
Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change

 , ,
, ,  ,
,  , ,
, , 

Abstract
:1. Introduction
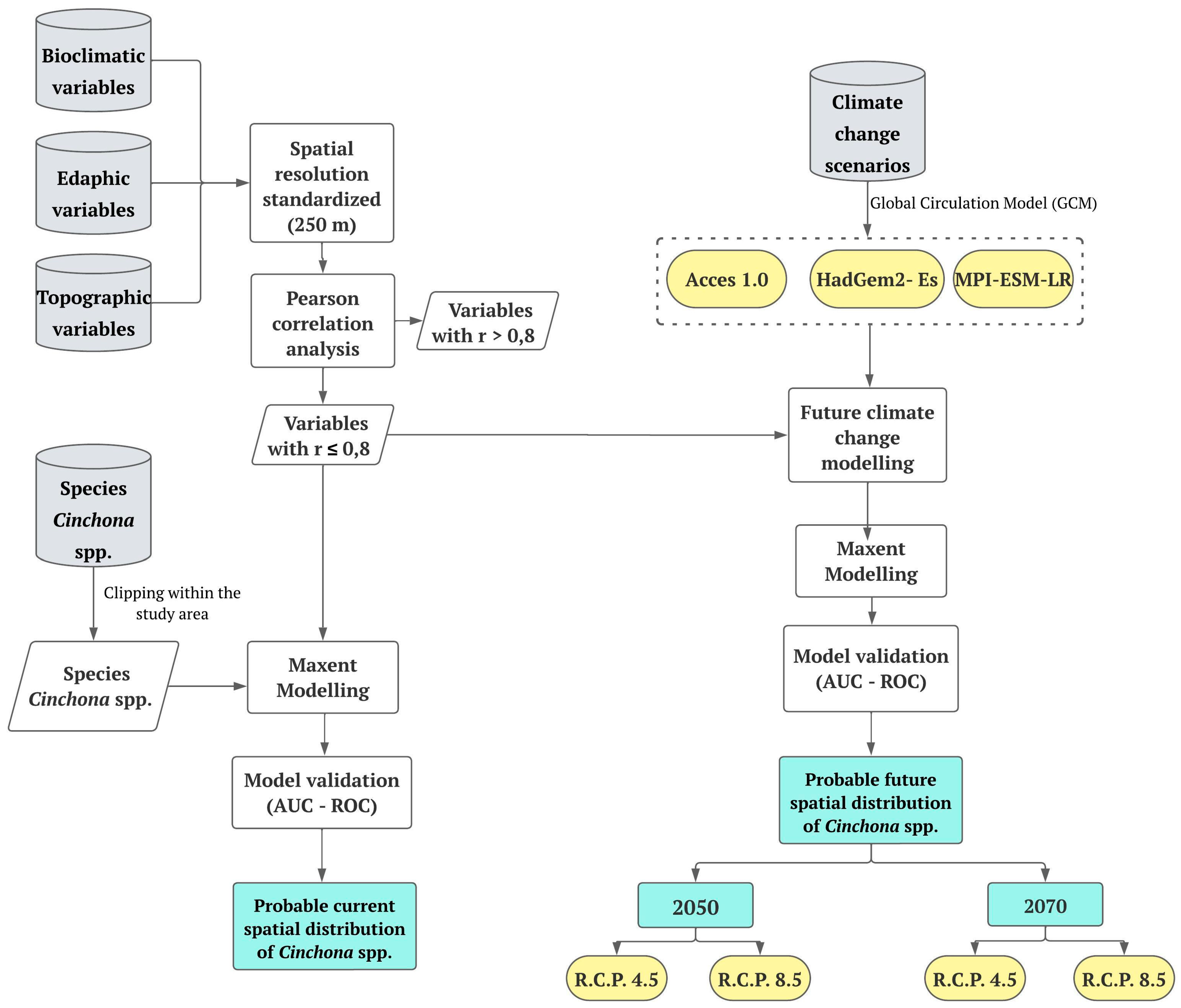
2. Materials and Methods
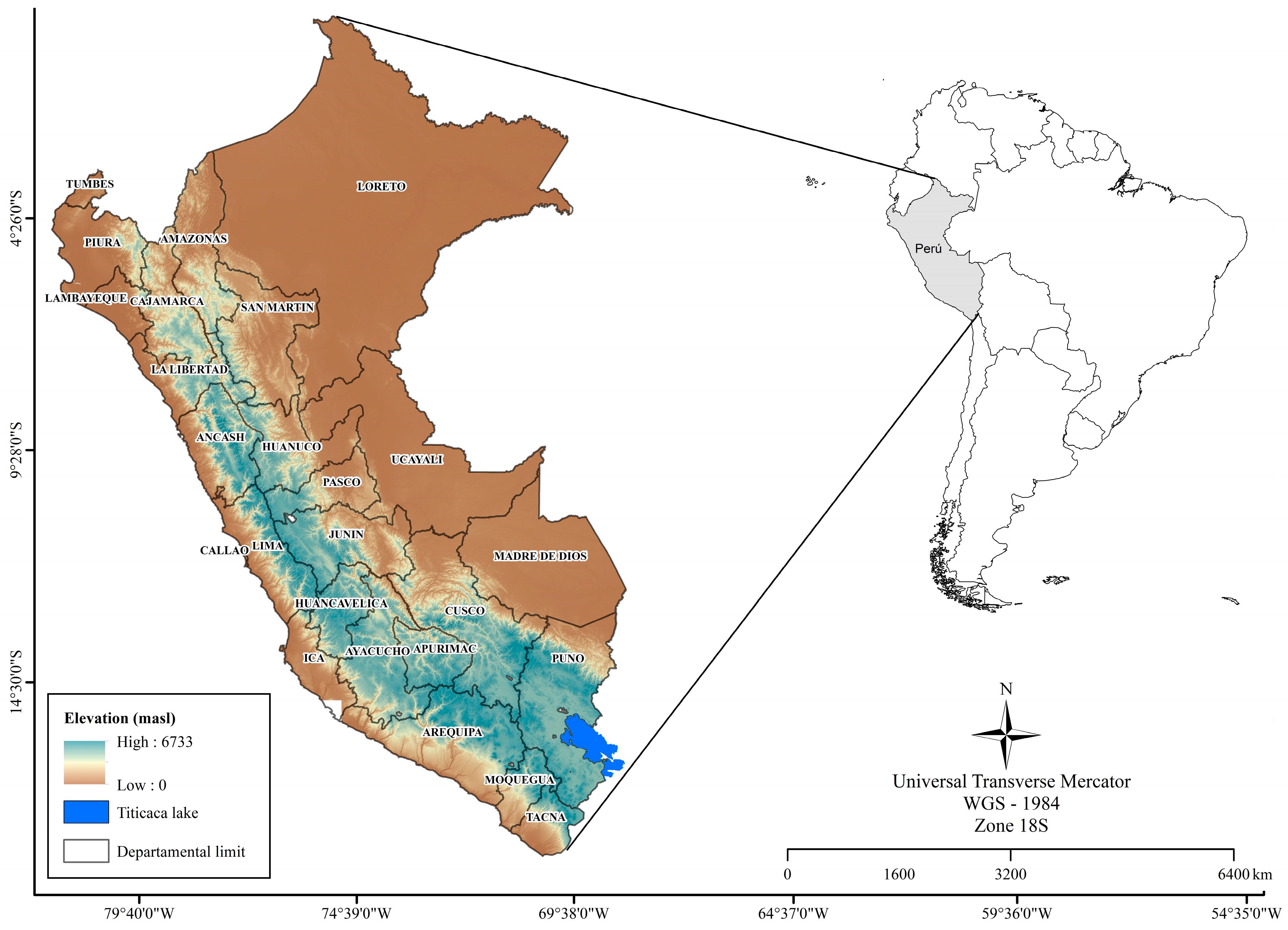
2.1. Study Area
2.2. Database and Processing of the Occurrence of the Genus Cinchona
2.3. Variables and Processing
2.4. Selection of Climate Models for Future Distribution
2.5. Modeling
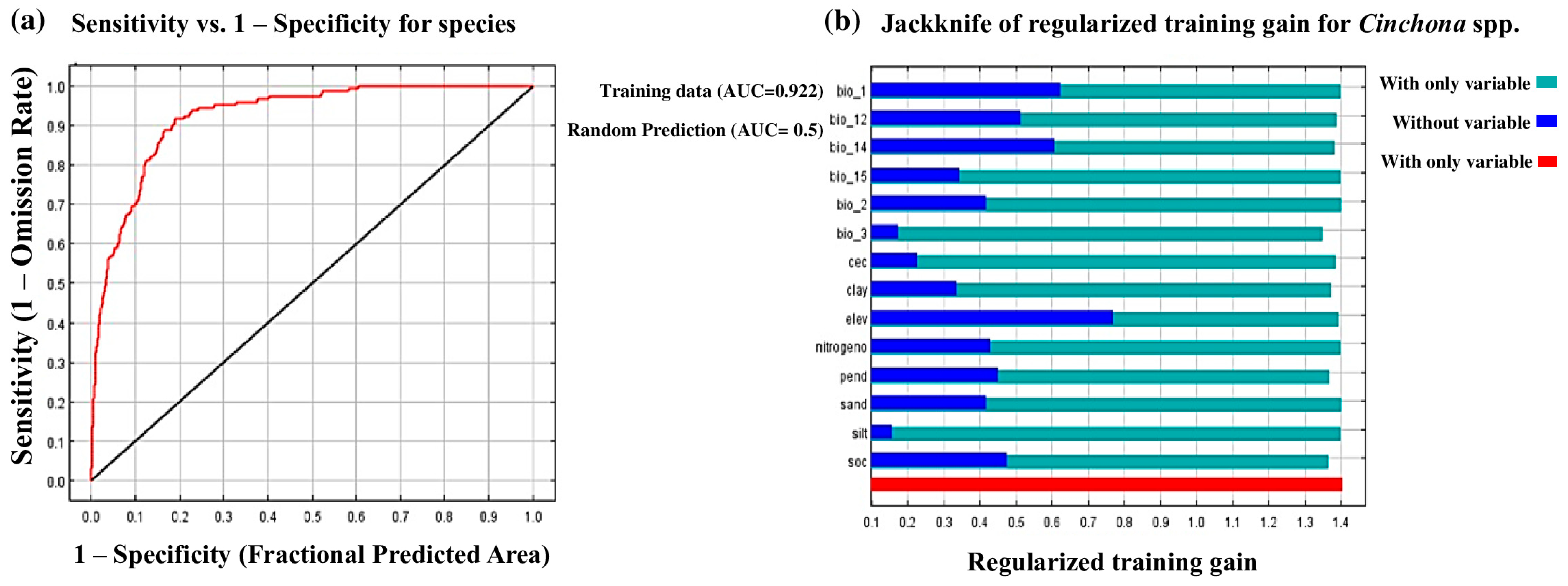
2.6. Mode Evaluation
3. Results
3.1. Contribution of the Variables Selected for the Study and the Probabilistic Model
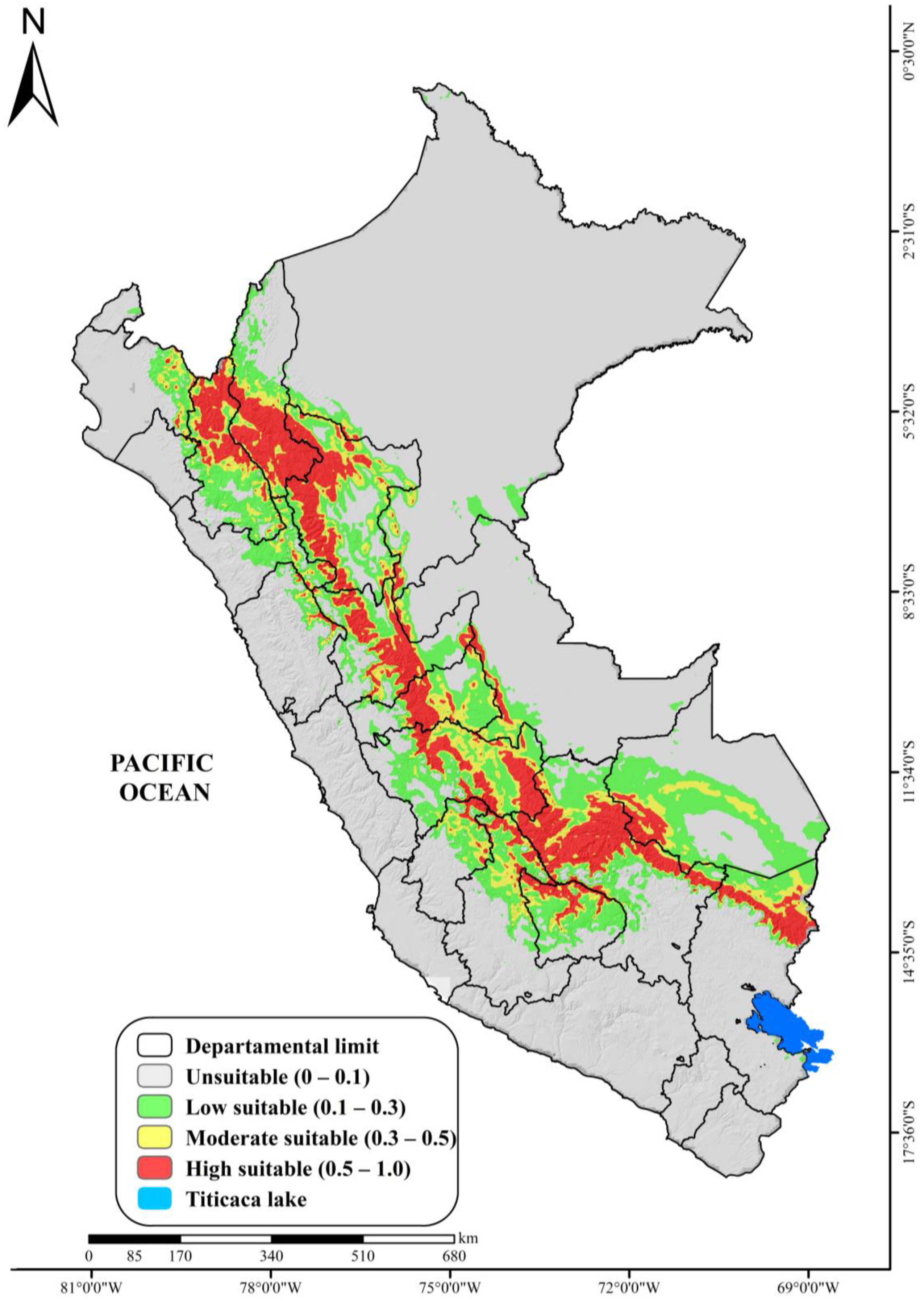
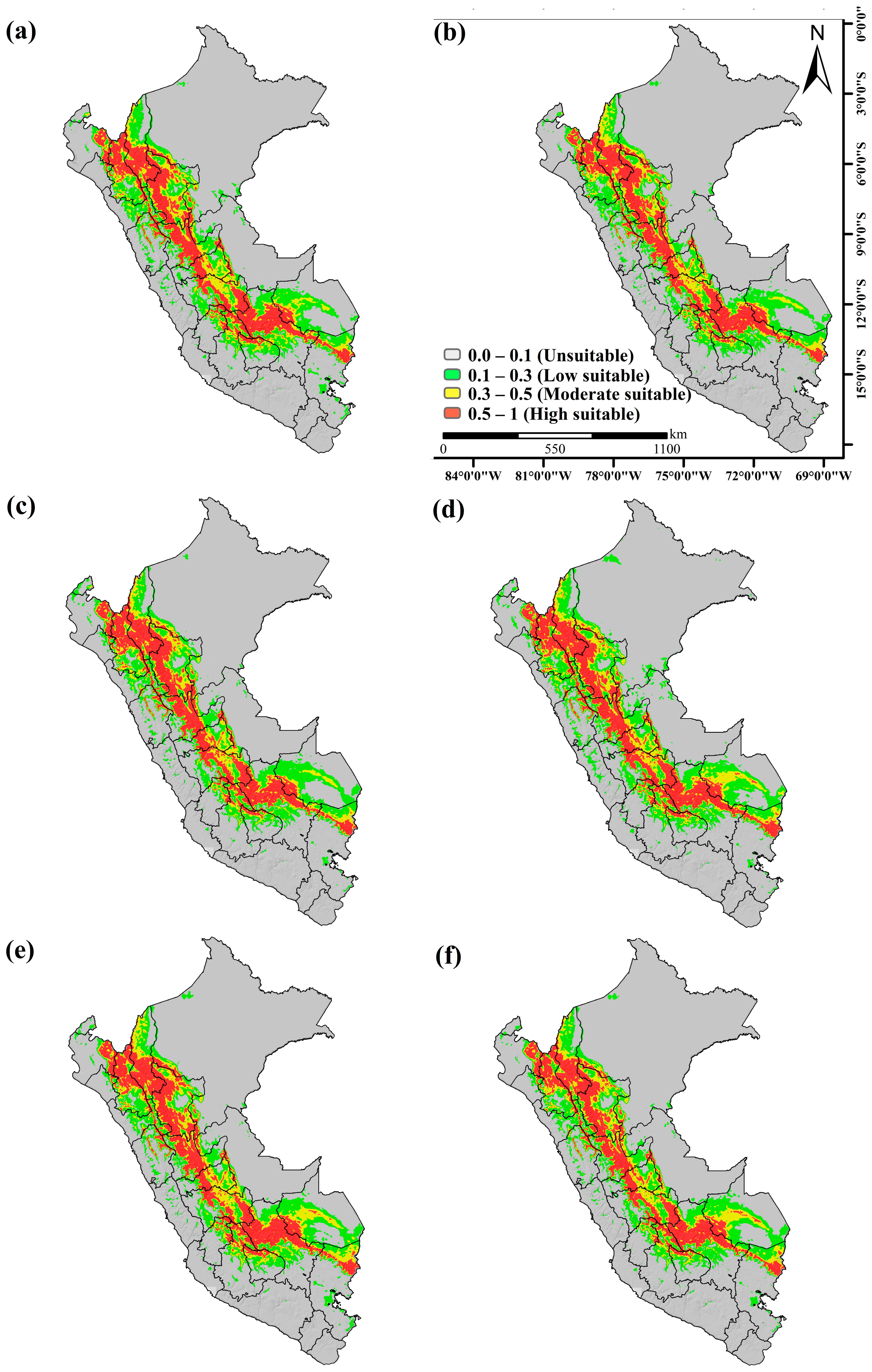
3.2. Probability of Areas for the Current Distribution of the Genus cinchona spp.
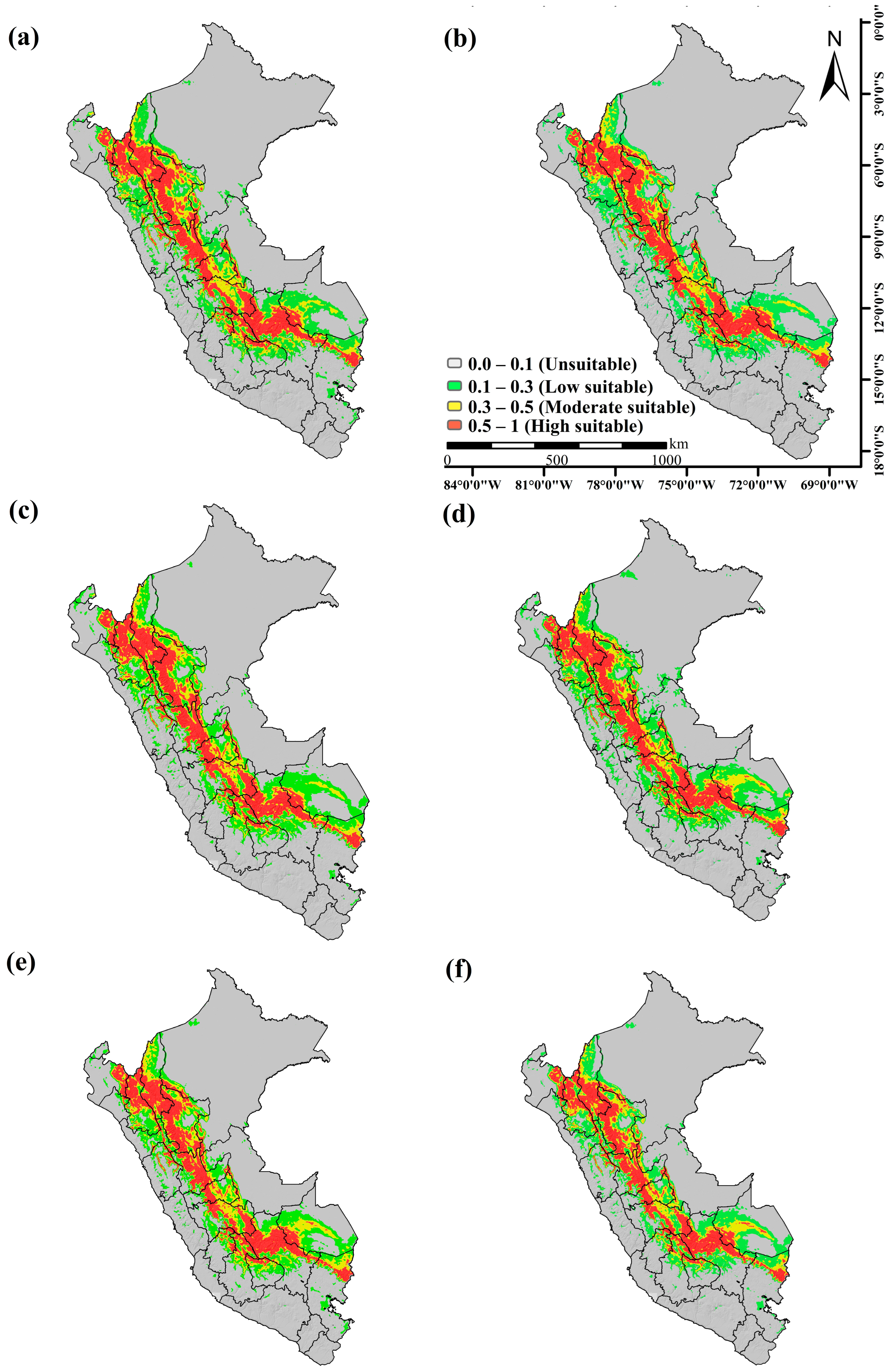
3.3. Potential Distribution of Cinchona spp. under Climate Change Scenario
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González-Orozco, C.E.; Guillén, E.G.; Cuvi, N. Changes of Cinchona Distribution over the Past Two Centuries in the Northern Andes. R. Soc. Open Sci. 2023, 10, 230229. [Google Scholar] [CrossRef]
- Cárdenas, G.P.; Bravo, N.; Barboza, E.; Salazar, W.; Ocaña, J.; Vázquez, M.; Lobato, R.; Injante, P.; Arbizu, C.I. Current and Future Distribution of Shihuahuaco (Dipteryx Spp.) under Climate Change Scenarios in the Central-Eastern Amazon of Peru. Sustainability 2023, 15, 7789. [Google Scholar] [CrossRef]
- Aymard, C.G.A.; Aymard, C.G.A. Breve Reseña de Los Aspectos Taxonómicos y Nomenclaturales Actuales Del Género Cinchona (Rubiaceae-Cinchoneae). Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 2019, 43, 234–241. [Google Scholar] [CrossRef]
- Vivanco-Galván, O.; Jiménez, Y.; Castillo Malla, D.P.; Lucero, H. Blue LED Light Enhances the Growth of Cinchona Officinalis 7L. Cultured in Vitro. SPIE 2022, 12210, 12210067. [Google Scholar] [CrossRef]
- Cuvi, N. Legacies of Boldness: Caldas, Humboldt and Knowledge about Cinchonas, 1801–1821. Hist. Cienc. Saude Manguinhos 2022, 29, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Canales, N.A.; Gress Hansen, T.N.; Cornett, C.; Walker, K.; Driver, F.; Antonelli, A.; Maldonado, C.; Nesbitt, M.; Barnes, C.J.; Rønsted, N. Historical Chemical Annotations of Cinchona Bark Collections Are Comparable to Results from Current Day High-Pressure Liquid Chromatography Technologies. J. Ethnopharmacol. 2020, 249, 112375. [Google Scholar] [CrossRef] [PubMed]
- Yacila, K.A.P.; Suarez-Peña, E.A.; Pera, C.A.T.; Requena, L.A.B.; Sánchez, L.X.L.; Rodríguez, I.Z.; Paredes-Vilca, O.J.; García, J.Y.D. Genetic Diversity and Phylogeny of the Genus Cinchona in Cutervo National Park, Peru. Sci. Agropecu. 2021, 12, 517–524. [Google Scholar] [CrossRef]
- García, L.; Veneros, J.; Chavez, S.G.; Oliva, M.; Rojas-Briceño, N.B. World Historical Mapping and Potential Distribution of Cinchona Spp. in Peru as a Contribution for Its Restoration and Conservation. J. Nat. Conserv. 2022, 70, 126290. [Google Scholar] [CrossRef]
- Sasongko, N.D.; Yuniati, A.; Billmora, L.A.G. Genetic Profiles of Three Cinchona Species in Junghuhn Natural Reserve, Indonesia. SABRAO J. Breed. Genet. 2021, 53, 697–709. [Google Scholar] [CrossRef]
- Aarts, G.; Fieberg, J.; Matthiopoulos, J. Comparative Interpretation of Count, Presence–Absence and Point Methods for Species Distribution Models. Methods Ecol. Evol. 2012, 3, 177–187. [Google Scholar] [CrossRef]
- Báez, J.C.; de Stephanis, R.; García-Ramos, M.C.; Ramos, M.L.; Pascual, P. Differential Space Distribution of the Genus Balaenoptera in the Eastern Tropical Atlantic Ocean. Reg. Stud. Mar. Sci. 2020, 37, 101346. [Google Scholar] [CrossRef]
- Pathak, A.K.; Verma, P.; Dayal, R.; Sarkar, U.K. Species Distribution Modeling and Assessment of Environmental Drivers Responsible for Distribution and Preferred Niche of Critically Endangered and Endemic Ornamental Freshwater Fish Species of the Genus Sahyadria. Environ. Sci. Pollut. Res. 2022, 29, 50584–50597. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhou, L.; Qi, S.; Jin, M.; Hu, J.; Lu, J. Effect of Habitat Factors on the Understory Plant Diversity of Platycladus Orientalis Plantations in Beijing Mountainous Areas Based on MaxEnt Model. Ecol. Indic. 2021, 129, 107917. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Stickley, S.F.; Fraterrigo, J.M. Microclimate Species Distribution Models Estimate Lower Levels of Climate-Related Habitat Loss for Salamanders. J. Nat. Conserv. 2023, 72, 126333. [Google Scholar] [CrossRef]
- Daniel, J.; Horrocks, J.; Umphrey, G.J. Efficient Modelling of Presence-Only Species Data via Local Background Sampling. J. Agric. Biol. Environ. Stat. 2020, 25, 90–111. [Google Scholar] [CrossRef]
- Fernandes, R.F.; Honrado, J.P.; Guisan, A.; Roxo, A.; Alves, P.; Martins, J.; Vicente, J.R. Species Distribution Models Support the Need of International Cooperation towards Successful Management of Plant Invasions. J. Nat. Conserv. 2019, 49, 85–94. [Google Scholar] [CrossRef]
- Torreblanca, E.; Caminãs, J.A.; MacIás, D.; Garciá-Barcelona, S.; Real, R.; Baéz, J.C. Using Opportunistic Sightings to Infer Differential Spatio-Temporal Use of Western Mediterranean Waters by the Fin Whale. PeerJ 2019, 2019, e6673. [Google Scholar] [CrossRef]
- Tang, Y.; Winkler, J.A.; Viña, A.; Wang, F.; Zhang, J.; Zhao, Z.; Connor, T.; Yang, H.; Zhang, Y.; Zhang, X.; et al. Expanding Ensembles of Species Present-Day and Future Climatic Suitability to Consider the Limitations of Species Occurrence Data. Ecol. Indic. 2020, 110, 105891. [Google Scholar] [CrossRef]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected Shifts in Coffea Arabica Suitability among Major Global Producing Regions Due to Climate Change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef]
- Schnase, J.L.; Carroll, M.L.; Gill, R.L.; Tamkin, G.S.; Li, J.; Strong, S.L.; Maxwell, T.P.; Aronne, M.E.; Spradlin, C.S. Toward a Monte Carlo Approach to Selecting Climate Variables in MaxEnt. PLoS ONE 2021, 16, e0237208. [Google Scholar] [CrossRef] [PubMed]
- Thomas Ledig, F.; Rehfeldt, G.E.; Sáenz-Romero, C.; Flores-López, C. Projections of Suitable Habitat for Rare Species under Global Warming Scenarios. Am. J. Bot. 2010, 97, 970–987. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, G.; Crouzet, P.; Gessner, J.; Rochard, E. Global Warming Impacts and Conservation Responses for the Critically Endangered European Atlantic Sturgeon. Biol. Conserv. 2010, 143, 2441–2452. [Google Scholar] [CrossRef]
- Guan, Y.; Hohn, S.; Wild, C.; Merico, A. Vulnerability of Global Coral Reef Habitat Suitability to Ocean Warming, Acidification and Eutrophication. Glob. Chang. Biol. 2020, 26, 5646–5660. [Google Scholar] [CrossRef]
- Mao, M.; Chen, S.; Ke, Z.; Qian, Z.; Xu, Y. Using MaxEnt to Predict the Potential Distribution of the Little Fire Ant (Wasmannia Auropunctata) in China. Insects 2022, 13, 1008. [Google Scholar] [CrossRef]
- Mahmoodi, S.; Heydari, M.; Ahmadi, K.; Khwarahm, N.R.; Karami, O.; Almasieh, K.; Naderi, B.; Bernard, P.; Mosavi, A. The Current and Future Potential Geographical Distribution of Nepeta Crispa Willd., an Endemic, Rare and Threatened Aromatic Plant of Iran: Implications for Ecological Conservation and Restoration. Ecol. Indic. 2022, 137, 108752. [Google Scholar] [CrossRef]
- Vásárhelyi, Z.; Scheuring, I.; Avilés, L. The Ecology of Spider Sociality: A Spatial Model. Am. Nat. 2022, 199, 776–788. [Google Scholar] [CrossRef]
- Hwang, J.H.; Kim, S.H.; Yoon, S.; Jung, S.; Kim, D.H.; Lee, W.H. Evaluation of Spatial Distribution of Three Major Leptocorisa (Hemiptera: Alydidae) Pests Using MaxEnt Model. Insects 2022, 13, 750. [Google Scholar] [CrossRef]
- Mugiyo, H.; Chimonyo, V.G.P.; Kunz, R.; Sibanda, M.; Nhamo, L.; Ramakgahlele Masemola, C.; Modi, A.T.; Mabhaudhi, T. Mapping the Spatial Distribution of Underutilised Crop Species under Climate Change Using the MaxEnt Model: A Case of KwaZulu-Natal, South Africa. Clim. Serv. 2022, 28, 100330. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Li, R.; Wang, X.; Cheng, J.; Yang, Q.; Kong, H. AHP-GIS and MaxEnt for Delineation of Potential Distribution of Arabica Coffee Plantation under Future Climate in Yunnan, China. Ecol. Indic. 2021, 132, 108339. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.; Xu, C. Study on Taiwania Cryptomerioides under Climate Change: MaxEnt Modeling for Predicting the Potential Geographical Distribution. Glob. Ecol. Conserv. 2020, 24, e01313. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, P.; Hu, W.; Chen, G.; Zhang, D.; Chen, B.; Lei, G. Modeling the Climate Suitability of Northernmost Mangroves in China under Climate Change Scenarios. Forests 2022, 13, 64. [Google Scholar] [CrossRef]
- Cotrina Sánchez, A.; Rojas Briceño, N.B.; Bandopadhyay, S.; Ghosh, S.; Torres Guzmán, C.; Oliva, M.; Guzman, B.K.; Salas López, R. Biogeographic Distribution of Cedrela Spp. Genus in Peru Using Maxent Modeling: A Conservation and Restoration Approach. Diversity 2021, 13, 261. [Google Scholar] [CrossRef]
- Blazquez-Soriano, A.; Ramos-Sandoval, R. Information Transfer as a Tool to Improve the Resilience of Farmers against the Effects of Climate Change: The Case of the Peruvian National Agrarian Innovation System. Agric. Syst. 2022, 200, 103431. [Google Scholar] [CrossRef]
- Ramírez, I.J.; Lee, J. Deconstructing the Spatial Effects of El Niño and Vulnerability on Cholera Rates in Peru: Wavelet and GIS Analyses. Spat. Spatiotemporal Epidemiol. 2022, 40, 100474. [Google Scholar] [CrossRef]
- Zhang, H.T.; Wang, W.T. Prediction of the Potential Distribution of the Endangered Species Meconopsis Punicea Maxim under Future Climate Change Based on Four Species Distribution Models. Plants 2023, 12, 1376. [Google Scholar] [CrossRef]
- Kayet, N.; Chakrabarty, A.; Pathak, K.; Sahoo, S.; Dutta, T.; Hatai, B.K. Comparative Analysis of Multi-Criteria Probabilistic FR and AHP Models for Forest Fire Risk (FFR) Mapping in Melghat Tiger Reserve (MTR) Forest. J. For. Res. 2020, 31, 565–579. [Google Scholar] [CrossRef]
- Xu, D.; Zhuo, Z.; Wang, R.; Ye, M.; Pu, B. Modeling the Distribution of Zanthoxylum Armatum in China with MaxEnt Modeling. Glob. Ecol. Conserv. 2019, 19, e00691. [Google Scholar] [CrossRef]
- Kufa, C.A.; Bekele, A.; Atickem, A. Impacts of Climate Change on Predicted Habitat Suitability and Distribution of Djaffa Mountains Guereza (Colobus Guereza Gallarum, Neumann 1902) Using MaxEnt Algorithm in Eastern Ethiopian Highland. Glob. Ecol. Conserv 2022, 35, e02094. [Google Scholar] [CrossRef]
- Scrivanti, L.R.; Anton, A.M. Impact of Climate Change on the Andean Distribution of Poa Scaberula (Poaceae). Flora 2021, 278, 151805. [Google Scholar] [CrossRef]
- Berhanu, Y.; Tassie, N.; Sintayehu, D.W. Predicting the Current and Future Suitable Habitats for Endemic and Endangered Ethiopian Wolf Using MaxEnt Model. Heliyon 2022, 8, e10223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting Potential Suitable Habitats of Chinese Fir under Current and Future Climatic Scenarios Based on Maxent Model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, P.; Lin, F.; Yang, W.; Gaisberger, H.; Christopher, K.; Zheng, Y. MaxEnt Modelling for Predicting the Potential Distribution of a near Threatened Rosewood Species (Dalbergia Cultrata Graham Ex Benth). Ecol. Eng. 2019, 141, 105612. [Google Scholar] [CrossRef]
- Mehmud, S.; Kalita, N.; Roy, H.; Sahariah, D. Species Distribution Modelling of Calamus Floribundus Griff. (Arecaceae) Using Maxent in Assam. Acta Ecol. Sin. 2022, 42, 115–121. [Google Scholar] [CrossRef]
- Hand, D.J. Measuring Classifier Performance: A Coherent Alternative to the Area under the ROC Curve. Mach. Learn. 2009, 77, 103–123. [Google Scholar] [CrossRef]
- Wang, R.; Jiang, C.; Liu, L.; Shen, Z.; Yang, J.; Wang, Y.; Hu, J.; Wang, M.; Hu, J.; Lu, X.; et al. Prediction of the Potential Distribution of the Predatory Mite Neoseiulus Californicus McGregor in China Using MaxEnt. Glob. Ecol. Conserv. 2021, 29, e01733. [Google Scholar] [CrossRef]
- Zheng, T.; Sun, J.Q.; Shi, X.J.; Liu, D.L.; Sun, B.Y.; Deng, Y.; Zhang, D.L.; Liu, S.M. Evaluation of Climate Factors Affecting the Quality of Red Huajiao (Zanthoxylum Bungeanum Maxim.) Based on UPLC-MS/MS and MaxEnt Model. Food Chem. X 2022, 16, 100522. [Google Scholar] [CrossRef]
- Shen, Y.; Tu, Z.; Zhang, Y.; Zhong, W.; Xia, H.; Hao, Z.; Zhang, C.; Li, H. Predicting the Impact of Climate Change on the Distribution of Two Relict Liriodendron Species by Coupling the MaxEnt Model and Actual Physiological Indicators in Relation to Stress Tolerance. J. Environ. Manag. 2022, 322, 116024. [Google Scholar] [CrossRef]
- Arbizu, C.I.; Ferro-Mauricio, R.D.; Chávez-Galarza, J.C.; Guerrero-Abad, J.C.; Vásquez, H.V.; Maicelo, J.L. The Complete Chloroplast Genome of the National Tree of Peru, Quina (Cinchona Officinalis L., Rubiaceae). Mitochondrial DNA B Resour. 2021, 6, 2781–2783. [Google Scholar] [CrossRef]
- Ma, Q.; Li, X.; Wu, S.; Zeng, F. Potential Geographical Distribution of Stipa Purpurea across the Tibetan Plateau in China under Climate Change in the 21st Century. Glob. Ecol. Conserv. 2022, 35, e02064. [Google Scholar] [CrossRef]
- Villar, M.; Marcelo, F.; Baselly, J. Evaluación de la Regeneración Natural de la Cinchona Officinalis L. en la Zona de Amortiguamiento del Bosque de Protección de Pagaibamba—II Etapa; Instituto Nacional de Innovación Agraria—INIA: La Molina, Peru, 2018; pp. 1–9. [Google Scholar]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the Spatial Distribution of Alternanthera Philoxeroides in China Based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Sandoval-Martínez, J.; Badano, E.I.; Guerra-Coss, F.A.; Flores Cano, J.A.; Flores, J.; Gelviz-Gelvez, S.M.; Barragán-Torres, F. Selecting Tree Species to Restore Forest under Climate Change Conditions: Complementing Species Distribution Models with Field Experimentation. J. Environ. Manag. 2023, 329, 117038. [Google Scholar] [CrossRef] [PubMed]
- Määttänen, A.M.; Virkkala, R.; Leikola, N.; Aalto, J.; Heikkinen, R.K. Combined Threats of Climate Change and Land Use to Boreal Protected Areas with Red-Listed Forest Species in Finland. Glob. Ecol. Conserv. 2023, 41, 1–15. [Google Scholar] [CrossRef]
- Guitérrez, E.; Trejo, I. Efecto Del Cambio Climático En La Distribución Potencial de Cinco Especies Arbóreas de Bosque Templado En México. Rev. Mex. Biodivers. 2014, 85, 179–188. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field Validation of an Invasive Species Maxent Model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Variables | Description | Source |
|---|---|---|---|
| Bioclimatic variables | bio01 | Annual Mean Temperature | Worldclim |
| bio02 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) | ||
| bio03 | Isothermality (BIO2/BIO7) (×100) | ||
| bio04 | Temperature Seasonality (standard deviation ×100) | ||
| bio05 | Max Temperature of Warmest Month | ||
| bio06 | Min Temperature of Coldest Month | ||
| bio07 | Temperature Annual Range (BIO5-BIO6) | ||
| bio08 | Mean Temperature of Wettest Quarter | ||
| bio09 | Mean Temperature of Driest Quarter | ||
| bio10 | Mean Temperature of Warmest Quarter | ||
| bio11 | Mean Temperature of Coldest Quarter | ||
| bio12 | Annual Precipitation | ||
| bio13 | Precipitation of Wettest Month | ||
| bio14 | Precipitation of Driest Month | ||
| bio15 | Precipitation Seasonality (Coefficient of Variación) | ||
| bio16 | Precipitation of Wettest Quarter | ||
| bio17 | Precipitation of Driest Quarter | ||
| bio18 | Precipitation of Warmest Quarter | ||
| bio19 | Precipitation of Coldest Quarter | ||
| Topographic variables | Pend | Slope | SRTM |
| Elev | Elevation | ||
| Aspect | Land aspect | ||
| Edaphic variables | Sand | Sand | Soilgrid |
| Clay | Clay | ||
| Limo | Silt | ||
| Nitrogeno | Nitrogen in the soil | ||
| Cec | Cation exchange capacity | ||
| Soc | Soil organic carbon |
| Departament | Unsuitable | Low Suitability | Moderate Suitability | High Suitability | ||||
|---|---|---|---|---|---|---|---|---|
| Km | % | Km | % | Km | % | Km | % | |
| Amazonas | 13,861.60 | 35.62 | 6855.00 | 17.61 | 4032.51 | 10.36 | 14,168.54 | 36.41 |
| Ancash | 30,777.46 | 87.07 | 2760.61 | 7.81 | 1250.05 | 3.54 | 561.85 | 1.59 |
| Apurimac | 5607.00 | 26.56 | 8978.83 | 42.53 | 2940.60 | 13.93 | 3587.72 | 16.99 |
| Arequipa | 60,423.51 | 100.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ayacucho | 28,100.59 | 64.59 | 7719.90 | 17.75 | 4911.70 | 11.29 | 2771.63 | 6.37 |
| Cajamarca | 5328.86 | 16.22 | 11,804.17 | 35.94 | 6420.38 | 19.55 | 9290.56 | 28.29 |
| Callao | 19.58 | 100.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cusco | 29,287.93 | 40.63 | 14,873.30 | 20.64 | 6621.21 | 9.19 | 21,293.71 | 29.54 |
| Huancavelica | 13,454.78 | 60.98 | 4595.35 | 20.83 | 2590.31 | 11.74 | 1424.60 | 6.46 |
| Huanuco | 12,369.96 | 33.25 | 9379.75 | 25.21 | 5104.99 | 13.72 | 10,345.83 | 27.81 |
| Ica | 19,825.18 | 100.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Junin | 8835.49 | 20.08 | 15,590.70 | 35.44 | 8779.67 | 19.96 | 10,791.43 | 24.53 |
| La Libertad | 15,040.74 | 61.83 | 6452.76 | 26.52 | 2052.49 | 8.44 | 781.65 | 3.21 |
| Lambayeque | 12,532.28 | 92.87 | 673.65 | 4.99 | 191.11 | 1.42 | 96.71 | 0.72 |
| Lima | 33,599.47 | 99.75 | 84.45 | 0.25 | 0.00 | 0.00 | 0.00 | 0.00 |
| Loreto | 355,021.50 | 96.80 | 7758.30 | 2.12 | 2970.35 | 0.81 | 1016.61 | 0.28 |
| Madre De Dios | 72,298.31 | 66.37 | 26,544.90 | 24.37 | 5914.25 | 5.43 | 4174.47 | 3.83 |
| Moquegua | 15,510.31 | 100.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Pasco | 6551.31 | 33.91 | 8013.65 | 41.48 | 4752.92 | 24.60 | 0.00 | 0.00 |
| Piura | 26,521.07 | 78.27 | 4512.74 | 13.32 | 2421.30 | 7.15 | 430.67 | 1.27 |
| Puno | 47,434.04 | 73.74 | 6327.83 | 9.84 | 4392.37 | 6.83 | 6167.76 | 9.59 |
| San Martin | 12,689.62 | 24.90 | 14,788.34 | 29.02 | 8879.58 | 17.42 | 14,603.72 | 28.66 |
| Tacna | 14,609.62 | 100.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tumbes | 3923.66 | 94.85 | 212.97 | 5.15 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ucayali | 92,285.07 | 89.06 | 6353.10 | 6.13 | 2938.02 | 2.84 | 2040.45 | 1.97 |
| Total | 935,908.94 | 73.07 | 164,280.30 | 12.83 | 77,163.81 | 6.02 | 103,547.89 | 8.08 |
| Climatic Model | RCP | Year | Unsuitable | Low Suitability | Moderate Suitability | High Suitability | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Km2 | % | Km2 | % | Km2 | % | Km | % | |||
| MPI-ESM-LR | 8.5 | 2050 | 887,610.74 | 70.31 | 154,305.43 | 12.22 | 75,749.80 | 6.00 | 144,812.33 | 11.47 |
| 2070 | 915,828.99 | 72.54 | 136,222.13 | 10.79 | 76,159.43 | 6.03 | 134,281.40 | 10.64 | ||
| HadGem2-Es | 2050 | 856,788.40 | 67.86 | 166,305.37 | 13.17 | 88,489.02 | 7.01 | 150,925.43 | 11.95 | |
| 2070 | 854,821.24 | 67.71 | 164,256.67 | 13.01 | 80,335.06 | 6.36 | 163,112.96 | 12.92 | ||
| Access 1.0 | 2050 | 881,473.61 | 69.71 | 142,311.06 | 11.25 | 82,134.19 | 6.50 | 158,625.01 | 12.54 | |
| 2070 | 875,345.81 | 69.34 | 155,740.81 | 12.34 | 84,643.80 | 6.70 | 146,691.04 | 11.62 | ||
| MPI-ESM-LR | 4.5 | 2050 | 861,742.47 | 68.26 | 163,697.77 | 12.97 | 82,076.25 | 6.50 | 154,988.40 | 12.28 |
| 2070 | 885,413.16 | 70.12 | 154,516.72 | 12.24 | 80,853.39 | 6.40 | 141,856.86 | 11.23 | ||
| HadGem2-Es | 2050 | 860,090.69 | 68.12 | 167,517.53 | 13.27 | 83,209.73 | 6.59 | 151,781.29 | 12.02 | |
| 2070 | 857,099.68 | 67.89 | 163,598.67 | 12.96 | 88,834.58 | 7.04 | 153,002.10 | 12.12 | ||
| Access 1.0 | 2050 | 882,769.28 | 69.92 | 152,931.78 | 12.11 | 84,630.40 | 6.70 | 142,185.15 | 11.26 | |
| 2070 | 861,766.00 | 68.26 | 164,886.54 | 13.06 | 87,146.92 | 6.90 | 148,705.64 | 11.78 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergara, A.J.; Cieza-Tarrillo, D.; Ocaña, C.; Quiñonez, L.; Idrogo-Vasquez, G.; Muñoz-Astecker, L.D.; Auquiñivin-Silva, E.A.; Cruzalegui, R.J.; Arbizu, C.I. Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change. Sustainability 2023, 15, 14109. https://doi.org/10.3390/su151914109
Vergara AJ, Cieza-Tarrillo D, Ocaña C, Quiñonez L, Idrogo-Vasquez G, Muñoz-Astecker LD, Auquiñivin-Silva EA, Cruzalegui RJ, Arbizu CI. Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change. Sustainability. 2023; 15(19):14109. https://doi.org/10.3390/su151914109
Chicago/Turabian StyleVergara, Alex J., Dennis Cieza-Tarrillo, Candy Ocaña, Lenin Quiñonez, Guillermo Idrogo-Vasquez, Lucas D. Muñoz-Astecker, Erick A. Auquiñivin-Silva, Robert J. Cruzalegui, and Carlos I. Arbizu. 2023. "Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change" Sustainability 15, no. 19: 14109. https://doi.org/10.3390/su151914109
APA StyleVergara, A. J., Cieza-Tarrillo, D., Ocaña, C., Quiñonez, L., Idrogo-Vasquez, G., Muñoz-Astecker, L. D., Auquiñivin-Silva, E. A., Cruzalegui, R. J., & Arbizu, C. I. (2023). Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change. Sustainability, 15(19), 14109. https://doi.org/10.3390/su151914109