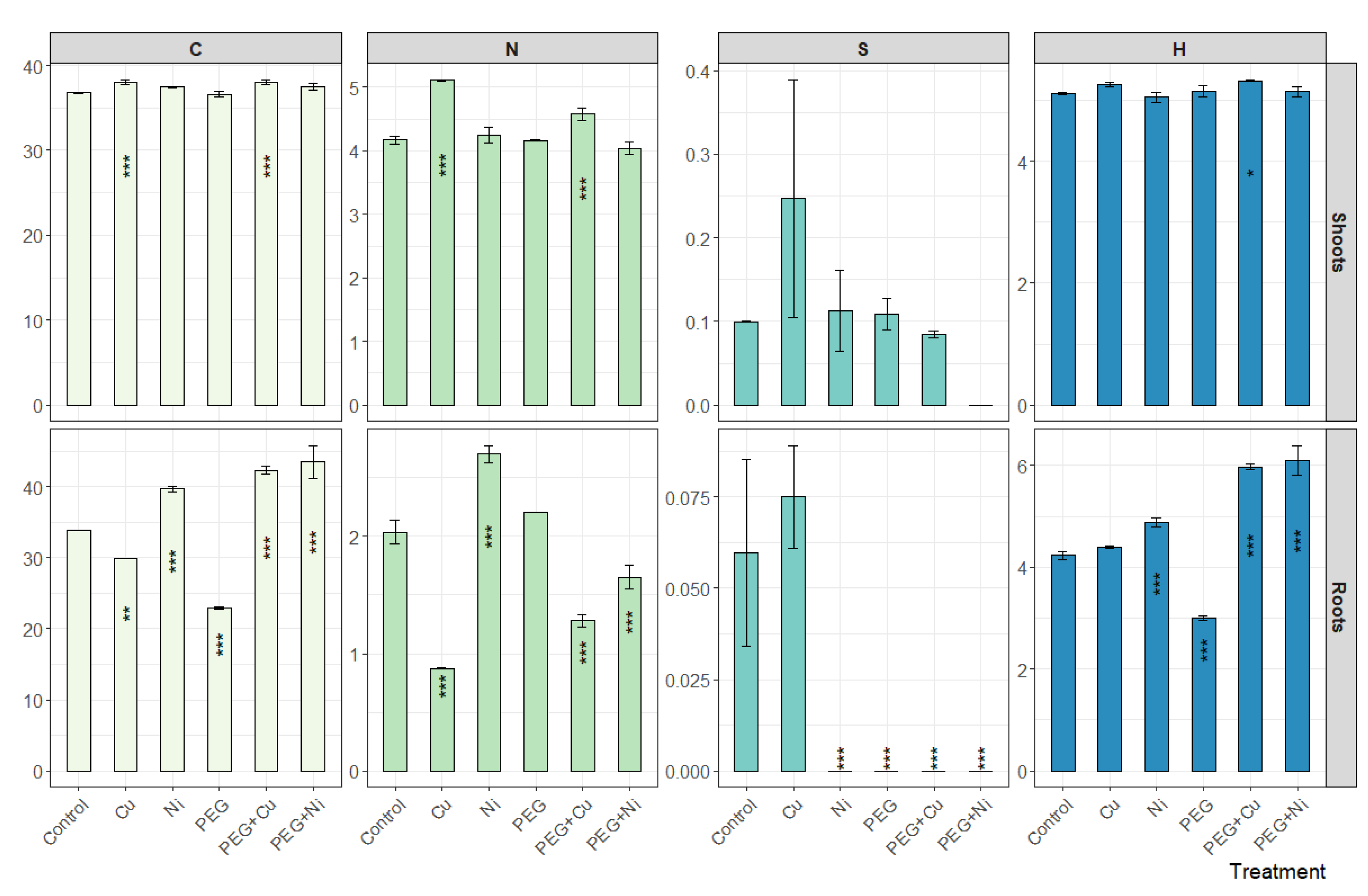
The analysis of elemental composition revealed a similar carbon content in willow shoots in experimental variants, amounting to 37.4% on average, and 36.8% for control. Among treatments, only Cu and PEG + Cu caused a slight but significant increase in C content in shoots. In the case of roots, the mean C content was similar to that in shoots; however, a greater variation was observed between treatments (Figure 6). The addition of Ni caused an increase, and Cu a decrease, in the total C content in roots compared to the control, while for drought simulation the most conspicuous decrease was noted. The highest C content in roots was observed for plants subjected to the metal accompanied with drought simulation, which was similar in the case of both applied metals, and were higher than in control plants by 8.4% (PEG + Cu) and 9.6% (PEG + Ni).
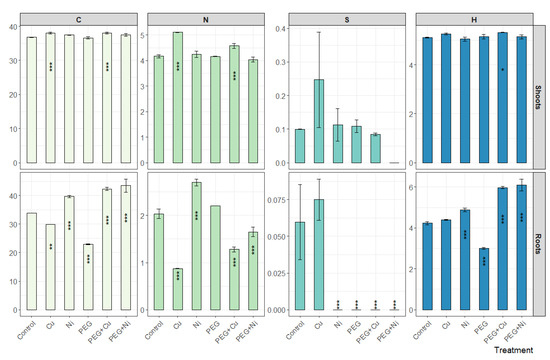
Figure 6.
Elemental composition [%] of shoots and roots of S. viminalis var. ‘Gigantea’ in experimental variants (data presented as mean ± SD, n = 3; * p < 0.05, ** p < 0.01, *** p < 0.001).
The mean content of nitrogen showed a higher level in shoots (4.38%) than in roots (1.79%) of the willow. In shoots, an increase in N content compared to the control was observed under the addition of Cu to the cultivation medium, also when combined with PEG, by 0.95 and 0.41%, respectively (Figure 6). The N content in roots also decreased under the influence of Cu and PEG + Cu additions, i.e., by 1.16 and 0.76%, respectively, compared to the control. The addition of Ni elevated the N content in roots in relation to the control (by 0.66%), while the limited availability of water reduced the induction effect of Ni on the N content leading to a decrease of 0.39% relative to the control.
The mean content of sulfur amounted to 0.11 and 0.02% in shoots and roots, respectively. The highest content of this element was recorded under the influence of Cu, while in other variants, the element was not present at a measurable level or the determined content was close to the control (Figure 6).
The hydrogen content amounted to 5.2 and 4.8% on average in shoots and roots, respectively. The addition of metals and PEG did not significantly affect the total H content in the willow shoots, and only PEG + Cu combined treatments slightly increased its content (Figure 6). In the case of roots, a slight increase of H content was observed following the addition of Ni and more intense in the case of metal and PEG combined treatments. At the same time, PEG addition caused a decrease in the total H content in roots.
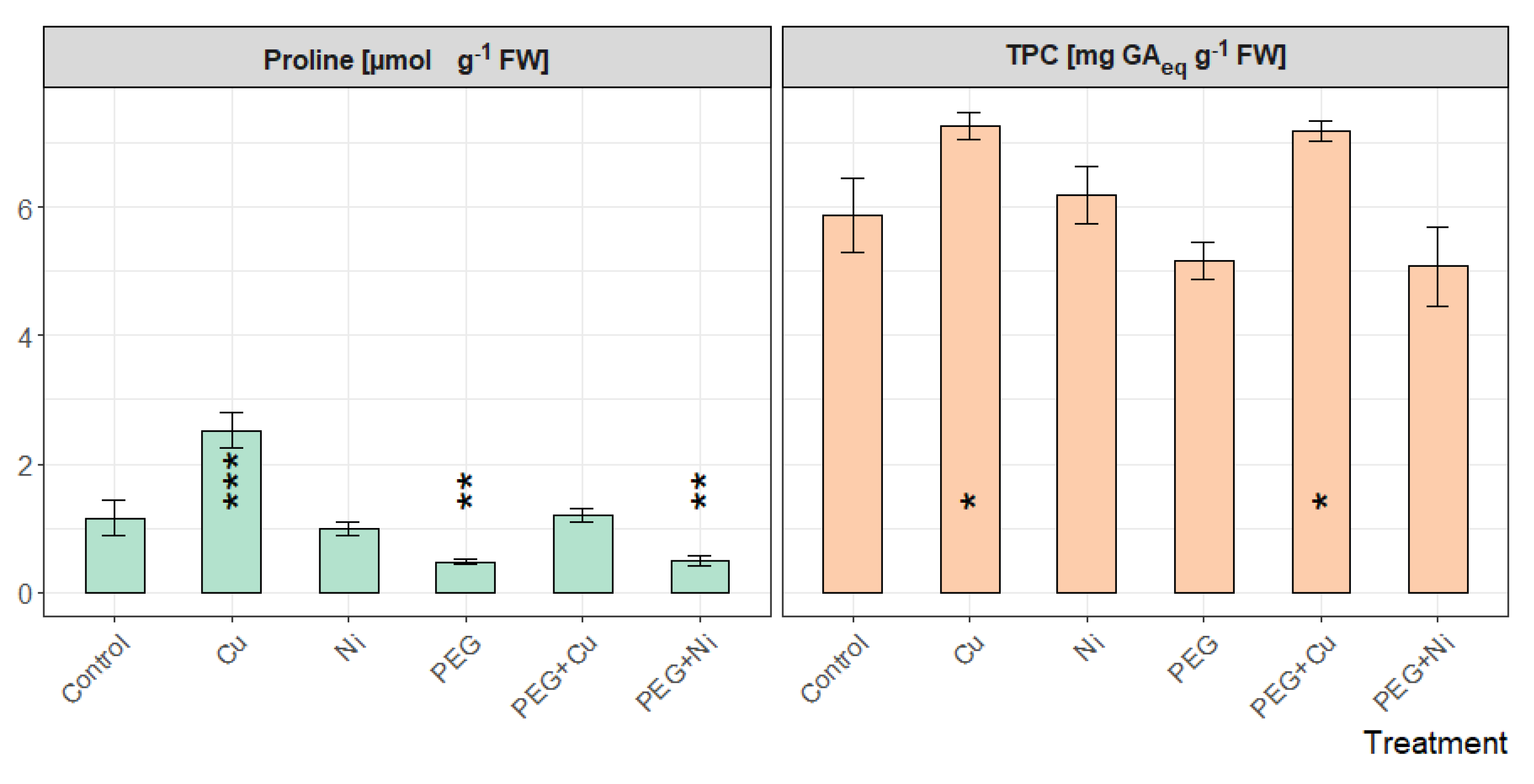
3.4. Proline and Total Phenolic Contents in Leaves
The highest content of proline in willow leaves was observed for plants cultivated with the addition of Cu and was significantly higher than that of control (Figure 7). PEG limited the accumulation of the amino acid caused by the presence of Cu in the medium (p < 0.001). The lowest proline content was observed in plants growing in medium with the addition of PEG, also combined with Ni, while (as in the case of Cu) the limited availability of water led to a decrease in proline content relative to the addition of Ni.
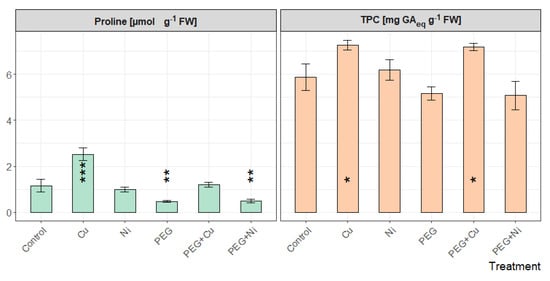
Figure 7.
Proline and total phenolic content (TPC) in leaves of S. viminalis var. ‘Gigantea’ in experimental variants (data presented as mean ± SD, n = 3; * p < 0.05, ** p < 0.01, *** p < 0.001).
Changes in phenolics accumulation shared a similar pattern of relative values as proline (Figure 7). The TPC in leaves showed increased values compared to the control for plants growing with the addition of Cu and PEG + Cu. In the case of remaining treatments, the TPC value was similar to the control.
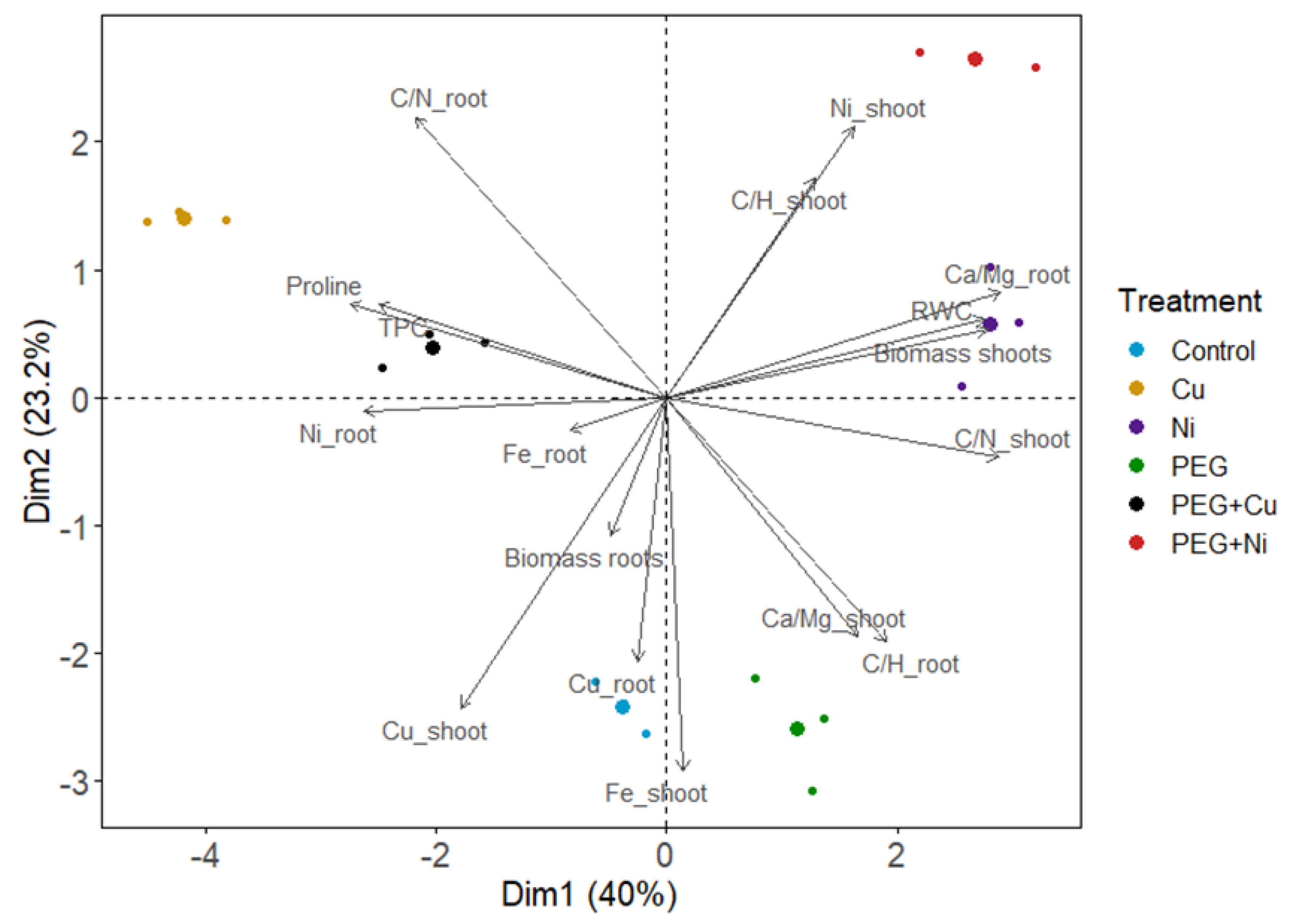
3.5. Principal Components Analysis (PCA)
The PCA analysis was performed for the investigated parameters, and the first two principal components explained 63.2% of the variability (Figure 8). The applied treatments were shown to differentiate studied parameters; however, major differences were found between metals (Cu and Ni), whereas drought simulation influenced plants to much lesser extent relative to the control. PEG + metal caused changes similar to the given metal, proving its dominant effect on investigated parameters. Considering correlations between variables, shoot biomass was strongly correlated with leaf RWC values and negatively related with proline and phenolics accumulation in willow leaves (Figure 8). Biomass of shoots was also correlated with Ni uptake, indicating Ni-induced stimulation, and accompanied by the increased C/N and C/H ratios. In contrast, the Cu level in shoots was negatively correlated with shoot biomass and C/N or C/H relative contents. Increased metabolite biosynthesis (proline and phenolics) was accompanied by a reduced C/N ratio and a shift in mineral nutrient balance manifested by the elevated Ca/Mg relative contents. Furthermore, Ni accumulation in shoots was negatively correlated with Fe content in this plant organ. Disruption in the content of mineral nutrient (Ca, Mg, Fe) in roots was also related with Cu and Ni uptake. Moreover, a decrease in the Ca/Mg ratio accompanied the induction of root growth.
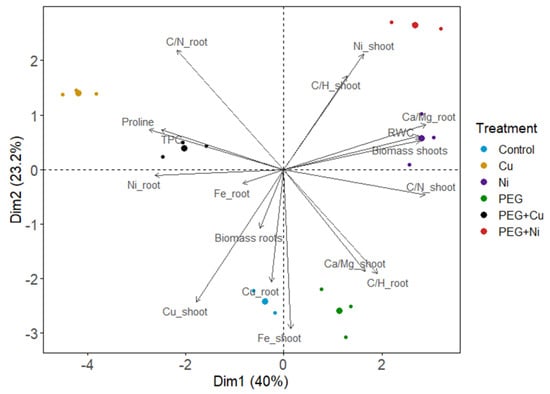
Figure 8.
Principal component analysis (PCA) of parameters of shoots and roots of S. viminalis var. ‘Gigantea’ in experimental variants (RWC—relative water content, TPC—total phenolic content).
4. Discussion
Biomass obtained from various plants is an environmentally friendly alternative to conventional energy sources [37,38]. Biomass cultivation, as well as other alternative sources of electricity and heat, is an increasingly important element of the energy market [38,39]. Recent studies seek tolerant plants capable of remediating polluted sites, such as mine tailing bearing high concentrations of heavy metals in soil [40]. Ideal plant species for phytoremediation should be high-biomass-producing, easily adaptive, tolerant to toxic contaminants, easy to cultivate and have high metal accumulation capacity [41]. Besides plant selection, various soil modifiers have been tested to improve restoration potential of particular plants [42]. The main features of willow meet the requirements for phytoremediation purposes [43]. However, a relatively high water demand may limit its application under drought conditions, especially when subjected to mutual metal and water stresses.
In this study, energy willow S. viminalis var. ‘Gigantea’ was subjected to elevated concentrations of metals (Cu or Ni) to evaluate tolerance and metal uptake. Additionally, plants were co-treated with PEG-6000 to simulate drought conditions and to assess the effect of limited availability of water on plant growth and tolerance to the applied metals. This study is a continuation of previous research on Salix showing diverse tolerance and response mechanisms to Cu and Ni under equimolar treatments and modified by mutual interaction between metals applied as a mixture [17]. The present study describes the effect of limited water availability as an additional stressor. The results are novel and shed light on the growing problem of drought potentially affecting willow plantations on degraded lands and modifying growth and phytoremediation efficiency.
The obtained results revealed that no increase in Cu uptake from the cultivation medium was observed, regardless of the drought simulation. In contrast, Ni was more strongly accumulated by willow plants than Cu and underwent strong translocation to photosynthetic organs, which confirms previous studies [44]. Drought conditions simulated by the addition of PEG strongly induced Ni transport and its accumulation in young shoots.
Copper is a necessary cofactor for various enzymes in plants, including cytochrome, phenolic, ascorbate oxidases, tyrosine iodinase, sulfide oxidase or laccase [45,46]. However, excess Cu can reduce the activity of these enzymes due to the supersaturation of the cofactor binding site. Although being a constituent of photosystem II (PSII), the excess Cu can also interfere with pigment and lipid biosynthesis affecting chloroplast ultrastructure and negatively influencing the efficiency of photosynthesis [47]. In the present study, despite the lack of significant Cu accumulation, toxicity symptoms were observed, in particular under drought conditions. Other studies also revealed high sensitivity of five other willow genotypes treated with a significantly lower Cu concentration (25 µM) in a hydroponic system [16]. In addition to direct toxicity by inhibition of the activity of key enzymes, Cu may also cause indirect effects by affecting the uptake of mineral nutrients from cultivation media/substrates, in particular Ca, Mg and Fe [45,48]. Significant and metal-specific changes in the Ca/Mg ratio were observed, i.e., in roots the ratio was increased by Ni, PEG and co-treatments, while in shoots a depletion accompanied Cu and PEG + metal additions. Disturbance of mineral homeostasis in leaves led to the inhibition of leaf growth and the occurrence of chlorosis. Further, Fe uptake was limited in plants treated with Cu, where causing chlorosis was the most apparent Fe-deficiency symptom in plants and significantly affected plant growth [46,49]. Fe is essential for crucial processes such as DNA synthesis, energy production (respiration), energy conversion (photosynthesis) and nitrogen reduction [50]. Due to its transition nature, Fe competes with other metals such as Cu, Zn and Mn in its uptake, transport and functioning within cells [50], which was confirmed in this study. Drought is also a cause of ionomic shift and its mode is linked to the severity of the applied stress. In Arabidopsis [51], nutrient uptake changed across the different water regimes tested, with underaccumulation (Fe, Ca, Mg) and overaccumulation (Na) with increasing water deficit. Similarly, a moderate water stress decreased nutrient uptake in of Solanum lycopersicum [52], Brassica napus and Triticum aestivum [53]. As assumed, drought affects genes primarily involved in ion transport and homeostasis and secondarily in metal chelation or responses to metals. The identification of specific mechanisms remains difficult, although disturbances in the hormonal balance and oxidative status of the plant may be key factors [53].
In contrast to copper, nickel is not an essential element for the proper functioning of plants, and in excess may damage the assimilation apparatus and cause chlorosis. However, in this study a stimulation of biomass production under the influence of Ni was observed despite increased accumulation and translocation of this element within the plant. Similar differences in Cu and Ni toxicity were demonstrated in our previous studies [17]. Cu strongly induced sucrose accumulation in leaves rather than Ni. In contrast, Ni caused elevated accumulation of salicylic acid, which correlated with glutathione (GSH) accumulation proving its induction effect on the GSH biosynthesis [17].
Metal application caused disturbances in the elemental composition of root and shoot tissue. In the case of Cu, an increase in carbon and nitrogen content was noted, indicating increased biosynthesis of nitrogen-containing molecules. This corresponded with a simultaneous increase in the content of the amino acid proline induced by the presence of Cu in the medium. A similar increase was observed for sulfur, which may indicate the accumulation of thiol compounds with antioxidant and chelating properties (GSH, phytochelatins (PCs)) under the influence of Cu. At toxic concentrations, Cu may induce PC accumulation and form stable complexes with PCs [54].
Moreover, drought may also increase carbon allocation belowground and shift root traits towards greater resource acquisition including soluble sugar and starch storage [55]. The observed allocation of C and N to roots could also result from the enhanced biosynthesis of primary metabolites (sugars, amino acids and organic acids) serving as root exudates secreted into the rhizosphere to enhance plant–microbiome interactions under stress stimuli and the availability of water, phosphorus and other nutrients [56].
Proline protects the plant against water loss in drought conditions [57,58]. The analysis of the content of proline in plants is, next to the determination of the relative water content, a method confirming the occurrence of drought stress [59]. The highest concentration of proline was observed in Cu-treated plants, while the lowest was in the PEG + Ni variant. For these variants, the highest and lowest percentage of nitrogen content was observed, which confirms an association between stress level and induction of amino acid and protein biosynthesis. The applied drought simulation by the addition of PEG reduced the content of proline relative to the control and the addition of Cu and Ni. This may indicate a relatively high tolerance of the willow cultivar to the applied PEG concentration, and proves regulatory function of proline towards antioxidant enzymes induced by Cu besides its osmoprotective properties [60]. Further, an induction of plant response to Cu-caused oxidative stress was manifested by increased accumulation of phenolic metabolites serving chelating and antioxidant functions [61,62]. The addition of Cu caused an increase in the concentration of proline correlated with the total content of phenolic compounds in willow leaves, while the lowest concentrations of these metabolites were present in plants grown in medium with the addition of PEG.
5. Conclusions
Copper at a concentration of 1 mM causes toxicity to energy willow S. viminalis var. ‘Gigantea’ in a hydroponic culture. The addition of nickel at the same level stimulates plant growth. Drought simulation by the addition of polyethylene glycol (PEG-6000) at a concentration of 5% did not cause significant effects in the tested willow cultivar, including a decrease in biomass yield and the content of protective metabolites. The simultaneous addition of Cu and PEG aggravates the toxicity of the metal. The accumulation of proline and phenolic compounds in leaf tissue was a defense reaction of willow plants to the presence of Cu. Enhanced biosynthesis of bioactive metabolites was associated with a decrease in the percentage of carbon and nitrogen in the roots in favor of young shoots. The willow strongly accumulates Ni in the shoots, and the simultaneous addition of PEG reduces the accumulation of metal in the roots and strongly increases its transport to young shoots. Due to high tolerance to Ni and drought, S. viminalis var. ‘Gigantea’ shows high potential for biomass production on Ni-polluted sites with accompanying metal accumulation properties increased under limited water availability. This study provides a basis for further studies on toxicity thresholds and uptake abilities of the willow in soil systems under differentiated water regimes.
Author Contributions
Conceptualization, K.D.; methodology, K.D., M.W. and M.R.; software, M.R.; validation, K.D., M.W. and M.R.; formal analysis, K.D. and M.R.; investigation, K.D., P.G. and M.W.; resources, K.D.; data curation, M.R.; writing—original draft preparation, K.D., M.W., P.G. and M.R.; writing—review and editing, K.D.; visualization, M.R.; supervision, K.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rentier, G.; Lelieveldt, H.; Kramer, G.J. Varieties of coal-fired power phase-out across Europe. Energy Policy 2019, 132, 620–632. [Google Scholar] [CrossRef]
- Akerboom, S.; Botzen, W.; Buijze, A.; Michels, A.; van Rijswick, M. Meeting goals of sustainability policy: CO2 emission reduction, cost-effectiveness and societal acceptance. An analysis of the proposal to phase-out coal in the Netherlands. Energy Policy 2020, 138, 111210. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Yield, energy parameters and chemical composition of short-rotation willow biomass. Ind. Crops Prod. 2013, 46, 60–65. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Krzyżaniak, M. Cost of heat energy generation from willow biomass. Renew. Energy 2013, 59, 100–104. [Google Scholar] [CrossRef]
- Sahu, S.G.; Chakraborty, N.; Sarkar, P. Coal–biomass co-combustion: An overview. Renew. Sustain. Energy Rev. 2014, 39, 575–586. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Łuczyński, M.; Załuski, D.; Szczukowski, S.; Tworkowski, J.; Gołaszewski, J. Lignocellulosic biomass from short rotation woody crops as a feedstock for second-generation bioethanol production. Ind. Crops Prod. 2015, 75, 66–75. [Google Scholar] [CrossRef]
- Su, T.; Zhao, D.; Khodadadi, M.; Len, C. Lignocellulosic biomass for bioethanol: Recent advances, technology trends, and barriers to industrial development. Curr. Opin. Green Sustain. Chem. 2020, 24, 56–60. [Google Scholar] [CrossRef]
- Urbaniak, M.; Wyrwicka, A.; Tołoczko, W.; Serwecińska, L.; Zieliński, M. The effect of sewage sludge application on soil properties and willow (Salix sp.) cultivation. Sci. Total Environ. 2017, 586, 66–75. [Google Scholar] [CrossRef]
- Schulz, U.; Brauner, O.; Gruß, H. Animal diversity on short-rotation coppices—A review. Landbauforsch. Volkenrode 2009, 59, 171–181. [Google Scholar]
- Volk, T.A.; Verwijst, T.; Tharakan, P.J.; Abrahamson, L.P.; White, E.H. Growing fuel: A sustainability assessment of willow biomass crops. Front. Ecol. Environ. 2004, 2, 411–418. [Google Scholar] [CrossRef]
- Marmiroli, M.; Pietrini, F.; Maestri, E.; Zacchini, M.; Marmiroli, N.; Massacci, A. Growth, physiological and molecular traits in Salicaceae trees investigated for phytoremediation of heavy metals and organics. Tree Physiol. 2011, 31, 1319–1334. [Google Scholar] [CrossRef]
- Prabha, J.; Kumar, M.; Tripathi, R. Opportunities and challenges of utilizing energy crops in phytoremediation of environmental pollutants: A review. Bioremediat. Environ. Sustain. 2021, 383–396. [Google Scholar] [CrossRef]
- Dimitriou, I.; Aronsson, P. Wastewater and sewage sludge application to willows and poplars grown in lysimeters—Plant response and treatment efficiency. Biomass Bioenergy 2011, 35, 161–170. [Google Scholar] [CrossRef]
- Scriba, C.; Lunguleasa, A.; Spirchez, C.; Ciobanu, V. Influence of INGER and TORDIS energetic willow clones planted on contaminated soil on the survival rates, yields and calorific value. Forests 2021, 12, 826. [Google Scholar] [CrossRef]
- Tőzsér, D.; Harangi, S.; Baranyai, E.; Lakatos, G.; Fülöp, Z.; Tóthmérész, B.; Simon, E. Phytoextraction with Salix viminalis in a moderately to strongly contaminated area. Environ. Sci. Pollut. Res. 2018, 25, 3275–3290. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Knee, M.; Quigley, M.F. Cadmium and Copper Uptake and Translocation in Five Willow (Salix L.) Species. Int. J. Phytoremediation 2004, 6, 269–287. [Google Scholar] [CrossRef]
- Drzewiecka, K.; Mleczek, M.; Gąsecka, M.; Magdziak, Z.; Budka, A.; Chadzinikolau, T.; Kaczmarek, Z.; Goliński, P. Copper and nickel co-treatment alters metal uptake and stress parameters of Salix purpurea × viminalis. J. Plant Physiol. 2017, 216, 125–134. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Verhagen, A. Climate change and its repercussions for the potato supply chain. Potato Res. 2008, 51, 223–237. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Naliwajski, M.; Skłodowska, M. The relationship between the antioxidant system and proline metabolism in the leaves of cucumber plants acclimated to salt stress. Cells 2021, 10, 609. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Ngole, V.M.; Ekosse, G.I.E. Copper, nickel and zinc contamination in soils within the precincts of mining and landfilling environments. Int. J. Environ. Sci. Technol. 2012, 9, 485–494. [Google Scholar] [CrossRef]
- Simončič, A.; Sušin, J.; Šinkovec, M.; Leskovšek, R.; Čuš, F.; Žnidaršič Pongrac, V.; Baša Česnik, H. Twelve-year investigation of copper soil concentrations shows that vineyards are at risk. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2017, 67, 381–394. [Google Scholar] [CrossRef]
- Tóth, G.; Hermann, T.; da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Helios, W. Effect of white clover (Trifolium repens L.) undersowing cultivation and nitrogen fertilization on weed infestation, biomass yield and its component, content and uptake of macroelements of willow (Salix viminalis L.). Agronomy 2021, 11, 786. [Google Scholar] [CrossRef]
- Kim, H.G.; Song, H.J.; Jeong, M.J.; Seo, Y.L.; Yang, J.K.; Yoo, S.B.; Choi, M.S. Bioethanol production by enzymatic saccharification of Salix viminalis var. gigantea biomass. Forest Sci. Technol. 2014, 10, 67–72. [Google Scholar] [CrossRef]
- Kuś, J.; Matyka, M. Productivity of selected crops planted for energy purposes depending on soil quality. Fragm. Agron. 2009, 26, 103–110. [Google Scholar]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef]
- Peršić, V.; Ament, A.; Antunović Dunić, J.; Drezner, G.; Cesar, V. PEG-induced physiological drought for screening winter wheat genotypes sensitivity–integrated biochemical and chlorophyll a fluorescence analysis. Front. Plant Sci. 2022, 13, 987702. [Google Scholar] [CrossRef]
- Baker, A.J.M.; McGrath, S.P.; Sidoli, C.M.D.; Reeves, R.D. The possibility of in situ heavy metal decontamination of polluted soils using crops of metal-accumulating plants. Resour. Conserv. Recycl. 1994, 11, 41–49. [Google Scholar] [CrossRef]
- Weatherley, P.E.; Slatyer, R.O. Relationship between relative turgidity and diffusion pressure deficit in leaves. Nature 1957, 179, 1085–1086. [Google Scholar] [CrossRef]
- Baker, A.J. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Ma, L.Q.; Komar, K.M.; Tu, C.; Zhang, W.; Cai, Y.; Kennelley, E.D. A fern that hyperaccumulates arsenic. Nature 2001, 409, 579. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Elegant graphics for data analysis (ggplot2). In Applied Spatial Data Analysis R; Springer: New York, NY, USA, 2009; pp. 784–785. [Google Scholar]
- Frejowski, A.; Bondaruk, J.; Duda, A. Challenges and opportunities for end-of-life coal mine sites: Black-to-Green energy approach. Energies 2021, 14, 1385. [Google Scholar] [CrossRef]
- Perea-Moreno, M.A.; Samerón-Manzano, E.; Perea-Moreno, A.J. Biomass as renewable energy: Worldwide research trends. Sustainability 2019, 11, 863. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Niksa, D.; Krzyżaniak, M.; Tworkowski, J.; Szczukowski, S. Willow productivity from small-and large-scale experimental plantations in Poland from 2000 to 2017. Renew. Sustain. Energy Rev. 2019, 101, 461–475. [Google Scholar] [CrossRef]
- Su, R.; Xie, T.; Yao, H.; Chen, Y.; Wang, H.; Dai, X.; Wang, Y.; Shi, L.; Luo, Y. Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil. Int. J. Environ. Res. Public Health 2022, 19, 14968. [Google Scholar] [CrossRef]
- Pouresmaieli, M.; Ataei, M.; Forouzandeh, P.; Azizollahi, P.; Mahmoudifard, M. Recent progress on sustainable phytoremediation of heavy metals from soil. J. Environ. Chem. Eng. 2022, 10, 108482. [Google Scholar] [CrossRef]
- Su, R.; Ou, Q.; Wang, H.; Dai, X.; Chen, Y.; Luo, Y.; Yao, H.; Ouyang, D.; Li, Z.; Wang, Z. Organic-inorganic composite modifiers enhance restoration potential of Nerium oleander L. to lead-zinc tailing: Application of phytoremediation. Environ. Sci. Pollut. Res. Int. 2023, 30, 56569–56579. [Google Scholar] [CrossRef] [PubMed]
- Fagnano, M.; Visconti, D.; Fiorentino, N. Agronomic approaches for characterization, remediation, and monitoring of contaminated sites. Agronomy 2020, 10, 1335. [Google Scholar] [CrossRef]
- Łapiński, D.; Wiater, J.; Szatyłowicz, E. The content of heavy metals in waste as an indicator determining the possibilities of their agricultural use. J. Ecol. Eng. 2019, 20, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Pandita, S.; Sidhu, G.P.S.; Sharma, A.; Khanna, K.; Kaur, P.; Bali, A.S.; Setia, R. Copper bioavailability, uptake, toxicity and tolerance in plants: A comprehensive review. Chemosphere 2021, 262, 127810. [Google Scholar] [CrossRef]
- Rai, S.; Singh, P.K.; Mankotia, S.; Swain, J.; Satbhai, S.B. Iron homeostasis in plants and its crosstalk with copper, zinc, and manganese. Plant Stress 2021, 1, 100008. [Google Scholar] [CrossRef]
- Martins, L.L.; Mourato, M.P. Effect of Excess Copper on Tomato Plants: Growth Parameters, Enzyme Activities, Chlorophyll, and Mineral Content. J. Plant Nutr. 2006, 29, 2179–2198. [Google Scholar] [CrossRef]
- Rehman, M.; Liu, L.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Pollut. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef]
- Krohling, C.A.; Eutrópio, F.J.; Bertolazi, A.A.; Dobbss, L.B.; Campostrini, E.; Dias, T.; Ramos, A.C. Ecophysiology of iron homeostasis in plants. Soil Sci. Plant Nutr. 2016, 62, 39–47. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef]
- Acosta-Gamboa, L.M.; Liu, S.; Langley, E.; Campbell, Z.; Castro-Guerrero, N.; Mendoza-Cozatl, D.; Lorence, A. Moderate to severe water limitation differentially affects the phenome and ionome of Arabidopsis. Funct. Plant Biol. 2017, 44, 94–106. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; del Mar Rubio-Wilhelmi, M.; Cervilla, L.M.; Blasco, B.; Rios, J.J.; Leyva, R.; Romero, L.; Ruiz, J.M. Study of the ionome and uptake fluxes in cherry tomato plants under moderate water stress conditions. Plant Soil 2010, 335, 339–347. [Google Scholar] [CrossRef]
- D’Oria, A.; Courbet, G.; Billiot, B.; Jing, L.; Pluchon, S.; Arkoun, M.; Maillard, A.; Roux, C.P.; Trouverie, J.; Etienne, P.; et al. Drought specifically downregulates mineral nutrition: Plant ionomic content and associated gene expression. Plant Direct 2022, 6, e402. [Google Scholar] [CrossRef] [PubMed]
- Schat, H. The role of phytochelatins in constitutive and adaptive heavy metal tolerances in hyperaccumulator and non-hyperaccumulator metallophytes. J. Exp. Bot. 2002, 53, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Chandregowda, M.H.; Tjoelker, M.G.; Pendall, E.; Zhang, H.; Churchill, A.C.; Power, S.A. Belowground carbon allocation, root trait plasticity, and productivity during drought and warming in a pasture grass. J. Exp. Bot. 2023, 74, 2127–2145. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Bündig, C.; Vu, T.H.; Meise, P.; Seddig, S.; Schum, A.; Winkelmann, T. Variability in osmotic stress tolerance of starch potato genotypes (Solanum tuberosum L.) as revealed by an in vitro screening: Role of proline, osmotic adjustment and drought response in pot trials. J. Agron. Crop Sci. 2017, 203, 206–218. [Google Scholar] [CrossRef]
- Lazcano-Ferrat, I.; Lovatt, C.J. Relationship between relative water content, nitrogen pools, and growth of Phaseolus vulgaris L. and P. acutifolius A. Gray during water deficit. Crop Sci. 1999, 39, 467–475. [Google Scholar] [CrossRef]
- Zouari, M.; Ahmed, C.B.; Elloumi, N.; Bellassoued, K.; Delmail, D.; Labrousse, P.; Ben Abdallah, F.; Rouina, B.B. Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv. Chemlali exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2016, 128, 195–205. [Google Scholar] [CrossRef]
- Drzewiecka, K.; Mleczek, M.; Gąsecka, M.; Magdziak, Z.; Goliński, P.; Chadzinikolau, T. Copper phytoextraction with Salix purpurea × viminalis under various Ca/Mg ratios. Part 2. Effect on organic acid, phenolics and salicylic acid contents. Acta Physiol. Plant. 2014, 36, 903–913. [Google Scholar] [CrossRef]
- Mleczek, M.; Budka, A.; Gąsecka, M.; Budzyńska, S.; Drzewiecka, K.; Magdziak, Z.; Rutkowski, P.; Goliński, P.; Niedzielski, P. Copper, lead and zinc interactions during phytoextraction using Acer platanoides L.—A pot trial. Environ. Sci. Pollut. Res. 2023, 30, 27191–27207. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).